
Effect of MHC Linked 7-Gene Signature on Delayed Hepatocellular Carcinoma Recurrence

 , ,
, ,
Abstract
:1. Introduction
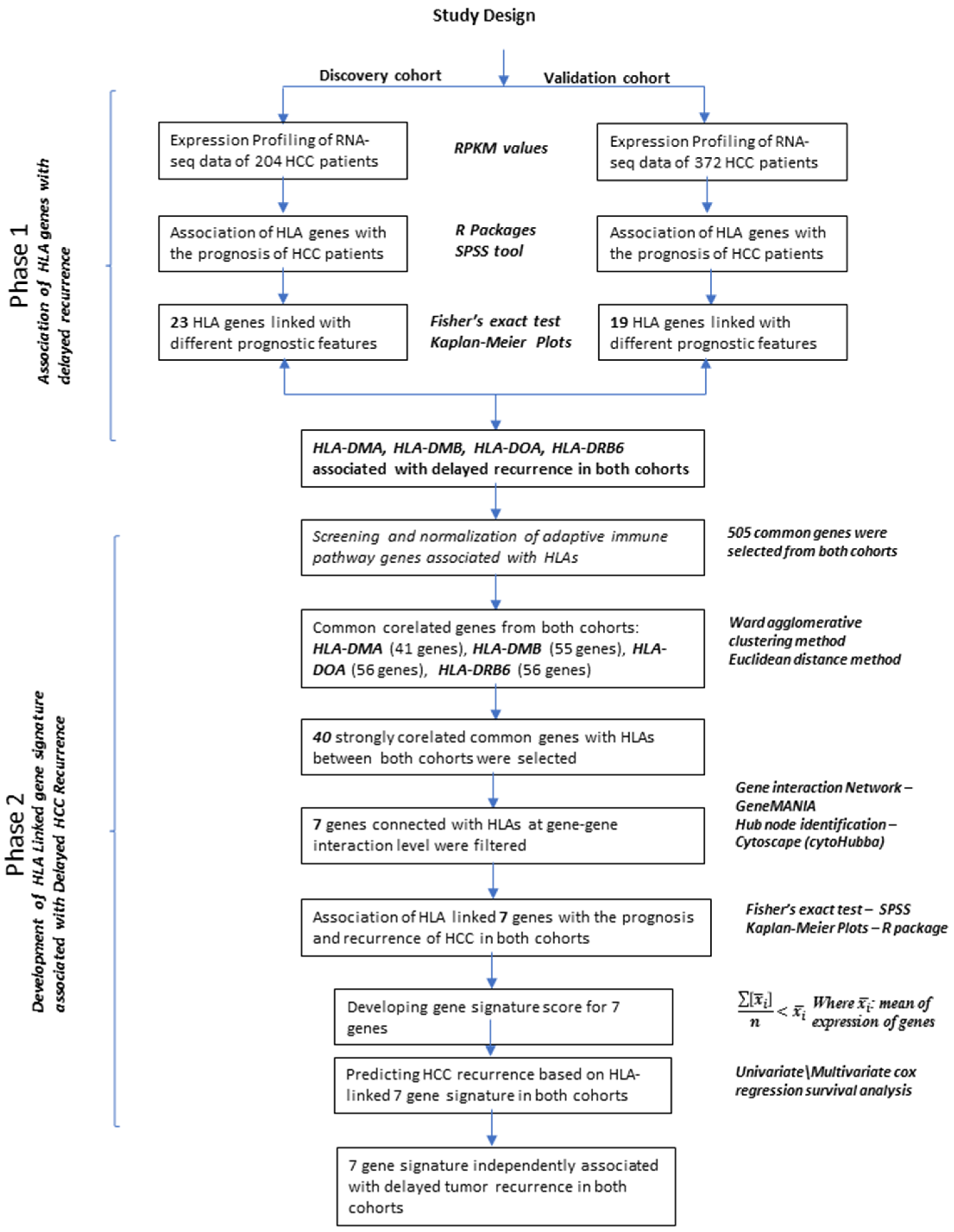
2. Materials and Methods
2.1. RNA Sequencing Data of 204 HCC Patients (Discovery Cohort)
2.2. RNA Sequencing Data of 372 HCC Patients (Validation Cohort)
2.3. Statistical Analysis
2.4. Clustering and Correlation Analysis
2.5. Network Analysis of Candidate Co-Expressed Genes
2.6. Gene Signature Score Calculation
3. Results
3.1. Clinical Analysis of MHC Genes (Discovery Cohort)
3.2. Clinical Analysis of HLA Genes (Validation Cohort)
3.3. Clustering Analysis of HLA Genes with Adaptive Immune and HCC Related Genes
3.4. Network Analysis and Hub Gene Identification of 40 Genes Co-Expressed with HLAs
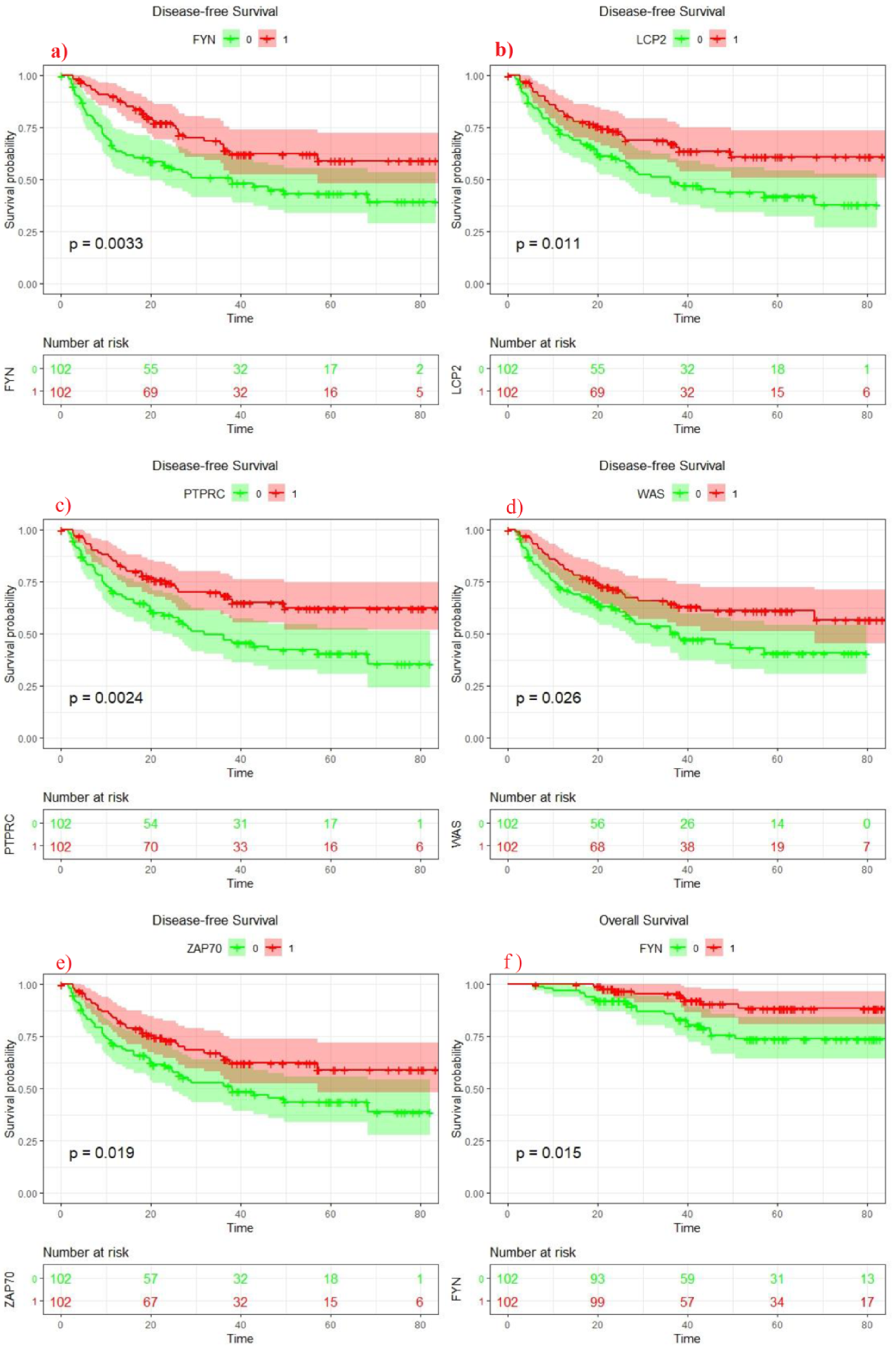
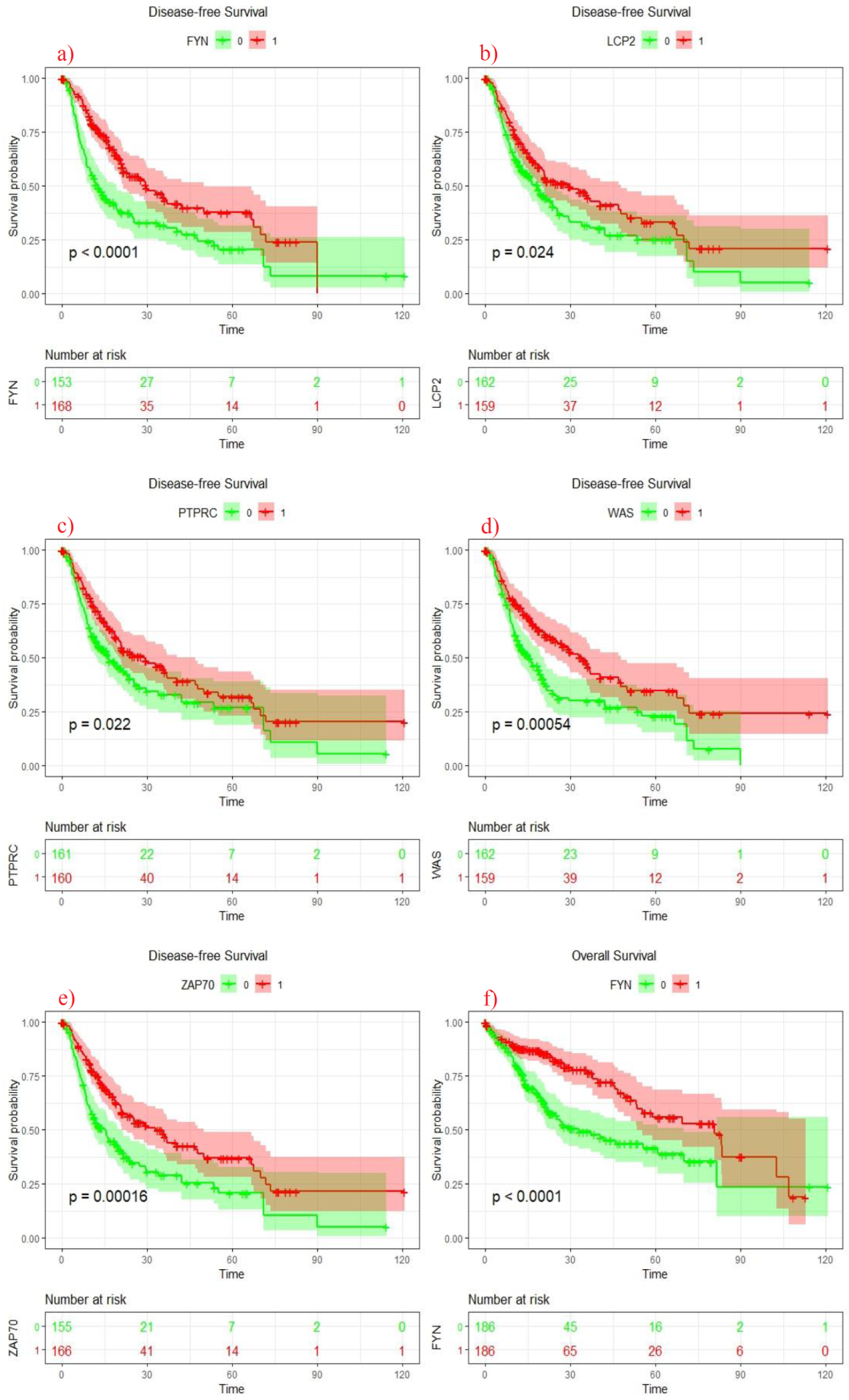
3.5. Clinical Analysis of Seven Genes Signature
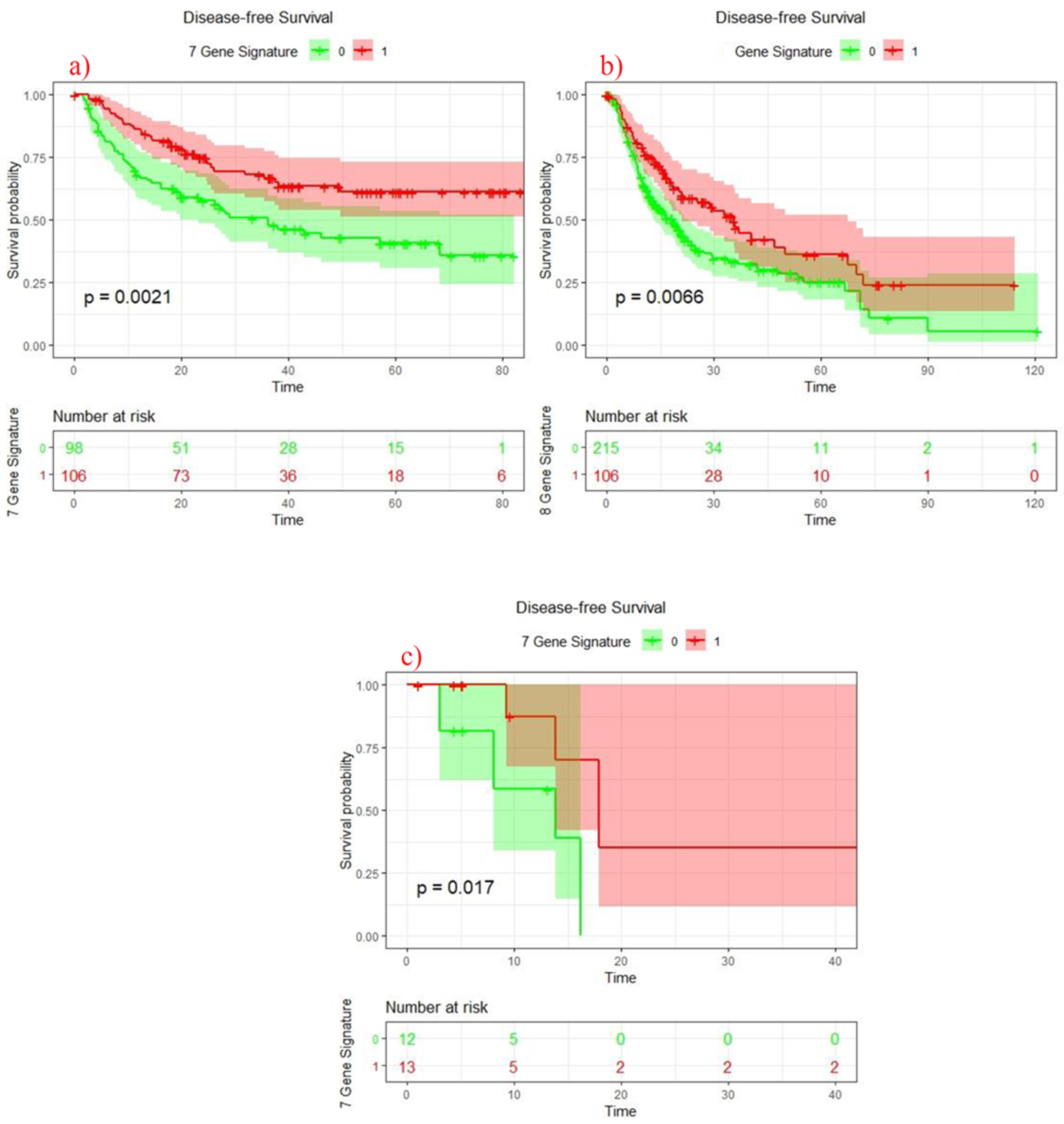
3.6. Survival Analysis of 7 Genes Signature Score
3.7. COX Regression Survival Analysis in Discovery and Validation Cohorts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, C.-T.; Lee, H.-L.; Chiou, H.-L.; Chou, C.-H.; Wang, P.-H.; Yang, S.-F.; Chou, Y.-E. Impacts of WNT1-Inducible Signaling Pathway Protein 1 Polymorphism on Hepatocellular Carcinoma Development. PLoS ONE 2018, 13, e0198967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, F.X.; Ribes, J.; Cléries, R.; Díaz, M. Epidemiology of Hepatocellular Carcinoma. Clin. Liver Dis. 2005, 9, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Charles, A.; Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M.J. The Major Histocompatibility Complex and Its Functions. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Choo, S.Y. The HLA System: Genetics, Immunology, Clinical Testing, and Clinical Implications. Yonsei Med. J. 2007, 48, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swets, M.; König, M.H.; Zaalberg, A.; Dekker-Ensink, N.G.; Gelderblom, H.; van de Velde, C.J.H.; van den Elsen, P.J.; Kuppen, P.J.K. HLA-G and Classical HLA Class I Expression in Primary Colorectal Cancer and Associated Liver Metastases. Hum. Immunol. 2016, 77, 773–779. [Google Scholar] [CrossRef]
