
Altered Cerebral Processing of Videos in Children with Motor Dysfunction Suggests Broad Embodiment of Perceptual Cognitive Functions
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Clinical Assessment of Patients
2.3. Cognitive Assessment of Patients
2.4. Stimuli
2.5. EEG Data Collection and Preprocessing
2.6. Correlated Components Analysis
2.7. Alpha Power Estimation
2.8. Automatic Annotation of Movements
3. Results
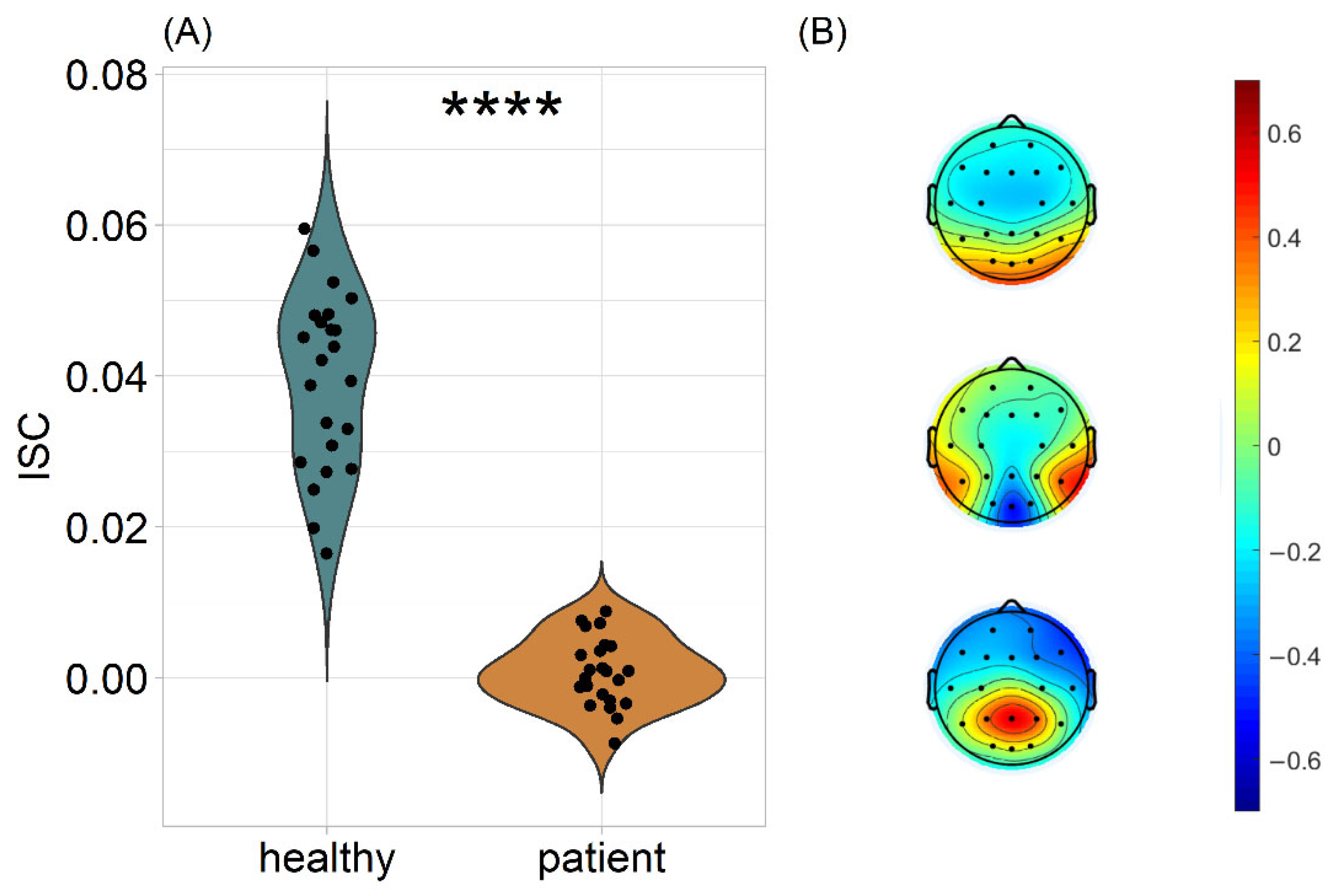
3.1. Patients’ Neural Responses Are Dissimilar to Those of Healthy Controls during Movie Watching
3.2. ISC Was Not Associated with the Diagnostic Status of the Patients
3.3. ISC Was Lower in Patients Irrespective of Video Clip Content
4. Discussion
5. Conclusions
- Patients with Arthrogryposis multiplex congenita or Obstetric brachial plexus palsy exhibit divergent neural responses to naturalistic videos
- The observed effect was not associated with the severity of the patients’ clinical condition
- The observed effect was not associated with the motor-related content of the stimuli, suggesting broad embodiment of perceptual cognitive functions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, M. Six views of embodied cognition. Psychon. Bull. Rev. 2002, 9, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.L. Embodied cognition: A field guide. Artif. Intell. 2003, 149, 91–130. [Google Scholar] [CrossRef]
- Foglia, L.; Wilson, R.A. Embodied cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2013, 4, 319–325. [Google Scholar] [CrossRef]
- Farina, M. Embodied cognition: Dimensions, domains and applications. Adapt. Behav. 2021, 29, 73–88. [Google Scholar] [CrossRef]
- Shapiro, L. The Embodied Cognition Research Programme. Philos. Compass 2007, 2, 338–346. [Google Scholar] [CrossRef]
- Clark, A. Supersizing the Mind: Embodiment, Action, and Cognitive Extension; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Wilson, A.D.; Golonka, S. Embodied cognition is not what you think it is. Front. Psychol. 2013, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- Mahon, B.Z. What is embodied about cognition? Lang. Cogn. Neurosci. 2015, 30, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Goldinger, S.D.; Papesh, M.H.; Barnhart, A.S.; Hansen, W.A.; Hout, M.C. The poverty of embodied cognition. Psychon. Bull. Rev. 2016, 23, 959–978. [Google Scholar] [CrossRef]
- Goldman, A.I.; de Vignemont, F. Is social cognition embodied? Trends Cogn. Sci. 2009, 13, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Mahon, B.Z.; Caramazza, A. A critical look at the embodied cognition hypothesis and a new proposal for grounding conceptual content. J. Physiol. 2008, 102, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. The ‘shared manifold’ hypothesis: From mirror neurons to empathy. J. Conscious. Stud. 2001, 8, 33–50. [Google Scholar]
- Gallese, V.; Lakoff, G. The brain’s concepts: The role of the sensorymotor system in conceptual knowledge. Cogn. Neuropsychol. 2005, 22, 455–479. [Google Scholar] [CrossRef]
