
Multi-Omics Analysis Reveals Myelin, Presynaptic and Nicotinate Alterations in the Hippocampus of G72/G30 Transgenic Mice

Abstract
:1. Introduction
2. Materials and Methods
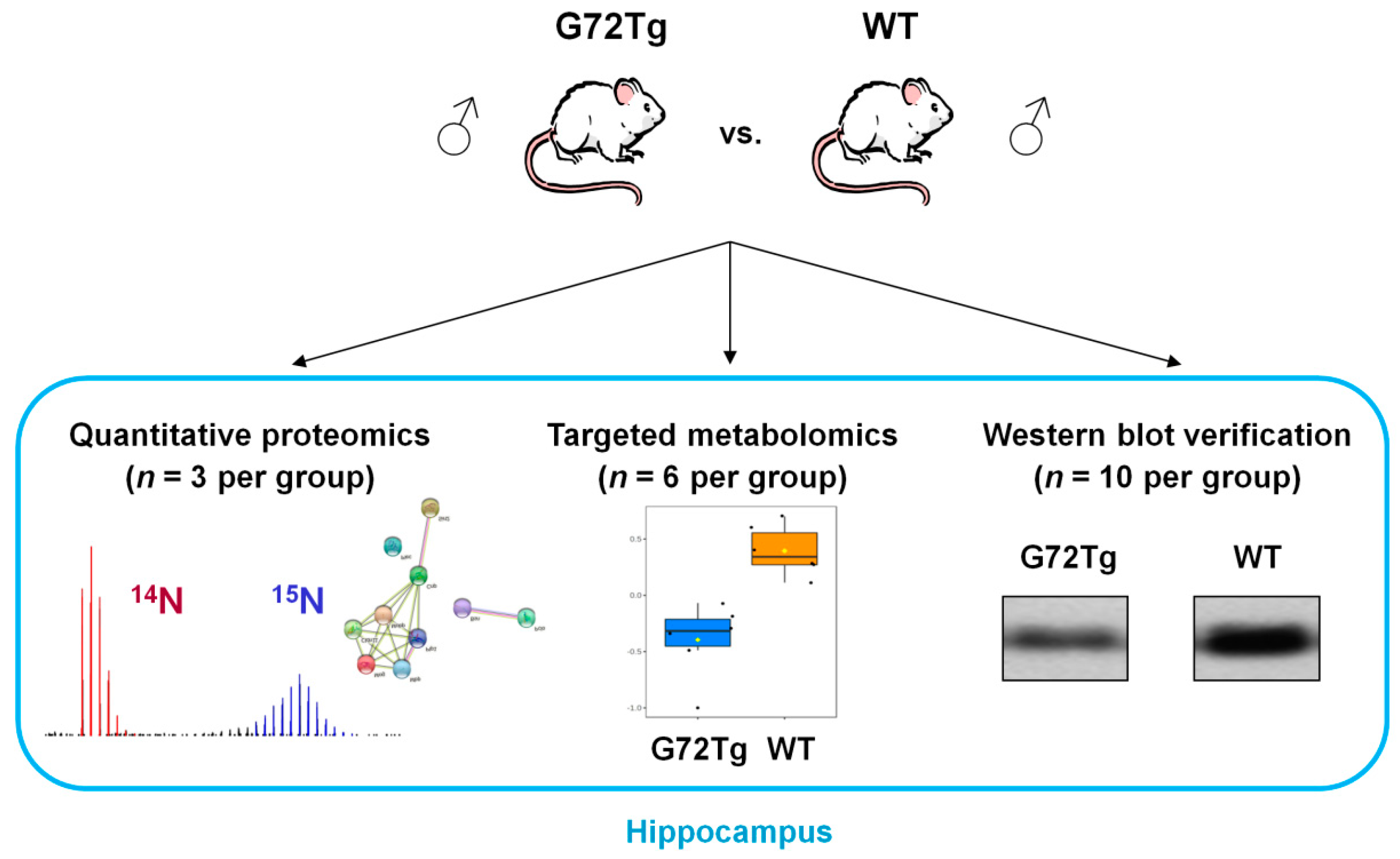
2.1. Experimental Design
2.2. Transgenic Mice Carrying the G72/G30 Locus (G72Tg) and Wild-Type (WT) Mice
2.3. 15N Metabolic Labeling of CD1 Mice
2.4. Sample Collection
2.5. Quantitative Proteomics Sample Preparation and Measurement
2.6. Quantitative Proteomics Data Analysis
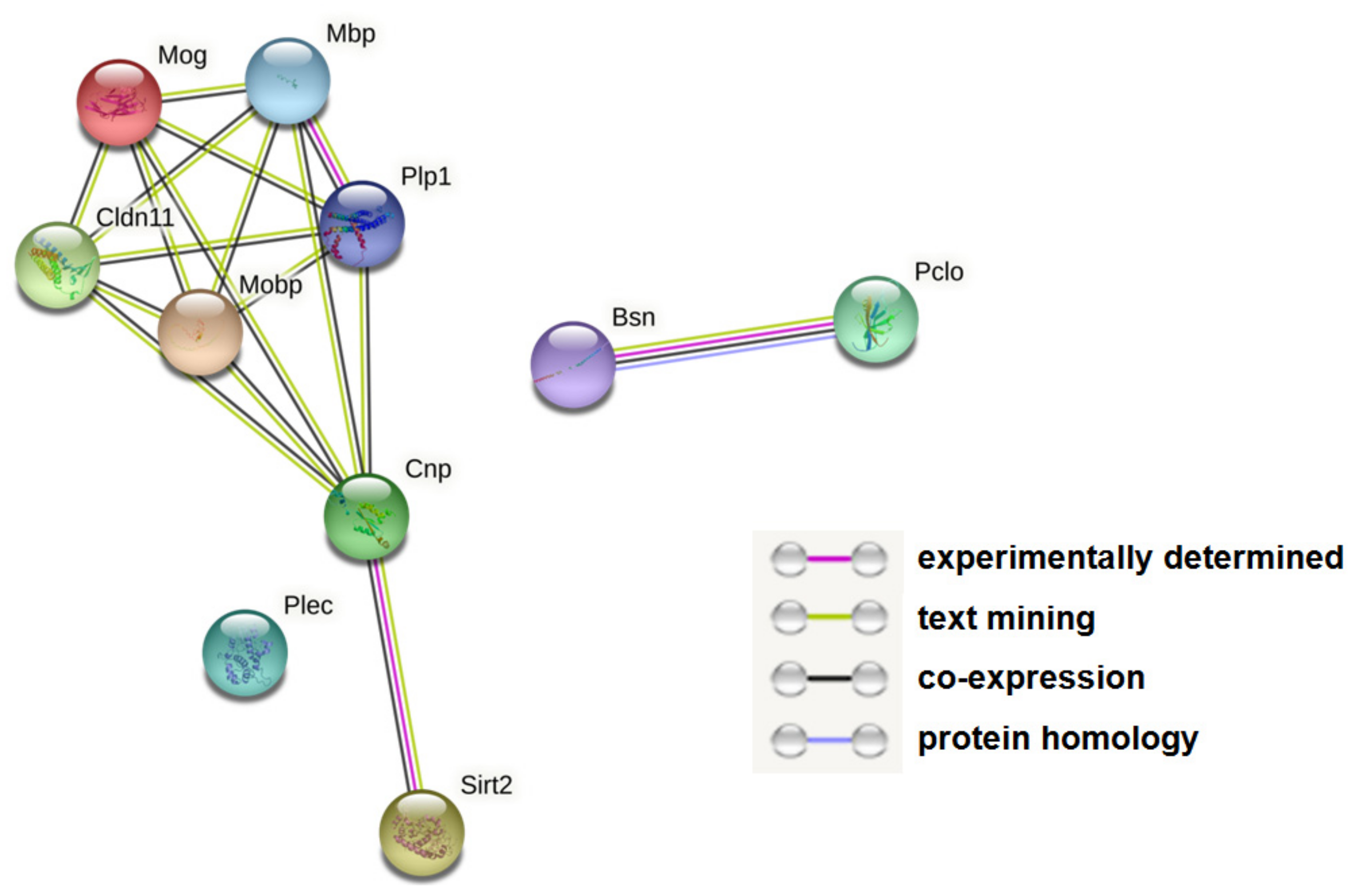
2.7. Protein Network Visualization
2.8. Targeted Metabolomics
2.9. Targeted Metabolomics Data Analysis
2.10. Western Blot
3. Results
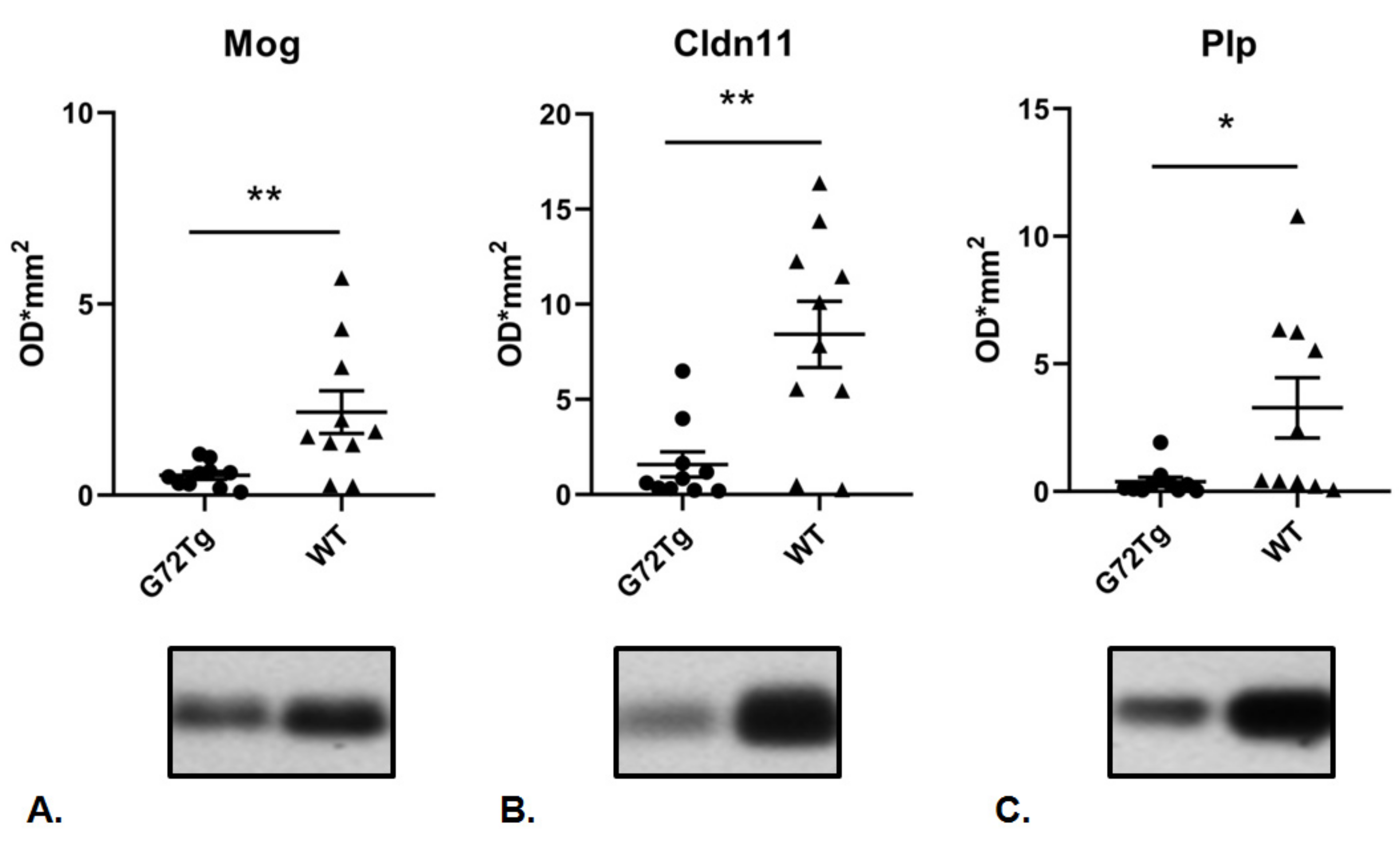
3.1. Decreased Expression of Myelin-Related Proteins and Increased Expression of Presynaptic Proteins in G72Tg Hippocampi
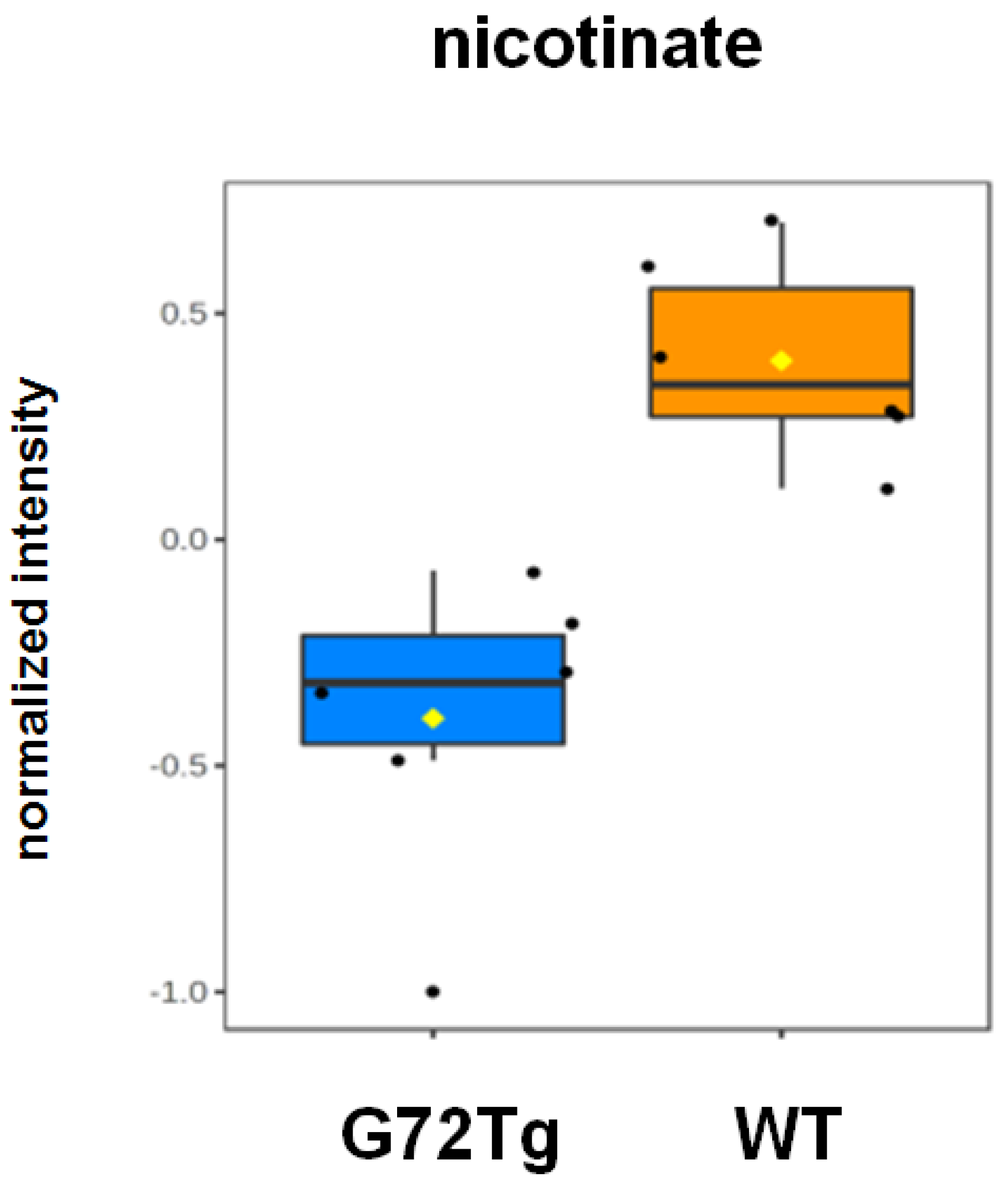
3.2. Decreased Nicotinate Levels in G72Tg Hippocampi
3.3. Convergent Protein Expression Changes in Hippocampus and Cerebellum in G72Tg Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chumakov, I.; Blumenfeld, M.; Guerassimenko, O.; Cavarec, L.; Palicio, M.; Abderrahim, H.; Bougueleret, L.; Barry, C.; Tanaka, H.; La Rosa, P.; et al. Genetic and physiological data implicating the new human gene G72 and the gene for D-amino acid oxidase in schizophrenia. Proc. Natl. Acad. Sci. USA 2002, 99, 13675–13680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detera-Wadleigh, S.D.; McMahon, F.J. G72/G30 in schizophrenia and bipolar disorder: Review and meta-analysis. Biol. Psychiatry 2006, 60, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, S.; Binelli, G.; Pollegioni, L. G72 primate-specific gene: A still enigmatic element in psychiatric disorders. Cell. Mol. Life Sci. 2016, 73, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Akyol, E.S.; Albayrak, Y.; Aksoy, N.; Sahin, B.; Beyazyuz, M.; Kuloglu, M.; Hashimoto, K. Increased serum G72 protein levels in patients with schizophrenia: A potential candidate biomarker. Acta Neuropsychiatr. 