- Krijgsman, D.; Roelands, J.; Hendrickx, W.; Bedognetti, D.; Kuppen, P.J.K. HLA-G: A New Immune Checkpoint in Cancer? Int. J. Mol. Sci. 2020, 21, 4528. [Google Scholar] [CrossRef]
- Sliker, B.H.; Goetz, B.T.; Barnes, R.; King, H.; Maurer, H.C.; Olive, K.P.; Solheim, J.C. HLA-B Influences Integrin Beta-1 Expression and Pancreatic Cancer Cell Migration. Exp. Cell Res. 2020, 390, 111960. [Google Scholar] [CrossRef]
- Ben Yahia, H.; Boujelbene, N.; Babay, W.; Ben Safta, I.; Dhouioui, S.; Zemni, I.; Ali Ayadi, M.; Charfi, L.; Ouzari, H.I.; Rebmann, V.; et al. Expression Analysis of Immune-Regulatory Molecules HLA-G, HLA-E and IDO in Endometrial Cancer. Hum. Immunol. 2020, 81, 305–313. [Google Scholar] [CrossRef]
- Najafimehr, H.; Hajizadeh, N.; Nazemalhosseini-Mojarad, E.; Pourhoseingholi, M.A.; Abdollahpour-Alitappeh, M.; Ashtari, S.; Zali, M.R. The Role of Human Leukocyte Antigen Class I on Patient Survival in Gastrointestinal Cancers: A Systematic Review and Meta- Analysis. Sci. Rep. 2020, 10, 728. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, K.; Ishigami, S.; Kijima, Y.; Funasako, Y.; Hirata, M.; Okumura, H.; Shinchi, H.; Koriyama, C.; Ueno, S.; Yoshinaka, H.; et al. Clinical Implication of HLA Class I Expression in Breast Cancer. BMC Cancer 2011, 11, 454. [Google Scholar] [CrossRef]
- He, Y.; Rozeboom, L.; Rivard, C.J.; Ellison, K.; Dziadziuszko, R.; Yu, H.; Zhou, C.; Hirsch, F.R. MHC Class II Expression in Lung Cancer. Lung Cancer 2017, 112, 75–80. [Google Scholar] [CrossRef]
