
Genetic Variations of the DPYD Gene and Its Relationship with Ancestry Proportions in Different Ecuadorian Trihybrid Populations

 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Genotyping of DPYD SNVs
2.3. Genetic Ancestry Analysis
2.4. Statistical Analysis
3. Results
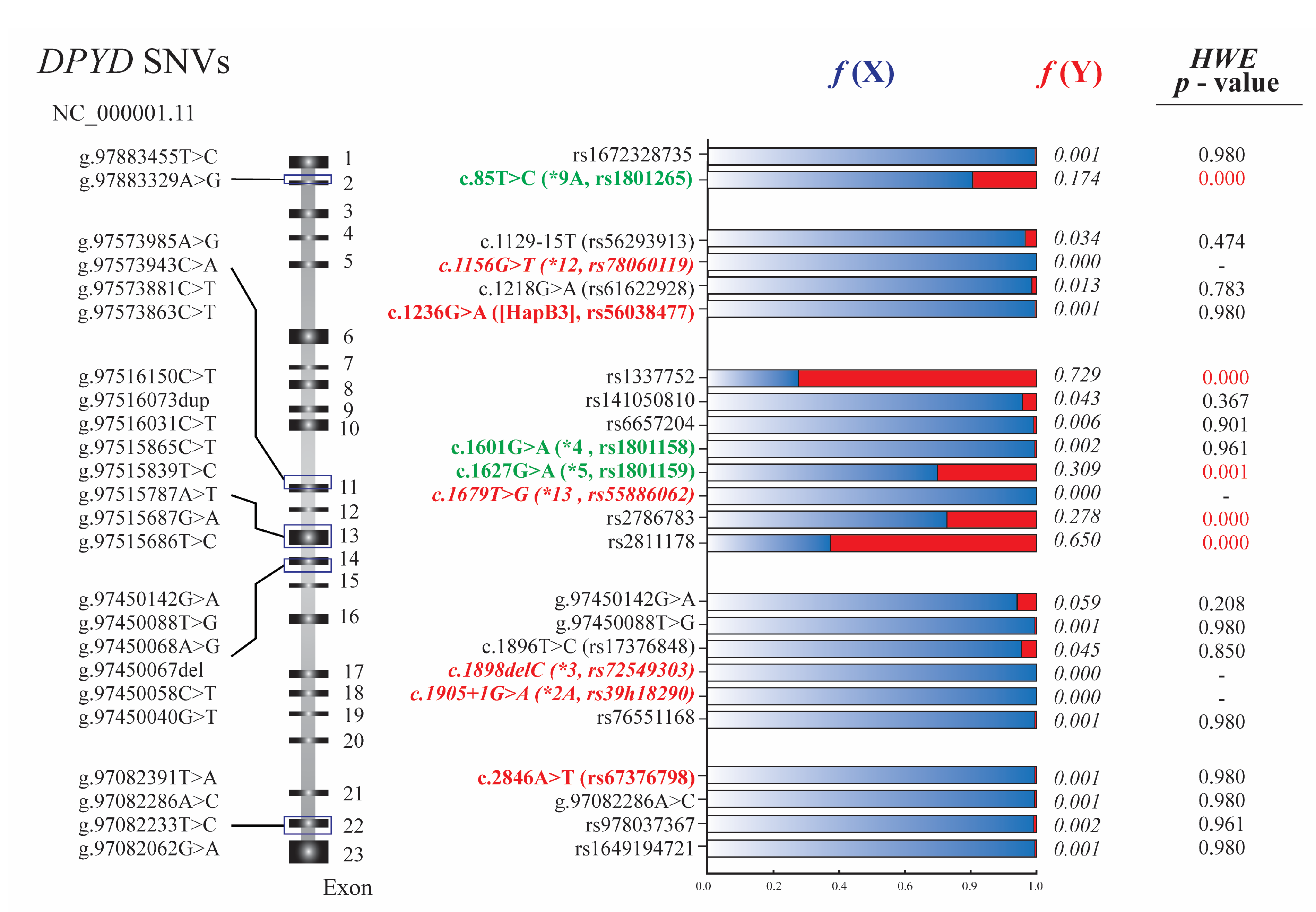
3.1. DPYD Genotyping and Hardy-Weinberg Equilibrium
3.2. Ancestry Proportions in Ecuadorian Populations
3.3. Relationship between DPYD Single Nucleotide Variants and Admixture Proportions in Ecuadorian Ethnic Groups
3.4. DPYD Variants and Genetic Distances in the Ecuadorian Population
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diasio, R.B. The role of dihydropyrimidine dehydrogenase (DPD) modulation in 5-FU pharmacology. Oncology 1998, 12, 23–27. [Google Scholar]
- Lamont, E.B.; Schilsky, R.L. The oral fluoropyrimidines in cancer chemotherapy. Clin. Cancer Res. 1999, 5, 2289–2296. [Google Scholar]
- Ichikawa, W. Prediction of clinical outcome of fluoropyrimidine-based chemotherapy for gastric cancer patients, in terms of the 5-fluorouracil metabolic pathway. Gastric Cancer 2006, 9, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Vodenkova, S.; Buchler, T.; Cervena, K.; Veskrnova, V.; Vodicka, P.; Vymetalkova, V. 5-fluorouracil and other fluoropyrimidines in colorectal cancer: Past, present and future. Pharmacol. Ther. 2020, 206, 107447. [Google Scholar] [CrossRef]
- Blum, J.L. The role of capecitabine, an oral, enzymatically activated fluoropyrimidine, in the treatment of metastatic breast cancer. Oncologist 2001, 6, 56–64. [Google Scholar] [CrossRef]
- Pointet, A.L.; Tougeron, D.; Pernot, S.; Pozet, A.; Bechade, D.; Trouilloud, I.; Lourenco, N.; Hautefeuille, V.; Locher, C.; Williet, N.; et al. Three fluoropyrimidine-based regimens in routine clinical practice after nab-paclitaxel plus gemcitabine for metastatic pancreatic cancer: An AGEO multicenter study. Clin. Res. Hepatol. Gastroenterol. 2020, 44, 295–301. [Google Scholar] [CrossRef]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Gmeiner, W.H. A narrative review of genetic factors affecting fluoropyrimidine toxicity. Precis. Cancer Med. 2021, 4, 38. [Google Scholar] [CrossRef]
- Koopman, M.; Antonini, N.F.; Douma, J.; Wals, J.; Honkoop, A.H.; Erdkamp, F.L.; de Jong, R.S.; Rodenburg, C.J.; Vreugdenhil, G.; Loosveld, O.J.; et al. Sequential versus combination chemotherapy with capecitabine, irinotecan, and oxaliplatin in advanced colorectal cancer (CAIRO): A phase III randomised controlled trial. Lancet 2007, 370, 135–142. [Google Scholar] [CrossRef]