2017, 29, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Korostishevsky, M.; Kaganovich, M.; Cholostoy, A.; Ashkenazi, M.; Ratner, Y.; Dahary, D.; Bernstein, J.; Bening-Abu-Shach, U.; Ben Asher, E.; Lancet, D.; et al. Is the G72/G30 locus associated with schizophrenia? single nucleotide polymorphisms, haplotypes, and gene expression analysis. Biol. Psychiatry 2004, 56, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Otte, D.M.; Bilkei-Gorzo, A.; Filiou, M.D.; Turck, C.W.; Yilmaz, O.; Holst, M.I.; Schilling, K.; Abou-Jamra, R.; Schumacher, J.; Benzel, I.; et al. Behavioral changes in G72/G30 transgenic mice. Eur. Neuropsychopharmacol. 2009, 19, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Otte, D.M.; Sommersberg, B.; Kudin, A.; Guerrero, C.; Albayram, O.; Filiou, M.D.; Frisch, P.; Yilmaz, O.; Drews, E.; Turck, C.W.; et al. N-acetyl cysteine treatment rescues cognitive deficits induced by mitochondrial dysfunction in G72/G30 transgenic mice. Neuropsychopharmacology 2011, 36, 2233–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filiou, M.D.; Turck, C.W. Psychiatric disorder biomarker discovery using quantitative proteomics. Methods Mol. Biol. 2012, 829, 531–539. [Google Scholar] [PubMed]
- Filiou, M.D.; Teplytska, L.; Otte, D.M.; Zimmer, A.; Turck, C.W. Myelination and oxidative stress alterations in the cerebellum of the G72/G30 transgenic schizophrenia mouse model. J. Psychiatr. Res. 2012, 46, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Frank, E.; Kessler, M.S.; Filiou, M.D.; Zhang, Y.; Maccarrone, G.; Reckow, S.; Bunck, M.; Heumann, H.; Turck, C.W.; Landgraf, R.; et al. Stable isotope metabolic labeling with a novel N-enriched bacteria diet for improved proteomic analyses of mouse models for psychopathologies. PLoS ONE 2009, 4, e7821. [Google Scholar] [CrossRef] [PubMed]
- Filiou, M.D.; Zhang, Y.; Teplytska, L.; Reckow, S.; Gormanns, P.; Maccarrone, G.; Frank, E.; Kessler, M.S.; Hambsch, B.; Nussbaumer, M.; et al. Proteomics and metabolomics analysis of a trait anxiety mouse model reveals divergent mitochondrial pathways. Biol. Psychiatry 2011, 70, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Filiou, M.D.; Bisle, B.; Reckow, S.; Teplytska, L.; Maccarrone, G.; Turck, C.W. Profiling of mouse synaptosome proteome and phosphoproteome by IEF. Electrophoresis 2010, 31, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, G.; Chen, A.; Filiou, M.D. Using 15N-Metabolic Labeling for Quantitative Proteomic Analyses. Methods Mol. Biol. 2017, 1546, 235–243. [Google Scholar]