- Park, I.A.; Hwang, S.-H.; Song, I.H.; Heo, S.-H.; Kim, Y.-A.; Bang, W.S.; Park, H.S.; Lee, M.; Gong, G.; Lee, H.J. Expression of the MHC Class II in Triple-Negative Breast Cancer Is Associated with Tumor-Infiltrating Lymphocytes and Interferon Signaling. PLoS ONE 2017, 12, e0182786. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xiang, Y.; Sheng, J.; Zhang, D.; Yao, X.; Yang, Y.; Zhang, X. Immunotherapy for Hepatocellular Carcinoma: Current Advances and Future Expectations. J. Immunol. Res. 2018, 2018. [Google Scholar] [CrossRef]
- Lee, M.-H.; Huang, Y.-H.; Chen, H.-Y.; Khor, S.-S.; Chang, Y.-H.; Lin, Y.-J.; Jen, C.-L.; Lu, S.-N.; Yang, H.-I.; Nishida, N.; et al. Human Leukocyte Antigen Variants and Risk of Hepatocellular Carcinoma Modified by Hepatitis C Virus Genotypes: A Genome-Wide Association Study. Hepatology 2018, 67, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qian, J.; Yang, Y.; Zhao, W.; Dai, J.; Bei, J.-X.; Foo, J.N.; McLaren, P.J.; Li, Z.; Yang, J.; et al. GWAS Identifies Novel Susceptibility Loci on 6p21.32 and 21q21.3 for Hepatocellular Carcinoma in Chronic Hepatitis B Virus Carriers. PLoS Genet 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Sawai, H.; Nishida, N.; Khor, S.-S.; Honda, M.; Sugiyama, M.; Baba, N.; Yamada, K.; Sawada, N.; Tsugane, S.; Koike, K.; et al. Genome-Wide Association Study Identified New Susceptible Genetic Variants in HLA Class I Region for Hepatitis B Virus-Related Hepatocellular Carcinoma. Sci. Rep. 2018, 8, 7958. [Google Scholar] [CrossRef]
- Wagenaar, T.R.; Zabludoff, S.; Ahn, S.-M.; Allerson, C.; Arlt, H.; Baffa, R.; Cao, H.; Davis, S.; Garcia-Echeverria, C.; Gaur, R.; et al. Anti–MiR-21 Suppresses Hepatocellular Carcinoma Growth via Broad Transcriptional Network Deregulation. Mol. Cancer Res. 2015, 13, 1009–1021. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.-M.; Jang, S.J.; Shim, J.H.; Kim, D.; Hong, S.-M.; Sung, C.O.; Baek, D.; Haq, F.; Ansari, A.A.; Lee, S.Y.; et al. Genomic Portrait of Resectable Hepatocellular Carcinomas: Implications of RB1 and FGF19 Aberrations for Patient Stratification. Hepatology 2014, 60, 1972–1982. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Deng, M.; Brägelmann, J.; Schultze, J.L.; Perner, S. Web-TCGA: An Online Platform for Integrated Analysis of Molecular Cancer Data Sets. BMC Bioinform. 2016, 17, 72. [Google Scholar] [CrossRef] [Green Version]