- Gallese, V. Embodied simulation: From neurons to phenomenal experience. Phenomenol. Cogn. Sci. 2005, 22, 455–479. [Google Scholar] [CrossRef]
- Gallese, V. Before and below ‘theory of mind’: Embodied simulation and the neural correlates of social cognition. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Hasson, U.; Nir, Y.; Levy, I.; Fuhrmann, G.; Malach, R. Intersubject synchronization of cortical activity during natural vision. Science 2004, 303, 1634–1640. [Google Scholar] [PubMed]
- Dmochowski, J.P.; Sajda, P.; Dias, J.; Parra, L.C. Correlated components of ongoing EEG point to emotionally laden attention—A possible marker of engagement? Front. Hum. Neurosci. 2012, 6, 112. [Google Scholar] [CrossRef] [PubMed]
- Jääskeläinen, I.P.; Sams, M.; Glerean, E.; Ahveninen, J. Movies and narratives as naturalistic stimuli in neuroimaging. Neuroimage 2021, 224, 117445. [Google Scholar] [CrossRef] [PubMed]
- Oberman, L.M.; Winkielman, P.; Ramachandran, V.S. Face to face: Blocking facial mimicry can selectively impair recognition of emotional expressions. Soc. Neurosci. 2007, 2, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Ponari, M.; Conson, M.; D’Amico, N.P.; Grossi, D.; Trojano, L. Mapping correspondence between facial mimicry and emotion recognition in healthy subjects. Emotion 2012, 12, 1398–1403. [Google Scholar] [PubMed]
- Wołoszyn, K.; Hohol, M.; Kuniecki, M.; Winkielman, P. Restricting movements of lower face leaves recognition of emotional vocalizations intact but introduces a valence positivity bias. Sci. Rep. 2022, 12, 16101. [Google Scholar] [CrossRef]
- Buccino, G.; Dalla Volta, R.; Arabia, G.; Morelli, M.; Chiriaco, C.; Lupo, A.; Silipo, F.; Quattrone, A. Processing graspable object images and their nouns is impaired in Parkinson’s disease patients. Cortex 2018, 100, 32–39. [Google Scholar] [CrossRef]
- Beilock, S.L. Grounding cognition in action: Expertise, comprehension, and judgment. Prog. Brain Res. 2009, 174, 3–11. [Google Scholar] [PubMed]
- Walle, T.; Hartikainen-Sorri, A.L. Obstetric shoulder injury: Associated risk factors, prediction and prognosis. Acta Obstet. Gynecol. Scand. 1993, 72, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Pondaag, W.; Malessy, M.J.; van Dijk, J.G.; Thomeer, R.T. Natural history of obstetric brachial plexus palsy: A systematic review. Dev. Med. Child Neurol. 2004, 46, 138–144. [Google Scholar] [CrossRef]
- Bellew, M.; Kay, S.P.; Webb, F.; Ward, A. Developmental and behavioural outcome in obstetric brachial plexus palsy. J. Hand Surg. 2000, 25, 49–51. [Google Scholar] [CrossRef]
- Pearl, M.L.; Edgerton, B.W. Glenoid deformity secondary to brachial plexus birth palsy. J. Bone Jt. Surg. 1998, 80, 659–667. [Google Scholar] [CrossRef]
- Anguelova, G.V.; Malessy, M.J.A.; van Zwet, E.W.; van Dijk, J.G. Extensive motor axonal misrouting after conservative treatment of obstetric brachial plexus lesions. Dev. Med. Child Neurol. 2014, 56, 984–989. [Google Scholar] [CrossRef]