- Twelves, C.; Wong, A.; Nowacki, M.P.; Abt, M.; Burris, H., 3rd; Carrato, A.; Cassidy, J.; Cervantes, A.; Fagerberg, J.; Georgoulias, V.; et al. Capecitabine as adjuvant treatment for stage III colon cancer. N. Engl. J. Med. 2005, 352, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Hou, J.; Deng, Q.; Zhong, Z. Predictive value of clinical toxicities of chemotherapy with fluoropyrimidines and oxaliplatin in colorectal cancer by DPYD and GSTP1 gene polymorphisms. World J. Surg. Oncol. 2020, 18, 321. [Google Scholar] [CrossRef]
- Smith, P.; Lavery, A.; Turkington, R.C. An overview of acute gastrointestinal side effects of systemic anti-cancer therapy and their management. Best Pract. Res. Clin. Gastroenterol. 2020, 48–49, 101691. [Google Scholar] [CrossRef]
- Kanduri, J.; More, L.A.; Godishala, A.; Asnani, A. Fluoropyrimidine-Associated Cardiotoxicity. Cardiol. Clin. 2019, 37, 399–405. [Google Scholar] [CrossRef]
- Omura, K. Clinical implications of dihydropyrimidine dehydrogenase (DPD) activity in 5-FU-based chemotherapy: Mutations in the DPD gene, and DPD inhibitory fluoropyrimidines. Int. J. Clin. Oncol. 2003, 8, 132–138. [Google Scholar] [CrossRef]
- Offer, S.M.; Fossum, C.C.; Wegner, N.J.; Stuflesser, A.J.; Butterfield, G.L.; Diasio, R.B. Comparative functional analysis of DPYD variants of potential clinical relevance to dihydropyrimidine dehydrogenase activity. Cancer Res. 2014, 74, 2545–2554. [Google Scholar] [CrossRef] [Green Version]
- Mattison, L.K.; Fourie, J.; Desmond, R.A.; Modak, A.; Saif, M.W.; Diasio, R.B. Increased prevalence of dihydropyrimidine dehydrogenase deficiency in African-Americans compared with Caucasians. Clin. Cancer Res. 2006, 12, 5491–5495. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Gupta, S.K.; Verma, M. Dihydropyrimidine dehydrogenase in the metabolism of the anticancer drugs. Cancer Chemother. Pharmacol. 2019, 84, 1157–1166. [Google Scholar] [CrossRef]
- Weng, L.; Zhang, L.; Peng, Y.; Huang, R.S. Pharmacogenetics and pharmacogenomics: A bridge to individualized cancer therapy. Pharmacogenomics 2013, 14, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Henricks, L.M.; Lunenburg, C.; de Man, F.M.; Meulendijks, D.; Frederix, G.W.J.; Kienhuis, E.; Creemers, G.J.; Baars, A.; Dezentje, V.O.; Imholz, A.L.T.; et al. DPYD genotype-guided dose individualisation of fluoropyrimidine therapy in patients with cancer: A prospective safety analysis. Lancet Oncol. 2018, 19, 1459–1467. [Google Scholar] [CrossRef]
- Lunenburg, C.; Henricks, L.M.; Guchelaar, H.J.; Swen, J.J.; Deenen, M.J.; Schellens, J.H.M.; Gelderblom, H. Prospective DPYD genotyping to reduce the risk of fluoropyrimidine-induced severe toxicity: Ready for prime time. Eur. J. Cancer 2016, 54, 40–48. [Google Scholar] [CrossRef]