- Ally, A.; Balasundaram, M.; Carlsen, R.; Chuah, E.; Clarke, A.; Dhalla, N.; Holt, R.A.; Jones, S.J.M.; Lee, D.; Ma, Y.; et al. Comprehensive and Integrative Genomic Characterization of Hepatocellular Carcinoma. Cell 2017, 169, 1327–1341.e23. [Google Scholar] [CrossRef] [Green Version]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Clustering Method: Clustering Criterion and Agglomerative Algorithm. J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. CytoHubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishayee, A. The Role of Inflammation and Liver Cancer. Adv. Exp. Med. Biol. 2014, 816, 401–435. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, S.; Zeng, S.; Shen, H. From Bench to Bed: The Tumor Immune Microenvironment and Current Immunotherapeutic Strategies for Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 396. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-K.; Liao, X.-W.; Yang, C.-K.; Yu, T.-D.; Liu, Z.-Q.; Gong, Y.-Z.; Huang, K.-T.; Zeng, X.-M.; Han, C.-Y.; Zhu, G.-Z.; et al. Diagnostic and Prognostic Biomarkers of Human Leukocyte Antigen Complex for Hepatitis B Virus-Related Hepatocellular Carcinoma. J. Cancer 2019, 10, 5173–5190. [Google Scholar] [CrossRef] [PubMed]
- Callahan, M.J.; Nagymanyoki, Z.; Bonome, T.; Johnson, M.E.; Litkouhi, B.; Sullivan, E.H.; Hirsch, M.S.; Matulonis, U.A.; Liu, J.; Birrer, M.J.; et al. Increased HLA-DMB Expression in the Tumor Epithelium Is Associated with Increased CTL Infiltration and Improved Prognosis in Advanced-Stage Serous Ovarian Cancer. Clin. Cancer Res. 2008, 14, 7667–7673. [Google Scholar] [CrossRef] [Green Version]
