
Differential Expression Profiles of Cell-to-Matrix-Related Molecules in Adrenal Cortical Tumors: Diagnostic and Prognostic Implications

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. PCR-Based Gene Expression Profile
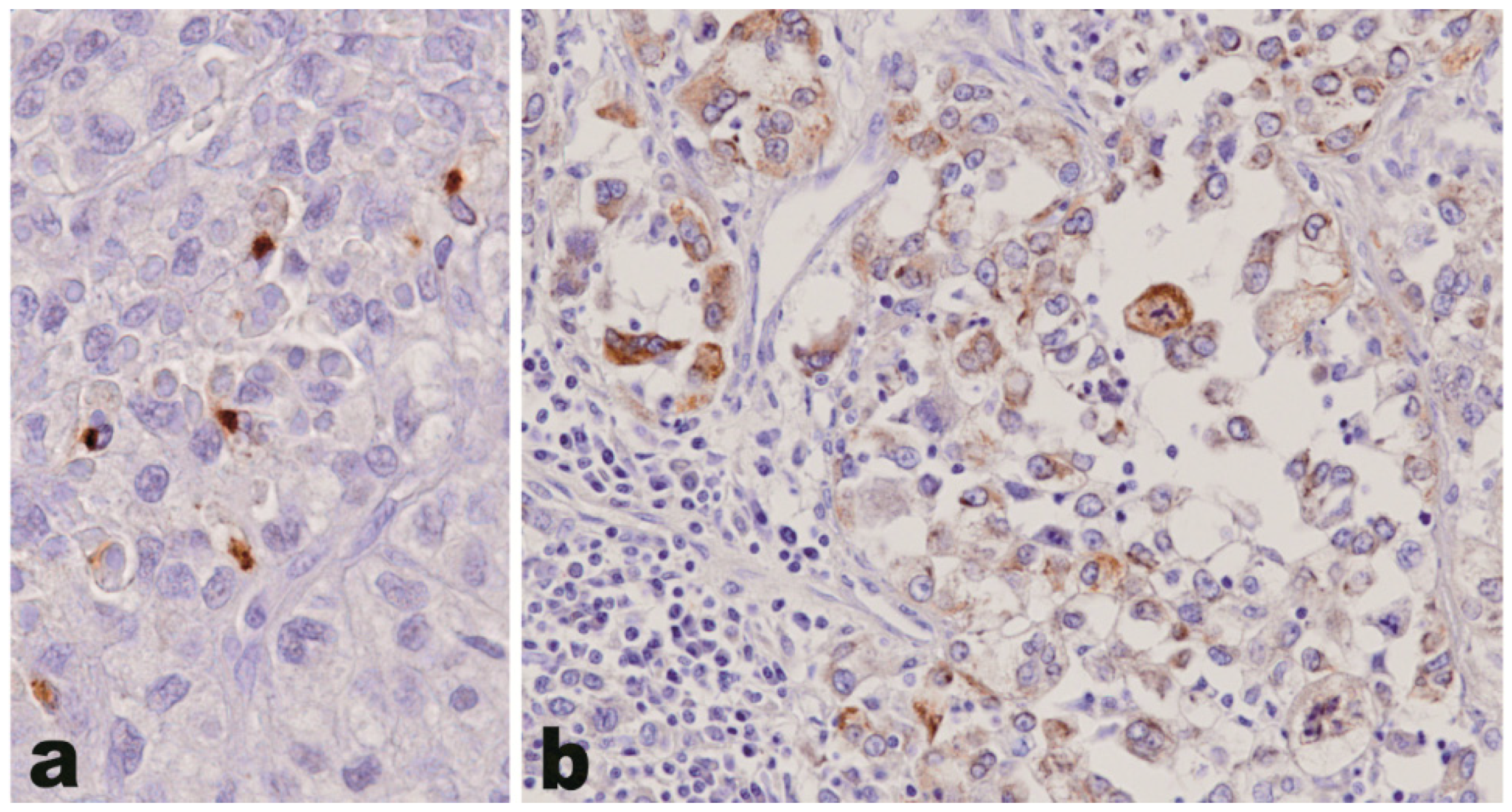
2.2. Osteopontin, Laminin α3 and HAS-1 Protein Expression in Adrenocortical Tumors: Case Selection and Immunohistochemistry
2.3. Statistical Analysis
3. Results
3.1. Gene Expression Profile of Cell-Matrix Interactive and Angiogenic Proteins in Adrenocortical Carcinomas as Compared to Adenomas
3.2. Osteopontin, Laminin α3 and HAS-1 Protein Expression in Adrenocortical Tumors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Classification of Tumours of Endocrine Organs; Lloyd, R.V.; Osamura, R.Y.; Klöppel, G.; Rosai, J. (Eds.) IARC (International Agency for Research on Cancer), Scientific Publications: Lyon, France, 2017; Volume 10, pp. 163–173. [Google Scholar]
- Giordano, T.J.; Berney, D.; de Krijger, R.R.; Erickson, L.; Fassnacht, M.; Mete, O.; Papathomas, T.; Papotti, M.; Sasano, H.; Thompson, L.D.R.; et al. Data set for reporting of carcinoma of the adrenal cortex: Explanations and recommendations of the guidelines from the International Collaboration on Cancer Reporting. Hum. Pathol. 2020. [Google Scholar] [CrossRef]
- Duregon, E.; Fassina, A.; Volante, M.; Nesi, G.; Santi, R.; Gatti, G.; Cappellesso, R.; Dalino Ciaramella, P.; Ventura, L.; Gambacorta, M.; et al. The reticulin algorithm for adrenocortical tumor diagnosis: A multicentric validation study on 245 unpublished cases. Am. J. Surg. Pathol. 2013, 37, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Duregon, E.; Cappellesso, R.; Maffeis, V.; Zaggia, B.; Ventura, L.; Berruti, A.; Terzolo, M.; Fassina, A.; Volante, M.; Papotti, M. Validation of the prognostic role of the “Helsinki Score” in 225 cases of adrenocortical carcinoma. Hum. Pathol. 2017, 62, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pilon, C.; Pistorello, M.; Moscon, A.; Altavilla, G.; Pagotto, U.; Boscaro, M.; Fallo, F. Inactivation of the p16 tumor suppressor gene in adrenocortical tumors. J. Clin. Endocrinol. Metab. 1999, 84, 2776–2779. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.M.; Billerbeck, A.E.C.; Fragoso, M.C.B.V.; Mendonca, B.B.; Latronico, A.C. Deletion mapping of chromosome 17 in benign and malignant adrenocortical tumors associated with the Arg337His mutation of the p53 tumor suppressor protein. J. Clin. Endocrinol. Metab. 2005, 90, 2976–2981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojadinovic, A.; Ghossein, R.A.; Hoos, A.; Nissan, A.; Marshall, D.; Dudas, M.; Cordon-Cardo, C.; Jaques, D.P.; Brennan, M.F. Adrenocortical carcinoma: Clinical, morphologic, and molecular characterization. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Aubert, S.; Wacrenier, A.; Leroy, X.; Devos, P.; Carnaille, B.; Proye, C.; Wemeau, J.L.; Lecomte-Houcke, M.; Leteurtre, E. Weiss system revisited. Am. J. Surg. Pathol. 2002, 26, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Terzolo, M.; Boccuzzi, A.; Bovio, S.; Cappia, S.; De Giuli, P.; Alì, A.; Paccotti, P.; Porpiglia, F.; Fontana, D.; Angeli, A. Immunohistochemical assessment of Ki-67 in the differential diagnosis of adrenocortical tumors. Urology 2001, 57, 176–182. [Google Scholar] [CrossRef]