- Vreken, P.; Van Kuilenburg, A.B.; Meinsma, R.; Smit, G.P.; Bakker, H.D.; De Abreu, R.A.; van Gennip, A.H. A point mutation in an invariant splice donor site leads to exon skipping in two unrelated Dutch patients with dihydropyrimidine dehydrogenase deficiency. J. Inherit. Metab. Dis. 1996, 19, 645–654. [Google Scholar] [CrossRef]
- Hishinuma, E.; Narita, Y.; Saito, S.; Maekawa, M.; Akai, F.; Nakanishi, Y.; Yasuda, J.; Nagasaki, M.; Yamamoto, M.; Yamaguchi, H.; et al. Functional Characterization of 21 Allelic Variants of Dihydropyrimidine Dehydrogenase Identified in 1070 Japanese Individuals. Drug Metab. Dispos. 2018, 46, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Naushad, S.M.; Hussain, T.; Alrokayan, S.A.; Kutala, V.K. Pharmacogenetic profiling of dihydropyrimidine dehydrogenase (DPYD) variants in the Indian population. J. Gene Med. 2021, 23, e3289. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, A.S.; McMillin, G.A.; Patel, G.K.; Awan, S.; Taylor, W.R.; Pai, S.; Frankel, A.E.; Nelson, C.; Wang, B.; Hosein, P.J.; et al. The Prevalence of DPYD*9A(c.85T>C) Genotype and the Genotype-Phenotype Correlation in Patients with Gastrointestinal Malignancies Treated with Fluoropyrimidines: Updated Analysis. Clin. Colorectal. Cancer 2019, 18, e280–e286. [Google Scholar] [CrossRef]
- Terrazzino, S.; Cargnin, S.; Del Re, M.; Danesi, R.; Canonico, P.L.; Genazzani, A.A. DPYD IVS14+1G>A and 2846A>T genotyping for the prediction of severe fluoropyrimidine-related toxicity: A meta-analysis. Pharmacogenomics 2013, 14, 1255–1272. [Google Scholar] [CrossRef]
- Leung, H.W.; Chan, A.L. Association and prediction of severe 5-fluorouracil toxicity with dihydropyrimidine dehydrogenase gene polymorphisms: A meta-analysis. Biomed. Rep. 2015, 3, 879–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toffoli, G.; Giodini, L.; Buonadonna, A.; Berretta, M.; De Paoli, A.; Scalone, S.; Miolo, G.; Mini, E.; Nobili, S.; Lonardi, S.; et al. Clinical validity of a DPYD-based pharmacogenetic test to predict severe toxicity to fluoropyrimidines. Int. J. Cancer 2015, 137, 2971–2980. [Google Scholar] [CrossRef]
- Loganayagam, A.; Arenas Hernandez, M.; Corrigan, A.; Fairbanks, L.; Lewis, C.M.; Harper, P.; Maisey, N.; Ross, P.; Sanderson, J.D.; Marinaki, A.M. Pharmacogenetic variants in the DPYD, TYMS, CDA and MTHFR genes are clinically significant predictors of fluoropyrimidine toxicity. Br. J. Cancer 2013, 108, 2505–2515. [Google Scholar] [CrossRef] [Green Version]
- Meulendijks, D.; Henricks, L.M.; Sonke, G.S.; Deenen, M.J.; Froehlich, T.K.; Amstutz, U.; Largiader, C.R.; Jennings, B.A.; Marinaki, A.M.; Sanderson, J.D.; et al. Clinical relevance of DPYD variants c.1679T>G, c.1236G>A/HapB3, and c.1601G>A as predictors of severe fluoropyrimidine-associated toxicity: A systematic review and meta-analysis of individual patient data. Lancet Oncol. 2015, 16, 1639–1650. [Google Scholar] [CrossRef]