- Deutsch, E.W.; Mendoza, L.; Shteynberg, D.; Farrah, T.; Lam, H.; Tasman, N.; Sun, Z.; Nilsson, E.; Pratt, B.; Prazen, B.; et al. A guided tour of the Trans-Proteomic Pipeline. Proteomics 2010, 10, 1150–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filiou, M.D.; Varadarajulu, J.; Teplytska, L.; Reckow, S.; Maccarrone, G.; Turck, C.W. The 15N isotope effect in Escherichia coli: A neutron can make the difference. Proteomics 2012, 12, 3121–3128. [Google Scholar] [CrossRef] [PubMed]
- Filiou, M.D.; Asara, J.M.; Nussbaumer, M.; Teplytska, L.; Landgraf, R.; Turck, C.W. Behavioral extremes of trait anxiety in mice are characterized by distinct metabolic profiles. J. Psychiatr. Res. 2014, 58, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Webhofer, C.; Gormanns, P.; Reckow, S.; Lebar, M.; Maccarrone, G.; Ludwig, T.; Pütz, B.; Asara, J.M.; Holsboer, F.; Sillaber, I.; et al. Proteomic and metabolomic profiling reveals time-dependent changes in hippocampal metabolism upon paroxetine treatment and biomarker candidates. J. Psychiatr. Res. 2013, 47, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Breitkopf, S.B.; Yang, X.M.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.E.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J. The hippocampus in schizophrenia: A review of the neuropathological evidence and its pathophysiological implications. Psychopharmacology 2004, 174, 151–162. [Google Scholar] [CrossRef]
- Haukvik, U.K.; Westlye, L.T.; Morch-Johnsen, L.; Jorgensen, K.N.; Lange, E.H.; Dale, A.M.; Melle, I.; Andreassen, O.A.; Agartz, I. In vivo hippocampal subfield volumes in schizophrenia and bipolar disorder. Biol. Psychiatry 2015, 77, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, D.A.; Eijsink, V.D.; Spoelder, M.; van Hulten, J.A.; De Weerd, P.; Homberg, J.R.; Valles, A.; Nait-Oumesmar, B.; Martens, G.J.M. Interneuron hypomyelination is associated with cognitive inflexibility in a rat model of schizophrenia. Nat. Commun. 2020, 11, 2329. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, L.P.; Yarnykh, V.L.; Parshukova, D.A.; Kornetova, E.G.; Semke, A.V.; Usova, A.V.; Pishchelko, A.O.; Khodanovich, M.Y.; Ivanova, S.A. Global hypomyelination of the brain white and gray matter in schizophrenia: Quantitative imaging using macromolecular proton fraction. Transl. Psychiatry 2021, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Maccarrone, G.; Wobrock, T.; Zerr, I.; Gormanns, P.; Reckow, S.; Falkai, P.; Schmitt, A.; Turck, C.W. Proteome analysis of the thalamus and cerebrospinal fluid reveals glycolysis dysfunction and potential biomarkers candidates for schizophrenia. J. Psychiatr. Res. 2010, 44, 1176–1189. [Google Scholar] [CrossRef]
- Saia-Cereda, V.M.; Cassoli, J.S.; Schmitt, A.; Falkai, P.; Nascimento, J.M.; Martins-de-Souza, D. Proteomics of the corpus callosum unravel pivotal players in the dysfunction of cell signaling, structure, and myelination in schizophrenia brains. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 601–612. [Google Scholar] [CrossRef]
- Martins-de-Souza, D.; Gattaz, W.F.; Schmitt, A.; Maccarrone, G.; Hunyadi-Gulyas, E.; Eberlin, M.N.; Souza, G.H.M.F.; Marangoni, S.; Novello, J.C.; Turck, C.W.; et al. Proteomic analysis of dorsolateral prefrontal cortex indicates the involvement of cytoskeleton, oligodendrocyte, energy metabolism and new potential markers in schizophrenia. J. Psychiatr. Res. 2009, 43, 978–986. [Google Scholar] [CrossRef]
- Marui, T.; Torii, Y.; Iritani, S.; Sekiguchi, H.; Habuchi, C.; Fujishiro, H.; Oshima, K.; Niizato, K.; Hayashida, S.; Masaki, K.; et al. The neuropathological study of myelin oligodendrocyte glycoprotein in the temporal lobe of schizophrenia patients. Acta Neuropsychiatr. 2018, 30, 232–240. [Google Scholar] [CrossRef]
- Falkai, P.; Malchow, B.; Wetzestein, K.; Nowastowski, V.; Bernstein, H.G.; Steiner, J.; Schneider-Axmann, T.; Kraus, T.; Hasan, A.; Bogerts, B.; et al. Decreased Oligodendrocyte and Neuron Number in Anterior Hippocampal Areas and the Entire Hippocampus in Schizophrenia: A Stereological Postmortem Study. Schizophr. Bull. 2016, 42 (Suppl. S1), S4–S12. [Google Scholar] [CrossRef]
- Wood, P.L.; Filiou, M.D.; Otte, D.M.; Zimmer, A.; Turck, C.W. Lipidomics reveals dysfunctional glycosynapses in schizophrenia and the G72/G30 transgenic mouse. Schizophr. Res. 2014, 159, 365–369. [Google Scholar] [CrossRef]