- Papathomas, T.G.; Pucci, E.; Giordano, T.J.; Lu, H.; Duregon, E.; Volante, M.; Papotti, M.; Lloyd, R.V.; Tischler, A.S.; van Nederveen, F.H.; et al. An International Ki67 reproducibility study in adrenal cortical carcinoma. Am. J. Surg. Pathol. 2016, 40, 569–576. [Google Scholar] [CrossRef]
- Duregon, E.; Molinaro, L.; Volante, M.; Ventura, L.; Righi, L.; Bolla, S.; Terzolo, M.; Sapino, A.; Papotti, M.G. Comparative diagnostic and prognostic performances of the hematoxylin-eosin and phospho-histone H3 mitotic count and Ki-67 index in adrenocortical carcinoma. Mod. Pathol. 2014, 27, 1246–1254. [Google Scholar] [CrossRef] [Green Version]
- Beuschlein, F.; Weigel, J.; Saeger, W.; Kroiss, M.; Wild, V.; Daffara, F.; Libé, R.; Ardito, A.; Al Ghuzlan, A.; Quinkler, M.; et al. Major prognostic role of Ki67 in localized adrenocortical carcinoma after complete resection. J. Clin. Endocrinol. Metab. 2015, 100, 841–849. [Google Scholar] [CrossRef]
- Assié, G.; Jouinot, A.; Fassnacht, M.; Libé, R.; Garinet, S.; Jacob, L.; Hamzaoui, N.; Neou, M.; Sakat, J.; de La Villéon, B.; et al. Value of molecular classification for prognostic assessment of adrenocortical carcinoma. JAMA Oncol. 2019, 5. [Google Scholar] [CrossRef]
- Assié, G.; Letouzé, E.; Fassnacht, M.; Jouinot, A.; Luscap, W.; Barreau, O.; Omeiri, H.; Rodriguez, S.; Perlemoine, K.; René-Corail, F.; et al. Integrated genomic characterization of adrenocortical carcinoma. Nat. Genet. 2014, 46, 607–612. [Google Scholar] [CrossRef]
- Guo, J.; Gu, Y.; Ma, X.; Zhang, L.; Li, H.; Yan, Z.; Han, Y.; Xie, L.; Guo, X. Identification of hub genes and pathways in adrenocortical carcinoma by integrated bioinformatic analysis. J. Cell. Mol. Med. 2020, 24, 4428–4438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Cherniack, A.D.; Dewal, N.; Moffitt, R.A.; Danilova, L.; Murray, B.A.; Lerario, A.M.; Else, T.; Knijnenburg, T.A.; Ciriello, G.; et al. Comprehensive pan-genomic characterization of adrenocortical carcinoma. Cancer Cell 2016, 29, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, T.J.; Thomas, D.G.; Kuick, R.; Lizyness, M.; Misek, D.E.; Smith, A.L.; Sanders, D.; Aljundi, R.T.; Gauger, P.G.; Thompson, N.W.; et al. Distinct transcriptional profiles of adrenocortical tumors uncovered by DNA microarray analysis. Am. J. Pathol. 2003, 162, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Gicquel, C.; Raffin-Sanson, M.L.; Gaston, V.; Bertagna, X.; Plouin, P.F.; Schlumberger, M.; Louvel, A.; Luton, J.P.; Le Bouc, Y. Structural and functional abnormalities at 11p15 are associated with the malignant phenotype in sporadic adrenocortical tumors: Study on a series of 82 tumors. J. Clin. Endocrinol. Metab. 1997, 82, 2559–2565. [Google Scholar] [CrossRef] [PubMed]