- Lunenburg, C.; van der Wouden, C.H.; Nijenhuis, M.; Crommentuijn-van Rhenen, M.H.; de Boer-Veger, N.J.; Buunk, A.M.; Houwink, E.J.F.; Mulder, H.; Rongen, G.A.; van Schaik, R.H.N.; et al. Dutch Pharmacogenetics Working Group (DPWG) guideline for the gene-drug interaction of DPYD and fluoropyrimidines. Eur. J. Hum. Genet. 2020, 28, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Amstutz, U.; Henricks, L.M.; Offer, S.M.; Barbarino, J.; Schellens, J.H.M.; Swen, J.J.; Klein, T.E.; McLeod, H.L.; Caudle, K.E.; Diasio, R.B.; et al. Clinical Pharmacogenetics Implementation Consortium (CPIC) Guideline for Dihydropyrimidine Dehydrogenase Genotype and Fluoropyrimidine Dosing: 2017 Update. Clin. Pharmacol. Ther. 2018, 103, 210–216. [Google Scholar] [CrossRef]
- Nie, Q.; Shrestha, S.; Tapper, E.E.; Trogstad-Isaacson, C.S.; Bouchonville, K.J.; Lee, A.M.; Wu, R.; Jerde, C.R.; Wang, Z.; Kubica, P.A.; et al. Quantitative Contribution of rs75017182 to Dihydropyrimidine Dehydrogenase mRNA Splicing and Enzyme Activity. Clin. Pharmacol. Ther. 2017, 102, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Amstutz, U.; Farese, S.; Aebi, S.; Largiader, C.R. Dihydropyrimidine dehydrogenase gene variation and severe 5-fluorouracil toxicity: A haplotype assessment. Pharmacogenomics 2009, 10, 931–944. [Google Scholar] [CrossRef] [Green Version]
- Ridge, S.A.; Sludden, J.; Wei, X.; Sapone, A.; Brown, O.; Hardy, S.; Canney, P.; Fernandez-Salguero, P.; Gonzalez, F.J.; Cassidy, J.; et al. Dihydropyrimidine dehydrogenase pharmacogenetics in patients with colorectal cancer. Br. J. Cancer 1998, 77, 497–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.R.; Diasio, R.B. Importance of dihydropyrimidine dehydrogenase (DPD) deficiency in patients exhibiting toxicity following treatment with 5-fluorouracil. Adv. Enzyme Regul. 2001, 41, 151–157. [Google Scholar] [CrossRef]
- Van Kuilenburg, A.B.; Haasjes, J.; Richel, D.J.; Zoetekouw, L.; van Lenthe, H.; De Abreu, R.A.; Maring, J.G.; Vreken, P.; van Gennip, A.H. Clinical implications of dihydropyrimidine dehydrogenase (DPD) deficiency in patients with severe 5-fluorouracil-associated toxicity: Identification of new mutations in the DPD gene. Clin. Cancer Res. 2000, 6, 4705–4712. [Google Scholar]
- Johnson, M.R.; Wang, K.; Diasio, R.B. Profound dihydropyrimidine dehydrogenase deficiency resulting from a novel compound heterozygote genotype. Clin. Cancer Res. 2002, 8, 768–774. [Google Scholar] [PubMed]
