
Serotonin after β-Adrenoreceptors’ Exposition: New Approaches for Personalized Data in Breast Cancer Cells

 ,
,  , , and
, , and 
Abstract
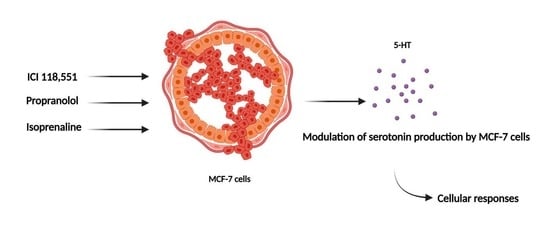
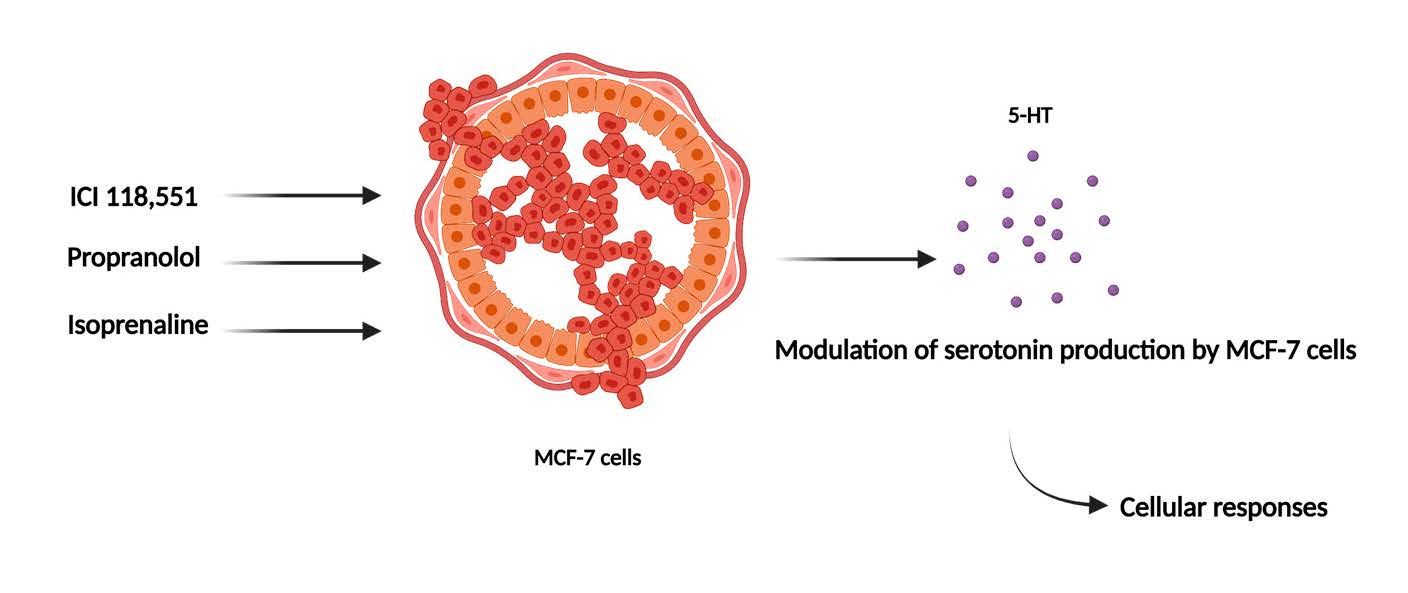