- De Fraipont, F.; El Atifi, M.; Cherradi, N.; Le Moigne, G.; Defaye, G.; Houlgatte, R.; Bertherat, J.; Bertagna, X.; Plouin, P.F.; Baudin, E.; et al. Gene expression profiling of human adrenocortical tumors using complementary deoxyribonucleic acid microarrays identifies several candidate genes as markers of malignancy. J. Clin. Endocrinol. Metab. 2005, 90, 1819–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velázquez-Fernández, D.; Laurell, C.; Geli, J.; Höög, A.; Odeberg, J.; Kjellman, M.; Lundeberg, J.; Hamberger, B.; Nilsson, P.; Bäckdahl, M. Expression profiling of adrenocortical neoplasms suggests a molecular signature of malignancy. Surgery 2005, 138, 1087–1094. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.; Saremaslani, P.; Schmid, S.; Rousson, V.; Montani, M.; Schmid, D.M.; Heitz, P.U.; Komminoth, P.; Perren, A. IGFII and MIB1 immunohistochemistry is helpful for the differentiation of benign from malignant adrenocortical tumours. Histopathology 2006, 49, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Agosta, C.; Laugier, J.; Guyon, L.; Denis, J.; Bertherat, J.; Libé, R.; Boisson, B.; Sturm, N.; Feige, J.J.; Chabre, O.; et al. MiR-483-5p and miR-139-5p promote aggressiveness by targeting N-myc downstream-regulated gene family members in adrenocortical cancer. Int. J. Cancer 2018, 143, 944–957. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; He, W.; Chen, P.; Xu, D.; Zeng, G.; Li, Z.; Huang, M.; Wang, X.; DiSanto, M.E.; Zhang, X. Identification of seven aberrantly methylated and expressed genes in adrenocortical carcinoma. Front. Endocrinol. 2019, 10, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buishand, F.O.; Liu-Chittenden, Y.; Fan, Y.; Tirosh, A.; Gara, S.K.; Patel, D.; Meerzaman, D.; Kebebew, E. Adrenocortical tumors have a distinct, long, non-coding RNA expression profile and LINC00271 is downregulated in malignancy. Surgery 2020, 167, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Altieri, B.; Sbiera, S.; Della Casa, S.; Weigand, I.; Wild, V.; Steinhauer, S.; Fadda, G.; Kocot, A.; Bekteshi, M.; Mambretti, E.M.; et al. Livin/BIRC7 expression as malignancy marker in adrenocortical tumors. Oncotarget 2017, 8, 9323–9338. [Google Scholar] [CrossRef] [Green Version]
- Ronchi, C.L.; Sbiera, S.; Altieri, B.; Steinhauer, S.; Wild, V.; Bekteshi, M.; Kroiss, M.; Fassnacht, M.; Allolio, B. Notch1 pathway in adrenocortical carcinomas: Correlations with clinical outcome. Endocr. Relat. Cancer 2015, 22, 531–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronchi, C.L.; Sbiera, S.; Volante, M.; Steinhauer, S.; Scott-Wild, V.; Altieri, B.; Kroiss, M.; Bala, M.; Papotti, M.; Deutschbein, T.; et al. CYP2W1 is highly expressed in adrenal glands and is positively associated with the response to mitotane in adrenocortical carcinoma. PLoS ONE 2014, 9, e105855. [Google Scholar] [CrossRef] [PubMed]