- Schwab, M.; Zanger, U.M.; Marx, C.; Schaeffeler, E.; Klein, K.; Dippon, J.; Kerb, R.; Blievernicht, J.; Fischer, J.; Hofmann, U.; et al. Role of genetic and nongenetic factors for fluorouracil treatment-related severe toxicity: A prospective clinical trial by the German 5-FU Toxicity Study Group. J. Clin. Oncol. 2008, 26, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.; Busse, B.; Riemenschneider, M.; Neubauer, S.; Seck, K.; Klein, H.G.; Kiechle, M.; Lordick, F.; Meindl, A. Strong association of a common dihydropyrimidine dehydrogenase gene polymorphism with fluoropyrimidine-related toxicity in cancer patients. PLoS ONE 2008, 3, e4003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popejoy, A.B. Diversity In Precision Medicine And Pharmacogenetics: Methodological And Conceptual Considerations For Broadening Participation. Pharmgenom. Pers. Med. 2019, 12, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.C.; Tanoshima, R.; Ross, C.J.D.; Carleton, B.C. Challenges and Opportunities in Implementing Pharmacogenetic Testing in Clinical Settings. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 65–84. [Google Scholar] [CrossRef]
- Zhang, H.; De, T.; Zhong, Y.; Perera, M.A. The Advantages and Challenges of Diversity in Pharmacogenomics: Can Minority Populations Bring Us Closer to Implementation? Clin. Pharmacol. Ther. 2019, 106, 338–349. [Google Scholar] [CrossRef]
- Radouani, F.; Zass, L.; Hamdi, Y.; Rocha, J.D.; Sallam, R.; Abdelhak, S.; Ahmed, S.; Azzouzi, M.; Benamri, I.; Benkahla, A.; et al. A review of clinical pharmacogenetics Studies in African populations. Pers. Med. 2020, 17, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Kurtz, G.; Parra, E.J. Population Diversity in Pharmacogenetics: A Latin American Perspective. Adv. Pharmacol. 2018, 83, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Phillips, C.; Pinto, N.; Santos, C.; dos Santos, S.E.; Amorim, A.; Carracedo, A.; Gusmao, L. Straightforward inference of ancestry and admixture proportions through ancestry-informative insertion deletion multiplexing. PLoS ONE 2012, 7, e29684. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Balloux, F.; Lugon-Moulin, N. The estimation of population differentiation with microsatellite markers. Mol. Ecol. 2002, 11, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Magne, N.; Etienne-Grimaldi, M.C.; Cals, L.; Renee, N.; Formento, J.L.; Francoual, M.; Milano, G. Dihydropyrimidine dehydrogenase activity and the IVS14+1G>A mutation in patients developing 5FU-related toxicity. Br. J. Clin. Pharmacol. 2007, 64, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Morel, A.; Boisdron-Celle, M.; Fey, L.; Soulie, P.; Craipeau, M.C.; Traore, S.; Gamelin, E. Clinical relevance of different dihydropyrimidine dehydrogenase gene single nucleotide polymorphisms on 5-fluorouracil tolerance. Mol. Cancer Ther. 2006, 5, 2895–2904. [Google Scholar] [CrossRef] [Green Version]