- Kedracka-Krok, S.; Swiderska, B.; Jankowska, U.; Skupien-Rabian, B.; Solich, J.; Dziedzicka-Wasylewska, M. Stathmin reduction and cytoskeleton rearrangement in rat nucleus accumbens in response to clozapine and risperidone treatment—Comparative proteomic study. Neuroscience 2016, 316, 63–81. [Google Scholar] [CrossRef]
- Maheras, K.J.; Peppi, M.; Ghoddoussi, F.; Galloway, M.P.; Perrine, S.A.; Gow, A. Absence of Claudin 11 in CNS Myelin Perturbs Behavior and Neurotransmitter Levels in Mice. Sci. Rep. 2018, 8, 3798. [Google Scholar] [CrossRef] [PubMed]
- Faludi, G.; Mirnics, K. Synaptic changes in the brain of subjects with schizophrenia. Int. J. Dev. Neurosci. 2011, 29, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egbujo, C.N.; Sinclair, D.; Hahn, C.G. Dysregulations of Synaptic Vesicle Trafficking in Schizophrenia. Curr. Psychiatry Rep. 2016, 18, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdenis van Berlekom, A.; Muflihah, C.H.; Snijders, G.; MacGillavry, H.D.; Middeldorp, J.; Hol, E.M.; Kahn, R.S.; De Witte, L.D. Synapse Pathology in Schizophrenia: A Meta-analysis of Postsynaptic Elements in Postmortem Brain Studies. Schizophr. Bull. 2020, 46, 374–386. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Huang, Y.S.; Liao, D.L.; Huang, C.Y.; Lin, C.H.; Fang, T.H. Identification of Rare Mutations of Two Presynaptic Cytomatrix Genes BSN and PCLO in Schizophrenia and Bipolar Disorder. J. Pers. Med. 2021, 11, 1057. [Google Scholar] [CrossRef]
- Weidenhofer, J.; Bowden, N.A.; Scott, R.J.; Tooney, P.A. Altered gene expression in the amygdala in schizophrenia: Up-regulation of genes located in the cytomatrix active zone. Mol. Cell. Neurosci. 2006, 31, 243–250. [Google Scholar] [CrossRef]
- Nitta, A.; Izuo, N.; Hamatani, K.; Inagaki, R.; Kusui, Y.; Fu, K.; Asano, T.; Torri, Y.; Habuchi, C.; Sekiguchi, H.; et al. Schizophrenia-Like Behavioral Impairments in Mice with Suppressed Expression of Piccolo in the Medial Prefrontal Cortex. J. Pers. Med. 2021, 11, 607. [Google Scholar] [CrossRef]
- Popiolek, M.; Ross, J.F.; Charych, E.; Chanda, P.; Gundelfinger, E.D.; Moss, S.J.; Brandon, N.J.; Pausch, M.H. D-amino acid oxidase activity is inhibited by an interaction with bassoon protein at the presynaptic active zone. J. Biol. Chem. 2011, 286, 28867–28875. [Google Scholar] [CrossRef] [Green Version]