- Roychowdhury, A.; Samadder, S.; Das, P.; Mazumder, D.I.; Chatterjee, A.; Addya, S.; Mondal, R.; Roy, A.; Roychoudhury, S.; Panda, C.K. Deregulation of H19 Is Associated with Cervical Carcinoma. Genomics 2020, 112, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Liu, L.; Li, W. A Prognostic Risk Model Based on Immune-related Genes Predicts Overall Survival of Patients with Hepatocellular Carcinoma. Health Sci. Rep. 2020, 3. [Google Scholar] [CrossRef]
- Malmberg, K.-J.; Levitsky, V.; Norell, H.; de Matos, C.T.; Carlsten, M.; Schedvins, K.; Rabbani, H.; Moretta, A.; Söderström, K.; Levitskaya, J.; et al. IFN-γ Protects Short-Term Ovarian Carcinoma Cell Lines from CTL Lysis via a CD94/NKG2A-Dependent Mechanism. J. Clin. Invest. 2002, 110, 1515–1523. [Google Scholar] [CrossRef]
- Koh, S.; Shimasaki, N.; Suwanarusk, R.; Ho, Z.Z.; Chia, A.; Banu, N.; Wu Howland, S.; Ong, A.S.M.; Gehring, A.J.; Stauss, H.; et al. A Practical Approach to Immunotherapy of Hepatocellular Carcinoma Using T Cells Redirected Against Hepatitis B Virus. Mol. Ther. Nucleic Acids 2013, 2, e114. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Wu, L.; Zhou, F.; Hong, Z.; Liu, Z.; Yuan, Y. T Cell–Associated Immunotherapy for Hepatocellular Carcinoma. CPB 2017, 41, 609–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Lan, X.; Deng, C.; Zhou, J.; Du, J.; Huang, S.; Li, Y. Molecular Mechanisms Underlying the Evolution of the Slp76 Signalosome. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtney, A.H.; Lo, W.-L.; Weiss, A. TCR Signaling: Mechanisms of Initiation and Propagation. Trends Biochem. Sci. 2018, 43, 108–123. [Google Scholar] [CrossRef]
- Shui, J.-W.; Boomer, J.S.; Han, J.; Xu, J.; Dement, G.A.; Zhou, G.; Tan, T.-H. Hematopoietic Progenitor Kinase 1 Negatively Regulates T Cell Receptor Signaling and T Cell-Mediated Immune Responses. Nat. Immunol. 2007, 8, 84–91. [Google Scholar] [CrossRef] [PubMed]
- zur Hausen, J.D.; Burn, P.; Amrein, K.E. Co-Localization of Fyn with CD3 Complex, CD45 or CD28 Depends on Different Mechanisms. Eur. J. Immunol. 1997, 27, 2643–2649. [Google Scholar] [CrossRef]