- Kjellman, M.; Enberg, U.; Höög, A.; Larsson, C.; Holst, M.; Farnebo, L.O.; Sato, H.; Bäckdahl, M. Gelatinase A and membrane-type 1 matrix metalloproteinase mRNA: Expressed in adrenocortical cancers but not in adenomas. World J. Surg. 1999, 23, 237–242. [Google Scholar] [CrossRef]
- Kolomecki, K.; Stepien, H.; Bartos, M.; Kuzdak, K. Usefulness of VEGF, MMP-2, MMP-3 and TIMP-2 serum level evaluation in patients with adrenal tumours. Endocr. Regul. 2001, 35, 9–16. [Google Scholar]
- De Fraipont, F.; El Atifi, M.; Gicquel, C.; Bertagna, X.; Chambaz, E.M.; Feige, J.J. Expression of the angiogenesis markers vascular endothelial growth factor-A, thrombospondin-1, and platelet-derived endothelial cell growth factor in human sporadic adrenocortical tumors: Correlation with genotypic alterations. J. Clin. Endocrinol. Metab. 2000, 85, 4734–4741. [Google Scholar] [CrossRef]
- Bernini, G.P.; Moretti, A.; Bonadio, A.G.; Menicagli, M.; Viacava, P.; Naccarato, A.G.; Iacconi, P.; Miccoli, P.; Salvetti, A. Angiogenesis in human normal and pathologic adrenal cortex. J. Clin. Endocrinol. Metab. 2002, 87, 4961–4965. [Google Scholar] [CrossRef] [Green Version]
- Geervliet, E.; Bansal, R. Matrix metalloproteinases as potential biomarkers and therapeutic targets in liver diseases. Cells 2020, 9, 1212. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, L.S. Review: Emerging treatment strategies for adrenocortical carcinoma: A new hope. J. Clin. Endocrinol. Metab. 2006, 91, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volante, M.; Sperone, P.; Bollito, E.; Frangipane, E.; Rosas, R.; Daffara, F.; Terzolo, M.; Berruti, A.; Papotti, M. Matrix metalloproteinase type 2 expression in malignant adrenocortical tumors: Diagnostic and prognostic significance in a series of 50 adrenocortical carcinomas. Mod. Pathol. 2006, 19, 1563–1569. [Google Scholar] [CrossRef] [Green Version]
- Weismann, D.; Briese, J.; Niemann, J.; Grüneberger, M.; Adam, P.; Hahner, S.; Johanssen, S.; Liu, W.; Ezzat, S.; Saeger, W.; et al. Osteopontin stimulates invasion of NCI-h295 cells but is not associated with survival in adrenocortical carcinoma. J. Pathol. 2009, 218, 232–240. [Google Scholar] [CrossRef]
- Sapino, A.; Marchiò, C.; Senetta, R.; Castellano, I.; Macrì, L.; Cassoni, P.; Ghisolfi, G.; Cerrato, M.; D’Ambrosio, E.; Bussolati, G. Routine assessment of prognostic factors in breast cancer using a multicore tissue microarray procedure. Virchows Arch. 2006, 449, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, I.; Korhonen, M.; Petäjäniemi, N.; Karhunen, T.; Thornell, L.E.; Sorokin, L.M.; Konttinen, Y.T. Laminin isoforms in fetal and adult human adrenal cortex. J. Clin. Endocrinol. Metab. 2003, 88, 4960–4966. [Google Scholar] [CrossRef] [Green Version]