- Brown, T.; Cupido, C.; Scarfone, H.; Pape, K.; Galea, V.; McComas, A. Developmental apraxia arising from neonatal brachial plexus palsy. Neurology 2000, 55, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Anguelova, G.V.; Malessy, M.J.; Buitenhuis, S.M.; Zwet, E.W.; Dijk, J.G. Impaired Automatic Arm Movements in Obstetric Brachial Plexus Palsy Suggest a Central Disorder. J. Child Neurol. 2016, 31, 1005–1009. [Google Scholar] [CrossRef]
- Dahan-Oliel, N.; Cachecho, S.; Barnes, D.; Bedard, T.; Davison, A.M.; Dieterich, K.; Donohoe, M.; Fąfara, A.; Hamdy, R.; Hjartarson, H.T.; et al. International multidisciplinary collaboration toward an annotated definition of arthrogryposis multiplex congenita. Am. J. Med. Genet. Part C Semin. Med. Genet. 2019, 181, 288–299. [Google Scholar] [CrossRef]
- Oishi, S.; Agranovich, O.; Zlotolow, D.; Wall, L.; Stutz, C.; Pajardi, G.; Novelli, C.; Abdel Ghani, H.; Jester, A.; Vuillermin, C.; et al. Treatment and outcomes of arthrogryposis in the upper extremity. Am. J. Med. Genet. Part C Semin. Med. Genet. 2019, 181, 363–371. [Google Scholar] [CrossRef]
- Van Heest, A.; Waters, P.M.; Simmons, B.P. Surgical treatment of arthrogryposis of the elbow. J. Hand Surg. Am. 1998, 23, 1063–1070. [Google Scholar] [CrossRef]
- Oishi, S.N.; Agranovich, O.; Pajardi, G.E.; Novelli, C.; Baindurashvili, A.G.; Trofimova, S.I.; Abdel-Ghani, H.; Kochenova, E.; Prosperpio, G.; Jester, A.; et al. Treatment of the Upper Extremity Contracture/Deformities. J. Pediatr. Orthop. 2017, 37 (Suppl. 1), S9–S15. [Google Scholar] [CrossRef] [PubMed]
- Zafeiriou, D.I.; Psychogiou, K. Obstetrical brachial plexus palsy. Pediatr. Neurol. 2008, 38, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Craighero, L. The mirror-neuron system. Annu. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [CrossRef]
- Koriakina, M.; Agranovich, O.; Petrova, E.; Kadieva, D.; Kopytin, G.; Ermolovich, E.; Moiseenko, O.; Alekseeva, M.; Bredikhin, D.; Bermúdez-Margaretto, B.; et al. Aberrant Auditory and Visual Memory Development of Children with Upper Limb Motor Disorders. Brain Sci. 2021, 11, 1650. [Google Scholar] [CrossRef] [PubMed]
- Blagoveschenskiy, E.D.; Agranovich, O.E.; Kononova, E.L.; Baindurashvili, A.G.; Nazarova, M.A.; Shestokova, A.N.; Gabbasova, E.L.; Nikulin, V.V. Characteristics of electrophysiological activity of the cerebral cortex in children with arthrogryposis. Neuromuscul. Dis. 2018, 8, 10–17. [Google Scholar] [CrossRef]
- Dixon, M.L.; Girn, M.; Christoff, K. Hierarchical Organization of Frontoparietal Control Networks Underlying Goal-Directed Behavior. In The prefrontal Cortex as an Executive, Emotional, and Social Brain; Watanabe, M., Ed.; Springer: Tokyo, Japan, 2017. [Google Scholar]
- Nastase, S.A.; Gazzola, V.; Hasson, U.; Keysers, C. Measuring shared responses across subjects using intersubject correlation. Soc. Cogn. Affect. Neurosci. 2019, 14, 667–685. [Google Scholar] [CrossRef]