- Turck, C.W.; Filiou, M.D. What Have Mass Spectrometry-Based Proteomics and Metabolomics (Not) Taught Us about Psychiatric Disorders? Mol. Neuropsychiatry 2015, 1, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Iliou, A.; Vlaikou, A.M.; Nussbaumer, M.; Benaki, D.; Mikros, E.; Gikas, E.; Filiou, M.D. Exploring the metabolomic profile of cerebellum after exposure to acute stress. Stress 2021, 24, 952–964. [Google Scholar] [CrossRef]
- Filiou, M.D.; Nussbaumer, M.; Teplytska, L.; Turck, C.W. Behavioral and Metabolome Differences between C57BL/6 and DBA/2 Mouse Strains: Implications for Their Use as Models for Depression- and Anxiety-Like Phenotypes. Metabolites 2021, 11, 128. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, Z.; Vlaikou, A.M.; Theodoridou, D.; Komini, C.; Chalkiadaki, G.; Vafeiadi, M.; Margetaki, K.; Trangas, K.; Turck, C.W.; Syrrou, M.; et al. Unraveling the Serum Metabolomic Profile of Post-partum Depression. Front. Neurosci. 2019, 13, 833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Filiou, M.D.; Reckow, S.; Gormanns, P.; Maccarrone, G.; Kessler, M.S.; Frank, E.; Hambsch, B.; Holsboer, F.; Landgraf, R.; et al. Proteomic and metabolomic profiling of a trait anxiety mouse model implicate affected pathways. Mol. Cell. Proteom. 2011, 10, M111.008110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chousidis, I.; Chatzimitakos, T.; Leonardos, D.; Filiou, M.D.; Stalikas, C.D.; Leonardos, I.D. Cannabinol in the spotlight: Toxicometabolomic study and behavioral analysis of zebrafish embryos exposed to the unknown cannabinoid. Chemosphere 2020, 252, 126417. [Google Scholar] [CrossRef]
- Weckmann, K.; Deery, M.J.; Howard, J.A.; Feret, R.; Asara, J.M.; Dethloff, F.; Filiou, M.D.; Labermaier, C.; Maccarrone, M.; Lilley, K.S. Ketamine’s Effects on the Glutamatergic and GABAergic Systems: A Proteomics and Metabolomics Study in Mice. Mol. Neuropsychiatry 2019, 5, 42–51. [Google Scholar] [CrossRef]
- Weckmann, K.; Deery, M.J.; Howard, J.A.; Feret, R.; Asara, J.M.; Dethloff, F.; Filiou, M.D.; Iannace, J.; Labermaier, C.; Maccarrone, G.; et al. Ketamine’s antidepressant effect is mediated by energy metabolism and antioxidant defense system. Sci. Rep. 2017, 7, 15788. [Google Scholar] [CrossRef] [Green Version]