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
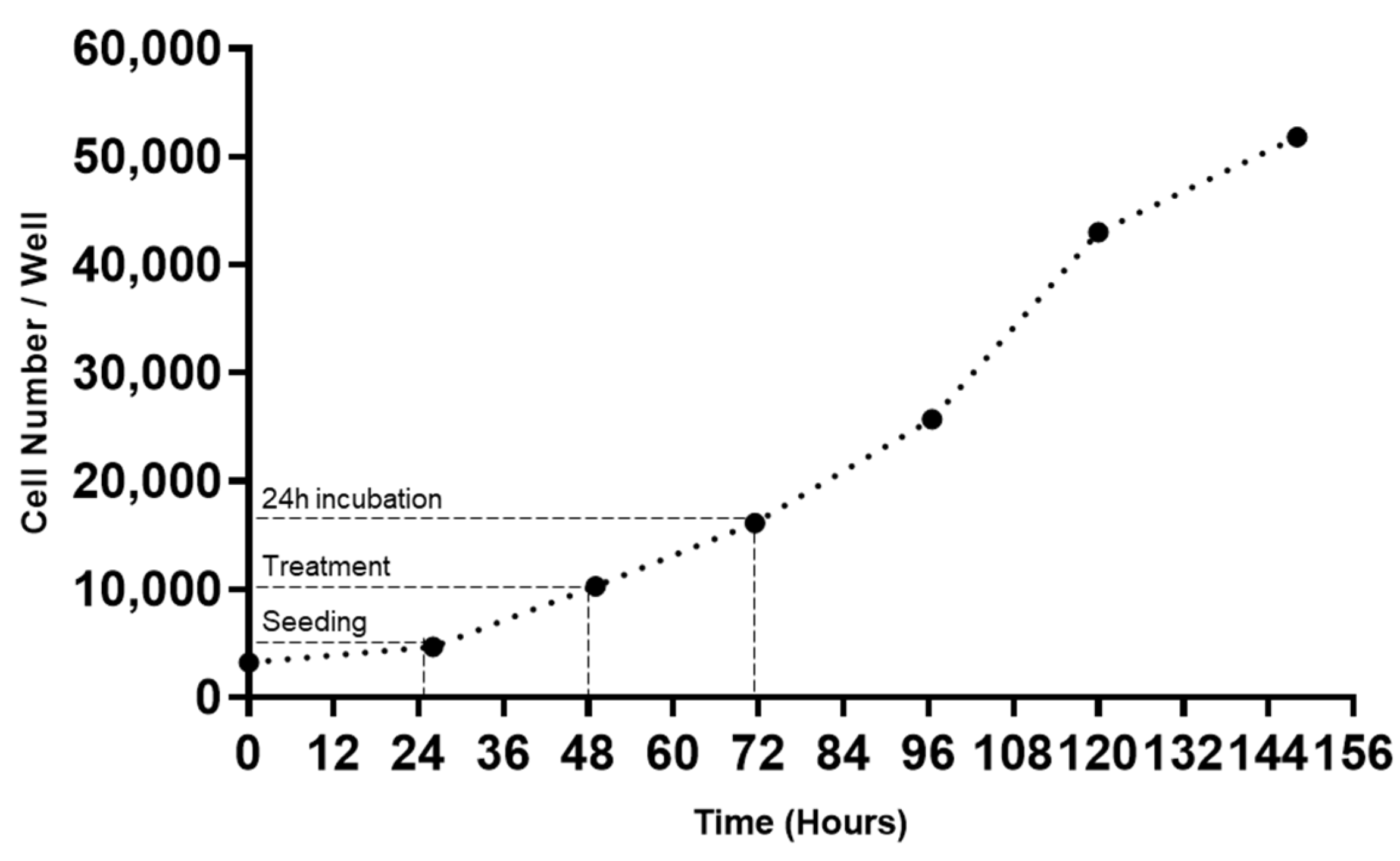
2.3. Cell Growth Curve Assay
2.4. Cell Treatment
2.5. Cell Viability Assays