- Ben Fredj, R.; Gross, E.; Chouchen, L.; B’Chir, F.; Ben Ahmed, S.; Neubauer, S.; Kiechle, M.; Saguem, S. Mutational spectrum of dihydropyrimidine dehydrogenase gene (DPYD) in the Tunisian population. C. R. Biol. 2007, 330, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Soria-Chacartegui, P.; Villapalos-Garcia, G.; Lopez-Fernandez, L.A.; Navares-Gomez, M.; Mejia-Abril, G.; Abad-Santos, F.; Zubiaur, P. Clinical Relevance of Novel Polymorphisms in the Dihydropyrimidine Dehydrogenase (DPYD) Gene in Patients with Severe Fluoropyrimidine Toxicity: A Spanish Case-Control Study. Pharmaceutics 2021, 13, 2036. [Google Scholar] [CrossRef] [PubMed]
- Meulendijks, D.; Henricks, L.M.; Jacobs, B.A.W.; Aliev, A.; Deenen, M.J.; de Vries, N.; Rosing, H.; van Werkhoven, E.; de Boer, A.; Beijnen, J.H.; et al. Pretreatment serum uracil concentration as a predictor of severe and fatal fluoropyrimidine-associated toxicity. Br. J. Cancer 2017, 116, 1415–1424. [Google Scholar] [CrossRef]
- Meulendijks, D.; Cats, A.; Beijnen, J.H.; Schellens, J.H. Improving safety of fluoropyrimidine chemotherapy by individualizing treatment based on dihydropyrimidine dehydrogenase activity—Ready for clinical practice? Cancer Treat. Rev. 2016, 50, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Buchel, B.; Rhyn, P.; Schurch, S.; Buhr, C.; Amstutz, U.; Largiader, C.R. LC-MS/MS method for simultaneous analysis of uracil, 5,6-dihydrouracil, 5-fluorouracil and 5-fluoro-5,6-dihydrouracil in human plasma for therapeutic drug monitoring and toxicity prediction in cancer patients. Biomed. Chromatogr. 2013, 27, 7–16. [Google Scholar] [CrossRef]
- Vreken, P.; Van Kuilenburg, A.B.; Meinsma, R.; van Gennip, A.H. Identification of novel point mutations in the dihydropyrimidine dehydrogenase gene. J. Inherit. Metab. Dis. 1997, 20, 335–338. [Google Scholar] [CrossRef]
- Van Kuilenburg, A.B.; De Abreu, R.A.; van Gennip, A.H. Pharmacogenetic and clinical aspects of dihydropyrimidine dehydrogenase deficiency. Ann. Clin. Biochem. 2003, 40, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joerger, M.; Huitema, A.D.; Boot, H.; Cats, A.; Doodeman, V.D.; Smits, P.H.; Vainchtein, L.; Rosing, H.; Meijerman, I.; Zueger, M.; et al. Germline TYMS genotype is highly predictive in patients with metastatic gastrointestinal malignancies receiving capecitabine-based chemotherapy. Cancer Chemother. Pharmacol. 2015, 75, 763–772. [Google Scholar] [CrossRef]