- Volante, M.; Bollito, E.; Sperone, P.; Tavaglione, V.; Daffara, F.; Porpiglia, F.; Terzolo, M.; Berruti, A.; Papotti, M. Clinicopathological study of a series of 92 adrenocortical carcinomas: From a proposal of simplified diagnostic algorithm to prognostic stratification. Histopathology 2009, 55, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Senger, D.R.; Perruzzi, C.A.; Gracey, C.F.; Papadopoulos, A.; Tenen, D.G. Secreted phosphoproteins associated with neoplastic transformation: Close homology with plasma proteins cleaved during blood coagulation. Cancer Res. 1988, 48, 5770–5774. [Google Scholar]
- Shijubo, N.; Uede, T.; Kon, S.; Nagata, M.; Abe, S. Vascular endothelial growth factor and osteopontin in tumor biology. Crit. Rev. Oncog. 2000, 11, 135–146. [Google Scholar]
- Rodrigues, L.R.; Teixeira, J.A.; Schmitt, F.L.; Paulsson, M.; Lindmark-Mänsson, H. The role of osteopontin in tumor progression and metastasis in breast cancer. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Takahashi, K.; Takahashi, F.; Shimizu, K.; Ohshita, F.; Kameda, Y.; Maeda, K.; Nishio, K.; Fukuchi, Y. Differential osteopontin expression in lung cancer. Cancer Lett. 2001, 171, 215–222. [Google Scholar] [CrossRef]
- Kim, J.-H.; Skates, S.J.; Uede, T.; Wong, K.; Schorge, J.O.; Feltmate, C.M.; Berkowitz, R.S.; Cramer, D.W.; Mok, S.C. Osteopontin as a potential diagnostic biomarker for ovarian cancer. JAMA 2002, 287, 1671–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ue, T.; Yokozaki, H.; Kitadai, Y.; Yamamoto, S.; Yasui, W.; Ishikawa, T.; Tahara, E. Co-expression of osteopontin and CD44v9 in gastric cancer. Int. J. Cancer 1998, 79, 127–132. [Google Scholar] [CrossRef]
- Gotoh, M.; Sakamoto, M.; Kanetaka, K.; Chuuma, M.; Hirohashi, S. Overexpression of osteopontin in hepatocellular carcinoma. Pathol. Int. 2002, 52, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Forootan, S.S.; Foster, C.S.; Aachi, V.R.; Adamson, J.; Smith, P.H.; Lin, K.; Ke, Y. Prognostic significance of osteopontin expression in human prostate cancer. Int. J. Cancer 2006, 118, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Cappia, S.; Righi, L.; Mirabelli, D.; Ceppi, P.; Bacillo, E.; Ardissone, F.; Molinaro, L.; Scagliotti, G.V.; Papotti, M. Prognostic role of osteopontin expression in malignant pleural mesothelioma. Am. J. Clin. Pathol. 2008, 130, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overgaard, J.; Eriksen, J.G.; Nordsmark, M.; Alsner, J.; Horsman, M.R. Plasma osteopontin, hypoxia, and response to the hypoxia sensitiser nimorazole in radiotherapy of head and neck cancer: Results from the DAHANCA 5 randomised double-blind placebo-controlled trial. Lancet Oncol. 2005, 6, 757–764. [Google Scholar] [CrossRef]
- Vergis, R.; Corbishley, C.M.; Norman, A.R.; Bartlett, J.; Jhavar, S.; Borre, M.; Heeboll, S.; Horwich, A.; Huddart, R.; Khoo, V.; et al. Intrinsic markers of tumour hypoxia and angiogenesis in localised prostate cancer and outcome of radical treatment: A retrospective analysis of two randomised radiotherapy trials and one surgical cohort study. Lancet Oncol. 2008, 9, 342–351. [Google Scholar] [CrossRef]