- Park, D.I.; Dournes, C.; Sillaber, I.; Ising, M.; Asara, J.M.; Webhofer, C.; Filiou, M.D.; Müller, M.B.; Turck, C.W. Delineation of molecular pathway activities of the chronic antidepressant treatment response suggests important roles for glutamatergic and ubiquitin-proteasome systems. Transl. Psychiatry 2017, 7, e1078. [Google Scholar] [CrossRef] [Green Version]
- Park, D.I.; Dournes, C.; Sillaber, I.; Uhr, M.; Asara, J.M.; Gassen, N.C.; Rein, T.; Ising, M.; Webhofer, C.; Filiou, M.D.; et al. Purine and pyrimidine metabolism: Convergent evidence on chronic antidepressant treatment response in mice and humans. Sci. Rep. 2016, 6, 35317. [Google Scholar] [CrossRef] [Green Version]
- Nussbaumer, M.; Asara, J.M.; Teplytska, L.; Murphy, M.P.; Logan, A.; Turck, C.W.; Filiou, M.D. Selective Mitochondrial Targeting Exerts Anxiolytic Effects In Vivo. Neuropsychopharmacology 2016, 41, 1751–1758. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.Y.; He, Z.; Henes, K.; Asara, J.M.; Webhofer, C.; Filiou, M.D.; Khaitovich, P.; Wotjak, C.T.; Turck, C.W. Fluoxetine Treatment Rescues Energy Metabolism Pathway Alterations in a Posttraumatic Stress Disorder Mouse Model. Mol. Neuropsychiatry 2016, 2, 46–59. [Google Scholar] [CrossRef]
- Periyasamy, S.; John, S.; Padmavati, R.; Rajendren, P.; Thirunavukkarasu, P.; Gratten, J.; Vinkhuizen, A.; McRae, A.; Holliday, E.G.; Nyholt, D.R. Association of Schizophrenia Risk With Disordered Niacin Metabolism in an Indian Genome-wide Association Study. JAMA Psychiatry 2019, 76, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Messamore, E. The niacin response biomarker as a schizophrenia endophenotype: A status update. Prostaglandins Leukot. Essent. Fat. Acids 2018, 136, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Otte, D.M.; Raskó, T.; Wang, M.; Dreiseidler, M.; Drews, E.; Schrage, H.; Wojtalla, A.; Hohfeld, J.; Wanker, E.; Zimmer, A. Identification of the mitochondrial MSRB2 as a binding partner of LG72. Cell. Mol. Neurobiol. 2014, 34, 1123–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filiou, M.D.; Sandi, C. Anxiety and Brain Mitochondria: A Bidirectional Crosstalk. Trends Neurosci. 2019, 42, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Wallace, D.C. Mitochondrial Etiology of Neuropsychiatric Disorders. Biol. Psychiatry 2018, 83, 722–730. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Protein Name | G72Tg/WT Fold Change | Adjusted p-Value | Protein Groups |