- Khushman, M.; Patel, G.K.; Hosein, P.J.; Laurini, J.A.; Cameron, D.; Clarkson, D.R.; Butler, T.W.; Norden, C.W.; Baliem, W.; Jones, V.; et al. Germline pharmacogenomics of DPYD*9A (c.85T>C) variant in patients with gastrointestinal malignancies treated with fluoropyrimidines. J. Gastrointest. Oncol. 2018, 9, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Madi, A.; Fisher, D.; Maughan, T.S.; Colley, J.P.; Meade, A.M.; Maynard, J.; Humphreys, V.; Wasan, H.; Adams, R.A.; Idziaszczyk, S.; et al. Pharmacogenetic analyses of 2183 patients with advanced colorectal cancer; potential role for common dihydropyrimidine dehydrogenase variants in toxicity to chemotherapy. Eur. J. Cancer 2018, 102, 31–39. [Google Scholar] [CrossRef]
- Etienne-Grimaldi, M.C.; Boyer, J.C.; Beroud, C.; Mbatchi, L.; van Kuilenburg, A.; Bobin-Dubigeon, C.; Thomas, F.; Chatelut, E.; Merlin, J.L.; Pinguet, F.; et al. New advances in DPYD genotype and risk of severe toxicity under capecitabine. PLoS ONE 2017, 12, e0175998. [Google Scholar] [CrossRef] [PubMed]
- Hamzic, S.; Scharer, D.; Offer, S.M.; Meulendijks, D.; Nakas, C.; Diasio, R.B.; Fontana, S.; Wehrli, M.; Schurch, S.; Amstutz, U.; et al. Haplotype structure defines effects of common DPYD variants c.85T > C (rs1801265) and c.496A > G (rs2297595) on dihydropyrimidine dehydrogenase activity: Implication for 5-fluorouracil toxicity. Br. J. Clin. Pharmacol 2021, 87, 3234–3243. [Google Scholar] [CrossRef] [PubMed]
- Varma, K.A.; Jayanthi, M.; Dubashi, B.; Shewade, D.G. Influence of DPYD*9A, DPYD*6 and GSTP1 ile105val Genetic Polymorphisms on Capecitabine and Oxaliplatin (CAPOX) Associated Toxicities in Colorectal Cancer (CRC) Patients. Asian Pac. J. Cancer Prev. 2019, 20, 3093–3100. [Google Scholar] [CrossRef] [PubMed]
- Detailleur, S.; Segelov, E.; Re, M.D.; Prenen, H. Dihydropyrimidine dehydrogenase deficiency in patients with severe toxicity after 5-fluorouracil: A retrospective single-center study. Ann. Gastroenterol. 2021, 34, 68–72. [Google Scholar] [CrossRef]
- Homburger, J.R.; Moreno-Estrada, A.; Gignoux, C.R.; Nelson, D.; Sanchez, E.; Ortiz-Tello, P.; Pons-Estel, B.A.; Acevedo-Vasquez, E.; Miranda, P.; Langefeld, C.D.; et al. Genomic Insights into the Ancestry and Demographic History of South America. PLoS Genet. 2015, 11, e1005602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarga, C.M. Origen y Evolución de las Comunidades Afroamericanas de Esmeraldas (Ecuador): Datos Históricos y Genéticos. In Proceedings of the Anales del Museo de América. 1997, pp. 129–138. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=2016518/ (accessed on 5 August 2021).
- Lauderbaugh, G. The History of Ecuador; ABC-CLIO: Santa Barbara, CA, USA, 2012. [Google Scholar]
- INEC (Instituto Nacional de Estadísticas y Censos). Proyecciones Poblacionales. Available online: http://www.ecuador-encifras.gob.ec/proyecciones-poblacionales/ (accessed on 11 August 2021).