- Kobayashi, T.; Chanmee, T.; Itano, N. Hyaluronan: Metabolism and function. Biomolecules 2020, 10, 1525. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Q.; Zhang, F.; Luo, W. Effect of ligand-receptor interaction of osteopontin-CD44 on the expression of hyaluronic acid in human knee osteoarthritic chondrocytes in vitro. Zhong Nan Da Xue Xue Bao. Yi Xue Ban J. Cent. South Univ. Med. Sci. 2020, 45, 1015–1023. [Google Scholar] [CrossRef]
- Tiainen, S.; Oikari, S.; Tammi, M.; Rilla, K.; Hämäläinen, K.; Tammi, R.; Kosma, V.-M.; Auvinen, P. High extent of O-GlcNAcylation in breast cancer cells correlates with the levels of HAS enzymes, accumulation of hyaluronan, and poor outcome. Breast Cancer Res. Treat. 2016, 160, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Sapudom, J.; Müller, C.D.; Nguyen, K.-T.; Martin, S.; Anderegg, U.; Pompe, T. Matrix remodeling and hyaluronan production by myofibroblasts and cancer-associated fibroblasts in 3D collagen matrices. Gels 2020, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Adamia, S.; Kriangkum, J.; Belch, A.R.; Pilarski, L.M. Aberrant posttranscriptional processing of hyaluronan synthase 1 in malignant transformation and tumor progression. Adv. Cancer Res. 2014, 123, 67–94. [Google Scholar] [CrossRef] [PubMed]
- De Sá, V.K.; Rocha, T.P.; Moreira, A.; Soares, F.A.; Takagaki, T.; Carvalho, L.; Nicholson, A.G.; Capelozzi, V.L. Hyaluronidases and hyaluronan synthases expression is inversely correlated with malignancy in lung/bronchial pre-neoplastic and neoplastic lesions, affecting prognosis. Braz. J. Med. Biol. Res. 2015, 48, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Poukka, M.; Bykachev, A.; Siiskonen, H.; Tyynelä-Korhonen, K.; Auvinen, P.; Pasonen-Seppänen, S.; Sironen, R. Decreased expression of hyaluronan synthase 1 and 2 associates with poor prognosis in cutaneous melanoma. BMC Cancer 2016, 16, 313. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Description | t-Test (p-Value) | Fold Up/Down Regulation |
|---|---|---|---|
| BFGF/FGFB | Fibroblast growth factor 2 | 0.0484 | −5.66 |
| DHAND2/Hed | Heart and neural crest derivatives expressed 2 | 0.0234 | −13.61 |
| HAS1 | Hyaluronan synthase 1 | 0.0287 | 4.46 |
| F-TCF/HGFB | Hepatocyte growth factor | 0.0489 | −9.02 |
| IL-1/IL1-BETA | Interleukin 1, β | 0.0448 | −8.00 |
| LAMA | Laminin, α2 | 0.0391 | −16.00 |
| E170/LAMNA | Laminin, α3 | 0.0409 | 4.23 |
| KIAA1907 | Laminin, α5 | 0.0280 | −3.86 |
| COX1/COX3 | Prostaglandin-endoperoxide synthase 1 (cyclooxygenase) | 0.0442 | −5.20 |
| BNSP/BSPI | osteopontin, bone sialoprotein I | 0.0450 | 20.32 |
| CLEVER1/FEEL1 | Stabilin 1 | 0.0215 | −7.45 |
| CED/DPD1 | Transforming growth factor, β 1 | 0.0365 | −5.53 |
| THBS/TSP | Thrombospondin 1 | 0.0456 | −10.82 |
| DIF/TNF-alpha | Tumor necrosis factor (TNF superfamily, member 2) | 0.0270 | −11.66 |