- Shiroo, M.; Goff, L.; Biffen, M.; Shivnan, E.; Alexander, D. CD45 Tyrosine Phosphatase-Activated P59fyn Couples the T Cell Antigen Receptor to Pathways of Diacylglycerol Production, Protein Kinase C Activation and Calcium Influx. EMBO J. 1992, 11, 4887–4897. [Google Scholar] [CrossRef] [PubMed]
- Mason, L.H.; Willette-Brown, J.; Taylor, L.S.; McVicar, D.W. Regulation of Ly49D/DAP12 Signal Transduction by Src-Family Kinases and CD45. J. Immunol. 2006, 176, 6615–6623. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.D.; Ruysscher, D.D.; Agostinis, P. Immunological Metagene Signatures Derived from Immunogenic Cancer Cell Death Associate with Improved Survival of Patients with Lung, Breast or Ovarian Malignancies: A Large-Scale Meta-Analysis. OncoImmunology 2016, 5, e1069938. [Google Scholar] [CrossRef] [Green Version]
- Landskron, J.; Kraggerud, S.M.; Wik, E.; Dørum, A.; Bjørnslett, M.; Melum, E.; Helland, Ø.; Bjørge, L.; Lothe, R.A.; Salvesen, H.B.; et al. C77G in PTPRC (CD45) Is No Risk Allele for Ovarian Cancer, but Associated with Less Aggressive Disease. PLoS ONE 2017, 12, e0182030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frugtniet, B.; Jiang, W.G.; Martin, T.A. Role of the WASP and WAVE Family Proteins in Breast Cancer Invasion and Metastasis. Breast Cancer 2015, 7, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Claus, R.; Lucas, D.M.; Ruppert, A.S.; Williams, K.E.; Weng, D.; Patterson, K.; Zucknick, M.; Oakes, C.C.; Rassenti, L.Z.; Greaves, A.W.; et al. Validation of ZAP-70 Methylation and Its Relative Significance in Predicting Outcome in Chronic Lymphocytic Leukemia. Blood 2014, 124, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Cai, L.; Zhu, J.; Chen, M.; Chen, J.; Li, Z.-H.; Liu, X.-D.; Wang, S.-G.; Bie, P.; Jiang, P.; et al. Fyn Requires HnRNPA2B1 and Sam68 to Synergistically Regulate Apoptosis in Pancreatic Cancer. Carcinogenesis 2011, 32, 1419–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Age (Age < 60) | Tumor Size (Size < 5) | Edmonson Grade (Advance) | Microvascular Invasion (Negative) | AFP Level | Overall Survival (Better) | Disease Free Survival (Delayed) |