- Nagar, S.D.; Conley, A.B.; Chande, A.T.; Rishishwar, L.; Sharma, S.; Marino-Ramirez, L.; Aguinaga-Romero, G.; Gonzalez-Andrade, F.; Jordan, I.K. Genetic ancestry and ethnic identity in Ecuador. HGG Adv. 2021, 2, 100050. [Google Scholar] [CrossRef]
- Santangelo, R.; Gonzalez-Andrade, F.; Borsting, C.; Torroni, A.; Pereira, V.; Morling, N. Analysis of ancestry informative markers in three main ethnic groups from Ecuador supports a trihybrid origin of Ecuadorians. Forensic Sci. Int. Genet. 2017, 31, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, A.K.; Gaviria, A.; Cobos-Navarrete, S.; Gruezo, C.; Rodriguez-Pollit, C.; Armendariz-Castillo, I.; Garcia-Cardenas, J.M.; Guerrero, S.; Lopez-Cortes, A.; Leone, P.E.; et al. The three-hybrid genetic composition of an Ecuadorian population using AIMs-InDels compared with autosomes, mitochondrial DNA and Y chromosome data. Sci. Rep. 2019, 9, 9247. [Google Scholar] [CrossRef] [Green Version]
- Reich, D.; Patterson, N.; Campbell, D.; Tandon, A.; Mazieres, S.; Ray, N.; Parra, M.V.; Rojas, W.; Duque, C.; Mesa, N.; et al. Reconstructing Native American population history. Nature 2012, 488, 370–374. [Google Scholar] [CrossRef]
- Phan, L.; Jin, Y.; Zhang, H.; Qiang, W.; Shekhtman, E.; Shao, D.; Revoe, D.; Villamarin, R.; Ivanchenko, E.; Kimura, M.; et al. ALFA: Allele Frequency Aggregator; US National Library of Medicine: Bethesda, MD, USA, 2020; Volume 10. [Google Scholar]
- Suarez-Kurtz, G. Population impact of pharmacogenetic tests in admixed populations across the Americas. Pharmacogenom. J. 2021, 21, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Botton, M.R.; Hentschke-Lopes, M.; Matte, U. Frequency of DPYD gene variants and phenotype inference in a Southern Brazilian population. Ann. Hum. Genet. 2021, 86, 102–107. [Google Scholar] [CrossRef]
- Goljan, E.; Abouelhoda, M.; ElKalioby, M.M.; Jabaan, A.; Alghithi, N.; Meyer, B.F.; Monies, D. Identification of pharmacogenetic variants from large scale next generation sequencing data in the Saudi population. PLoS ONE 2022, 17, e0263137. [Google Scholar] [CrossRef]
- Zhou, Y.; Lauschke, V.M. Population pharmacogenomics: An update on ethnogeographic differences and opportunities for precision public health. Hum. Genet. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zintzaras, E.; Lau, J. Synthesis of genetic association studies for pertinent gene-disease associations requires appropriate methodological and statistical approaches. J. Clin. Epidemiol. 2008, 61, 634–645. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farinango, C.; Gallardo-Cóndor, J.; Freire-Paspuel, B.; Flores-Espinoza, R.; Jaramillo-Koupermann, G.; López-Cortés, A.; Burgos, G.; Tejera, E.; Cabrera-Andrade, A. Genetic Variations of the DPYD Gene and Its Relationship with Ancestry Proportions in Different Ecuadorian Trihybrid Populations. J. Pers. Med. 2022, 12, 950. https://doi.org/10.3390/jpm12060950
Farinango C, Gallardo-Cóndor J, Freire-Paspuel B, Flores-Espinoza R, Jaramillo-Koupermann G, López-Cortés A, Burgos G, Tejera E, Cabrera-Andrade A. Genetic Variations of the DPYD Gene and Its Relationship with Ancestry Proportions in Different Ecuadorian Trihybrid Populations. Journal of Personalized Medicine. 2022; 12(6):950. https://doi.org/10.3390/jpm12060950
Chicago/Turabian StyleFarinango, Camila, Jennifer Gallardo-Cóndor, Byron Freire-Paspuel, Rodrigo Flores-Espinoza, Gabriela Jaramillo-Koupermann, Andrés López-Cortés, Germán Burgos, Eduardo Tejera, and Alejandro Cabrera-Andrade. 2022. "Genetic Variations of the DPYD Gene and Its Relationship with Ancestry Proportions in Different Ecuadorian Trihybrid Populations" Journal of Personalized Medicine 12, no. 6: 950. https://doi.org/10.3390/jpm12060950