- Iotzov, I.; Fidali, B.C.; Petroni, A.; Conte, M.M.; Schiff, N.D.; Parra, L.C. Divergent neural responses to narrative speech in disorders of consciousness. Ann. Clin. Transl. Neurol. 2017, 4, 784–792. [Google Scholar] [CrossRef]
- Lankinen, K.; Saari, J.; Hari, R.; Koskinen, M. Intersubject consistency of cortical MEG signals during movie viewing. Neuroimage 2014, 92, 217–224. [Google Scholar] [CrossRef]
- Bezdek, M.A.; Wenzel, W.G.; Schumacher, E.H. The effect of visual and musical suspense on brain activation and memory during naturalistic viewing. Biol. Psychol. 2017, 129, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Lehne, M.; Engel, P.; Rohrmeier, M.; Menninghaus, W.; Jacobs, A.M.; Koelsch, S. Reading a suspenseful literary text activates brain areas related to social cognition and predictive inference. PLoS ONE 2015, 10, e0124550. [Google Scholar] [CrossRef]
- Wechsler, D.; Kodama, H. Wechsler Intelligence Scale for Children, 5th ed.; Psychological Corporation: New York, NY, USA, 2014. [Google Scholar]
- Shipitsyna, L. Psychological Diagnosis of Developmental Deviations in Children of Primary School Age, 1st ed.; Speech: Saint Petersburg, Russia, 2008. [Google Scholar]
- Cohen, S.S.; Madsen, J.; Touchan, G.; Robles, D.; Lima, S.F.; Henin, S.; Parra, L.C. Neural engagement with online educational videos predicts learning performance for individual students. Neurobiol. Learn. Mem. 2018, 155, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.S.; Parra, L.C. Memorable Audiovisual Narratives Synchronize Sensory and Supramodal Neural Responses. eNeuro 2016, 3, 203–216. [Google Scholar] [CrossRef]
- Hyvärinen, A. Fast and robust fixed-point algorithms for independent component analysis. IEEE Trans. Neural Netw. 1999, 10, 626–634. [Google Scholar]
- Parra, L.C.; Haufe, S.; Dmochowski, J.P. Correlated components analysis-extracting reliable dimensions in multivariate data. arXiv 2018, arXiv:1801.08881. Available online: https://arxiv.org/pdf/1801.08881.pdf (accessed on 10 July 2022).
- Dmochowski, J.P.; Bezdek, M.A.; Abelson, B.P.; Johnson, J.S.; Schumacher, E.H.; Parra, L.C. Audience preferences are predicted by temporal reliability of neural processing. Nat. Commun. 2014, 5, 4567. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. On the “probable error” of a coefficient of correlation deduced from a small sample. Metron 1921, 1, 3–32. [Google Scholar]
- Glerean, E.; Salmi, J.; Lahnakoski, J.M.; Jääskeläinen, I.P.; Sams, M. Functional magnetic resonance imaging phase synchronization as a measure of dynamic functional connectivity. Brain Connect. 2012, 2, 91–101. [Google Scholar] [CrossRef]
- Ki, J.J.; Kelly, S.P.; Parra, L.C. Attention Strongly Modulates Reliability of Neural Responses to Naturalistic Narrative Stimuli. J. Neurosci. 2016, 36, 3092–3101. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Hidalgo, G.; Simon, T.; Wei, S.-E.; Sheikh, Y. OpenPose: Realtime Multi-Person 2D Pose Estimation using Part Affinity Fields. arXiv 2019, arXiv:1812.08008. Available online: https://arxiv.org/abs/1812.08008 (accessed on 10 July 2022).