|---|---|---|---|---|---|---|---|
| FYN | - | Upregulation | - | Upregulation | Upregulation(High) | Upregulation - | Upregulation |
| LCP2 | - | Upregulation | - | Upregulation | - | - | Upregulation |
| NCF4 | - | Upregulation | Upregulation | - | - | - | - |
| PTPRC | - | Upregulation | - | Upregulation | Upregulation(Low) | - | Upregulation |
| TYROBP | Upregulation | - | Upregulation | - | - | - | - |
| WAS | Upregulation | Upregulation | - | - | - | - | Upregulation |
| ZAP70 | - | Upregulation | - | Upregulation | Upregulation(Low) | - | Upregulation |
| Gene ID | Sex (Male) | Grade (Early) | HCV (Negative) | AFP Level (Low) | Alcohol Consumption (High) | Tumor Stages (Early) | Overall Survival (Better) | Disease Free Survival (Delayed) |
|---|---|---|---|---|---|---|---|---|
| FYN | Upregulation | Upregulation | - | Upregulation | - | - | Upregulation- | Upregulation |
| LCP2 | - | - | - | - | - | - | - | Upregulation |
| NCF4 | - | - | Upregulation | - | Upregulation | - | - | - |
| PTPRC | - | - | - | - | - | - | - | Upregulation |
| TYROBP | - | - | - | - | Upregulation | - | - | - |
| WAS | - | - | Upregulation | - | Upregulation | Upregulation | - | Upregulation |
| ZAP70 | - | - | Upregulation | - | Upregulation | Upregulation | Upregulation | Upregulation |
| Univariate Analysis | Multivariate Analysis | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B | SE | Wald | Sig. | Exp(B) | 95.0% CI for Exp(B) | B | SE | Wald | Sig. | HR | 95.0% CI for Exp(B) | |||
| Lower | Upper | Lower | Upper | |||||||||||
| 7GeneSignature | −0.665 | 0.221 | 9.075 | 0.003 | 0.514 | 0.334 | 0.793 | −0.789 | 0.228 | 11.983 | 0.001 | 0.454 | 0.291 | 0.710 |
| AFP high/low | −0.509 | 0.238 | 4.584 | 0.032 | 0.601 | 0.377 | 0.958 | −0.556 | 0.249 | 4.980 | 0.026 | 0.573 | 0.352 | 0.935 |