|---|---|---|---|---|
| Decreased expression in G72Tg hippocampi | ||||
| Mobp | Myelin-associated oligodendrocyte basic protein | 0.50 | 4.88409 × 10−5 | MOBP_MOUSE |
| Plp1 | Myelin proteolipid protein | 0.54 | 8.06522 × 10−81 | Q3UYM8_MOUSE, MYPR_MOUSE |
| Cldn11 (Osp) | Claudin-11, oligodentrocyte-specific protein | 0.54 | 1.72045 × 10−9 | CLD11_MOUSE |
| Mbp | Myelin basic protein | 0.57 | 2.16222 × 10−24 | Q542T4_MOUSE |
| Mog | Myelin oligodentrocyte glycoprotein | 0.57 | 1.26402 × 10−9 | MOG_MOUSE, Q80YU5_MOUSE, Q3UY21_MOUSE |
| Cnp | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase | 0.66 | 8.69626 × 10−23 | CN37_MOUSE, Q3TYV5_MOUSE |
| Sirt2 | NAD-dependent protein deacetylase sirtuin-2 | 0.71 | 0.001562104 | SIRT2_MOUSE, Q3UJK6_MOUSE |
| Increased expression in G72Tg hippocampi | ||||
| Bsn | Protein bassoon | 1.41 | 1.34696 × 10−8 | BSN_MOUSE |
| Plec1 | Plectin | 1.41 | 0.015085062 | Q6S387_MOUSE, Q6S390_MOUSE, Q6S388_MOUSE, Q6S385_MOUSE, Q6S392_MOUSE, PLEC1_MOUSE, Q6S393_MOUSE |
| Pclo | Protein piccolo | 1.74 | 0.00012123 | PCLO_MOUSE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filiou, M.D.; Teplytska, L.; Nussbaumer, M.; Otte, D.-M.; Zimmer, A.; Turck, C.W. Multi-Omics Analysis Reveals Myelin, Presynaptic and Nicotinate Alterations in the Hippocampus of G72/G30 Transgenic Mice. J. Pers. Med. 2022, 12, 244. https://doi.org/10.3390/jpm12020244
Filiou MD, Teplytska L, Nussbaumer M, Otte D-M, Zimmer A, Turck CW. Multi-Omics Analysis Reveals Myelin, Presynaptic and Nicotinate Alterations in the Hippocampus of G72/G30 Transgenic Mice. Journal of Personalized Medicine. 2022; 12(2):244. https://doi.org/10.3390/jpm12020244
Chicago/Turabian StyleFiliou, Michaela D., Larysa Teplytska, Markus Nussbaumer, David-M. Otte, Andreas Zimmer, and Christoph W. Turck. 2022. "Multi-Omics Analysis Reveals Myelin, Presynaptic and Nicotinate Alterations in the Hippocampus of G72/G30 Transgenic Mice" Journal of Personalized Medicine 12, no. 2: 244. https://doi.org/10.3390/jpm12020244