| IHC Semiquantitative Score | * ACC (#73) | § ACA (#20) | p Value | |
|---|---|---|---|---|
| Laminin α3 (vascular network) | 0 | 13 (17.8%) | 3 (15%) | 0.04 |
| 1+ | 20 (27.4%) | 11 (55%) | ||
| 2+ | 23 (31.5%) | 6 (30%) | ||
| 3+ | 17 (23.3%) | 0 | ||
| HAS-1 (tumor cells) | 0 | 42 (57.5%) | 13 (65%) | 0.55 |
| 1+ | 17 (23.3%) | 5 (25%) | ||
| 2+ | 7 (9.6%) | 2 (10%) | ||
| 3+ | 7 (9.6%) | 0 | ||
| HAS-1 (vascular network) | 0 | 12 (16.4%) | 2 (10%) | 0.51 |
| 1+ | 21 (28.8%) | 9 (45%) | ||
| 2+ | 18 (24.7%) | 5 (25%) | ||
| 3+ | 22 (30.1%) | 4 (20%) | ||
| Osteopontin (tumor cells) | negative | 48 (65.8%) | 20 (100%) | 0.0022 |
| positive | 25 (34.2%) | 0 |
| Parameter | HAS-1 Positive | HAS-1 Negative | p Value | |
|---|---|---|---|---|
| Sex | M | 8 | 6 | 0.02 |
| F | 7 | 10 | ||
| Age | <46 * | 3 | 10 | 0.02 |
| ≥46 | 12 | 6 | ||
| Functional status | Nonfunctioning | 8 | 10 | 0.62 |
| Functioning † | 7 | 6 | ||
| Tumor weight (g) | <275 * | 10 | 9 | 0.55 |
| ≥275 | 5 | 7 | ||
| Tumor size (cm) | <11 * | 10 | 6 | 0.10 |
| ≥11 | 5 | 10 | ||
| Mitoses | <8 * | 10 | 4 | 0.02 |
| ≥8 | 5 | 12 | ||
| Atypical mitoses | absent | 11 | 6 | 0.045 |
| present | 4 | 10 | ||
| Necrosis | absent | 4 | 1 | 0.12 |
| present | 11 | 15 | ||
| Venous invasion | absent | 2 | 6 | 0.12 |
| present | 13 | 10 | ||
| Sinusoid invasion | absent | 8 | 9 | 0.87 |
| present | 7 | 7 | ||
| Capsular invasion | absent | 8 | 8 | 0.85 |
| present | 7 | 8 | ||
| Nuclear atypia | absent | 3 | 4 | 0.74 |
| present | 12 | 12 | ||
| Weiss score | 3–6 | 8 | 7 | 0.59 |
| 7–9 | 7 | 9 | ||
| ENSAT stage | I | 4 | 0 | 0.068 |
| II | 10 | 13 | ||
| III | 1 | 3 | ||
| Mitotane treatment (4 cases missing) | no | 3 | 1 | 0.60 |
| yes | 12 | 11 | ||
| Status | NED | 10 | 3 | 0.0069 |
| AWD/DOD | 5 | 13 | ||
| Median DFS | months | not reached | 49 | 0.040 |
| Median OS | months | not reached | 17 | 0.197 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volante, M.; Rapa, I.; Metovic, J.; Napoli, F.; Tampieri, C.; Duregon, E.; Terzolo, M.; Papotti, M. Differential Expression Profiles of Cell-to-Matrix-Related Molecules in Adrenal Cortical Tumors: Diagnostic and Prognostic Implications. J. Pers. Med. 2021, 11, 378. https://doi.org/10.3390/jpm11050378
Volante M, Rapa I, Metovic J, Napoli F, Tampieri C, Duregon E, Terzolo M, Papotti M. Differential Expression Profiles of Cell-to-Matrix-Related Molecules in Adrenal Cortical Tumors: Diagnostic and Prognostic Implications. Journal of Personalized Medicine. 2021; 11(5):378. https://doi.org/10.3390/jpm11050378
Chicago/Turabian StyleVolante, Marco, Ida Rapa, Jasna Metovic, Francesca Napoli, Cristian Tampieri, Eleonora Duregon, Massimo Terzolo, and Mauro Papotti. 2021. "Differential Expression Profiles of Cell-to-Matrix-Related Molecules in Adrenal Cortical Tumors: Diagnostic and Prognostic Implications" Journal of Personalized Medicine 11, no. 5: 378. https://doi.org/10.3390/jpm11050378