| Size | 0.461 | 0.222 | 4.298 | 0.038 | 1.586 | 1.026 | 2.453 | - | - | - | - | - | - | - |
| Tumor Number | 1.440 | 0.375 | 14.710 | 0.000 | 4.221 | 2.022 | 8.810 | 1.774 | 0.414 | 18.403 | 0.000 | 5.895 | 2.621 | 13.259 |
| Univariate Analysis | Multivariate Analysis | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B | SE | Wald | Sig. | Exp(B) | 95.0% CI for Exp(B) | B | SE | Wald | Sig. | HR | 95.0% CI for Exp(B) | |||
| Lower | Upper | Lower | Upper | |||||||||||
| 7GeneSignature | −0.450 | 0.167 | 7.247 | 0.007 | 0.638 | 0.460 | 0.885 | −0.514 | 0.218 | 5.563 | 0.018 | 0.598 | 0.390 | 0.917 |
| HBV | −0.664 | 0.180 | 13.592 | 0.000 | 0.515 | 0.361 | 0.733 | −0.497 | 0.211 | 5.561 | 0.018 | 0.608 | 0.403 | 0.920 |
| Metastasis | 1.581 | 0.592 | 7.134 | 0.008 | 4.862 | 1.523 | 15.516 | - | - | - | - | - | - | - |
| T-stage | 0.834 | 0.163 | 26.038 | 0.000 | 2.302 | 1.671 | 3.170 | 0.628 | 0.207 | 9.181 | 0.002 | 1.874 | 1.248 | 2.814 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tariq, F.; Khan, W.; Ahmad, W.; Riaz, S.K.; Khan, M.; Sherwani, S.; Haque, S.; Malik, M.F.A.; Iftikhar, M.J.; Khan, S.; et al. Effect of MHC Linked 7-Gene Signature on Delayed Hepatocellular Carcinoma Recurrence. J. Pers. Med. 2021, 11, 1129. https://doi.org/10.3390/jpm11111129
Tariq F, Khan W, Ahmad W, Riaz SK, Khan M, Sherwani S, Haque S, Malik MFA, Iftikhar MJ, Khan S, et al. Effect of MHC Linked 7-Gene Signature on Delayed Hepatocellular Carcinoma Recurrence. Journal of Personalized Medicine. 2021; 11(11):1129. https://doi.org/10.3390/jpm11111129
Chicago/Turabian StyleTariq, Fomaz, Walizeb Khan, Washaakh Ahmad, Syeda Kiran Riaz, Mahvish Khan, Subuhi Sherwani, Shafiul Haque, Muhammad Faraz Arshad Malik, Muhammad Jahangir Iftikhar, Saif Khan, and et al. 2021. "Effect of MHC Linked 7-Gene Signature on Delayed Hepatocellular Carcinoma Recurrence" Journal of Personalized Medicine 11, no. 11: 1129. https://doi.org/10.3390/jpm11111129