- Ray, W.J.; Cole, H.W. EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Khamis, H. Measures of Association: How to Choose? J. Diagn. Med. Sonogr. 2008, 24, 155–162. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar]
- Gallese, V. Bodily selves in relation: Embodied simulation as secondperson perspective on intersubjectivity. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130177. [Google Scholar]
- Petroni, A.; Cohen, S.S.; Ai, L.; Langer, N.; Henin, S.; Vanderwal, T.; Milham, M.P.; Parra, L.C. The Variability of Neural Responses to Naturalistic Videos Change with Age and Sex. eNeuro 2018, 5, ENEURO.0244-17.2017. [Google Scholar]
- Kostenetskiy, P.S.; Chulkevich, R.A.; Kozyrev, V.I. HPC Resources of the Higher School of Economics. J. Phys. Conf. Ser. 2021, 1740, 012050. [Google Scholar]



{kind=link}
{kind=link}
| Code | Age | Sex | Diagnosis | Paresis | CNS Damage | DMD | GMD | Attention | Auditory Memory | Visual Memory | Story |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | 8 | F | OBPL | 4 | 3 | 4 | 7 | 5 | 5 | 6 | 4 |
| P2 | 6 | M | AMC | 3 | 2 | 3 | 5 | 1 | 3 | 4 | 2 |
| P3 | 15 | F | OBPL | 4 | 3 | 3 | 6 | 3 | 6 | 3 | 4 |
| P4 | 5 | F | AMC | 1 | 3 | 3 | 6 | 3 | 3 | 6 | 3 |
| P5 | 9 | F | OBPL | 4 | 3 | 3 | 6 | 3 | 5 | 3 | 4 |
| P6 | 7 | M | AMC | 1 | 2 | 2 | 6 | 3 | 4 | 2 | 4 |
| P7 | 4 | M | AMC | 1 | 3 | 2 | 5 | 4 | 3 | 3 | 4 |
| P8 | 8 | F | AMC | 1 | 2 | 1 | 3 | 2 | 3 | 3 | 3 |
| P9 | 13 | M | OBPL | 4 | 3 | 4 | 12 | 5 | 7 | 10 | 2 |
| P10 | 5 | M | OBPL | 4 | 4 | 4 | 13 | 3 | 5 | 6 | 1 |
| P11 | 4 | F | AMC | 1 | 3 | 2 | 5 | 3 | 5 | 5 | 3 |
| P12 | 10 | F | OBPL | 4 | 3 | 4 | 11 | 3 | 7 | 6 | 2 |
| P13 | 8 | F | AMC | 1 | 2 | 3 | 5 | 2 | 4 | 3 | 4 |
| P14 | 4 | F | AMC | 1 | 1 | 1 | 2 | 1 | 1 | 4 | 1 |
| P15 | 8 | M | AMC | 1 | 2 | 2 | 4 | 2 | 4 | 5 | 3 |
| P16 | 8 | M | OBPL | 4 | 3 | 4 | 12 | 3 | 6 | 6 | 3 |
| P17 | 15 | M | AMC | 1 | 2 | 2 | 4 | 4 | 6 | 7 | 3 |
| P18 | 5 | M | AMC | 1 | 2 | 2 | 4 | 2 | 3 | 2 | 1 |
| P19 | 9 | F | AMC | 1 | 1 | 2 | 3 | 2 | 5 | 6 | 3 |
| P20 | 14 | M | OBPL | 4 | 3 | 4 | 12 | 5 | 7 | 9 | 4 |
| P21 | 8 | M | AMC | 1 | 2 | 1 | 3 | 2 | 3 | 5 | 3 |
| P22 | 5 | M | AMC | 1 | 3 | 3 | 8 | 3 | 5 | 2 | 2 |
| P23 | 9 | F | AMC | 1 | 3 | 3 | 6 | 3 | 7 | 4 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntoumanis, I.; Agranovich, O.; Shestakova, A.N.; Blagovechtchenski, E.; Koriakina, M.; Kadieva, D.; Kopytin, G.; Jääskeläinen, I.P. Altered Cerebral Processing of Videos in Children with Motor Dysfunction Suggests Broad Embodiment of Perceptual Cognitive Functions. J. Pers. Med. 2022, 12, 1841. https://doi.org/10.3390/jpm12111841
Ntoumanis I, Agranovich O, Shestakova AN, Blagovechtchenski E, Koriakina M, Kadieva D, Kopytin G, Jääskeläinen IP. Altered Cerebral Processing of Videos in Children with Motor Dysfunction Suggests Broad Embodiment of Perceptual Cognitive Functions. Journal of Personalized Medicine. 2022; 12(11):1841. https://doi.org/10.3390/jpm12111841
Chicago/Turabian StyleNtoumanis, Ioannis, Olga Agranovich, Anna N. Shestakova, Evgeny Blagovechtchenski, Maria Koriakina, Dzerassa Kadieva, Grigory Kopytin, and Iiro P. Jääskeläinen. 2022. "Altered Cerebral Processing of Videos in Children with Motor Dysfunction Suggests Broad Embodiment of Perceptual Cognitive Functions" Journal of Personalized Medicine 12, no. 11: 1841. https://doi.org/10.3390/jpm12111841