2.6. Cell Morphology Visualization
2.7. HPLC-ECD
2.8. Statistical and Data Analysis
3. Results
3.1. MCF-7 Growth Curve
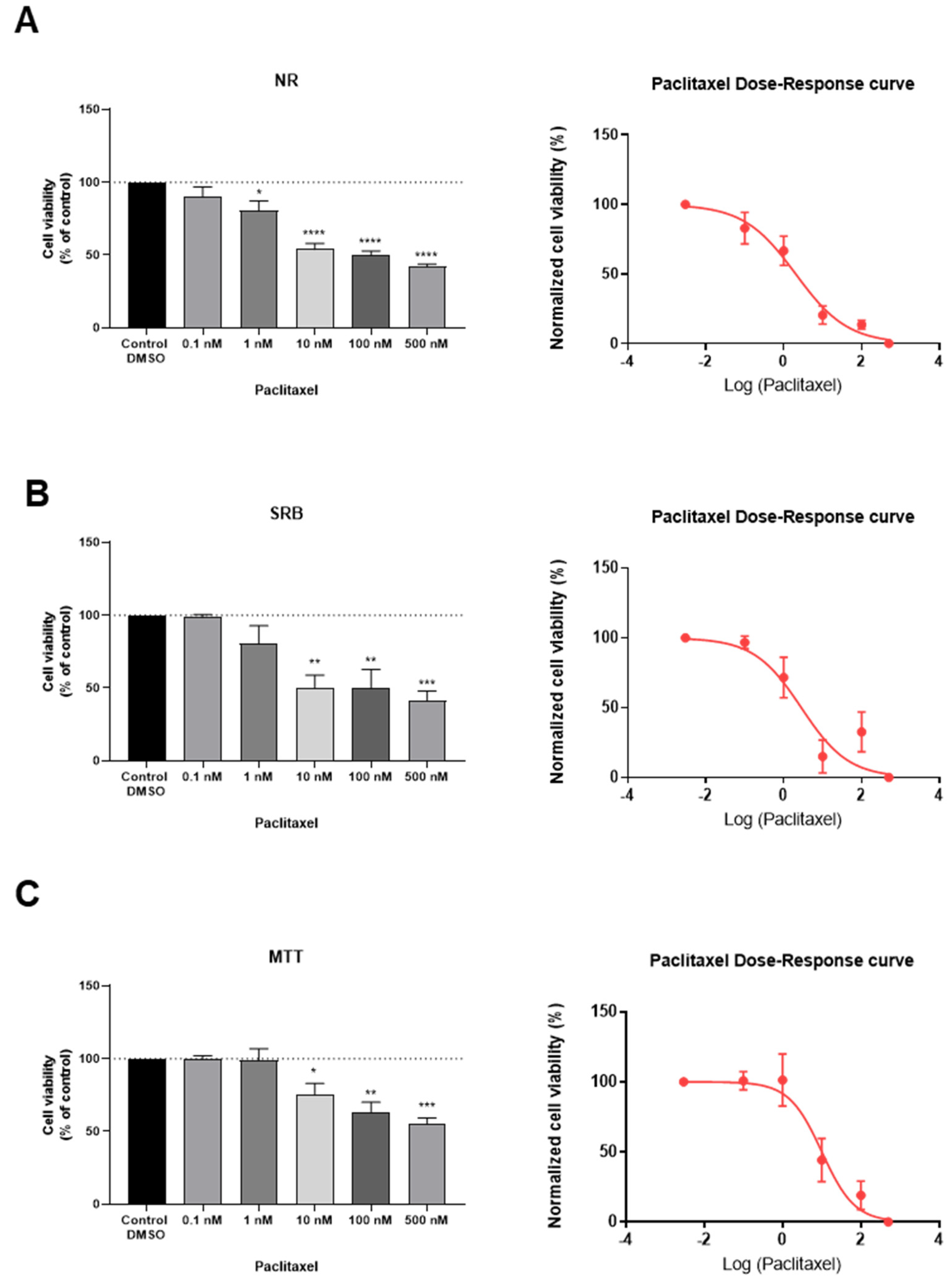
3.2. Effect of Paclitaxel on MCF-7 Cellular Viability
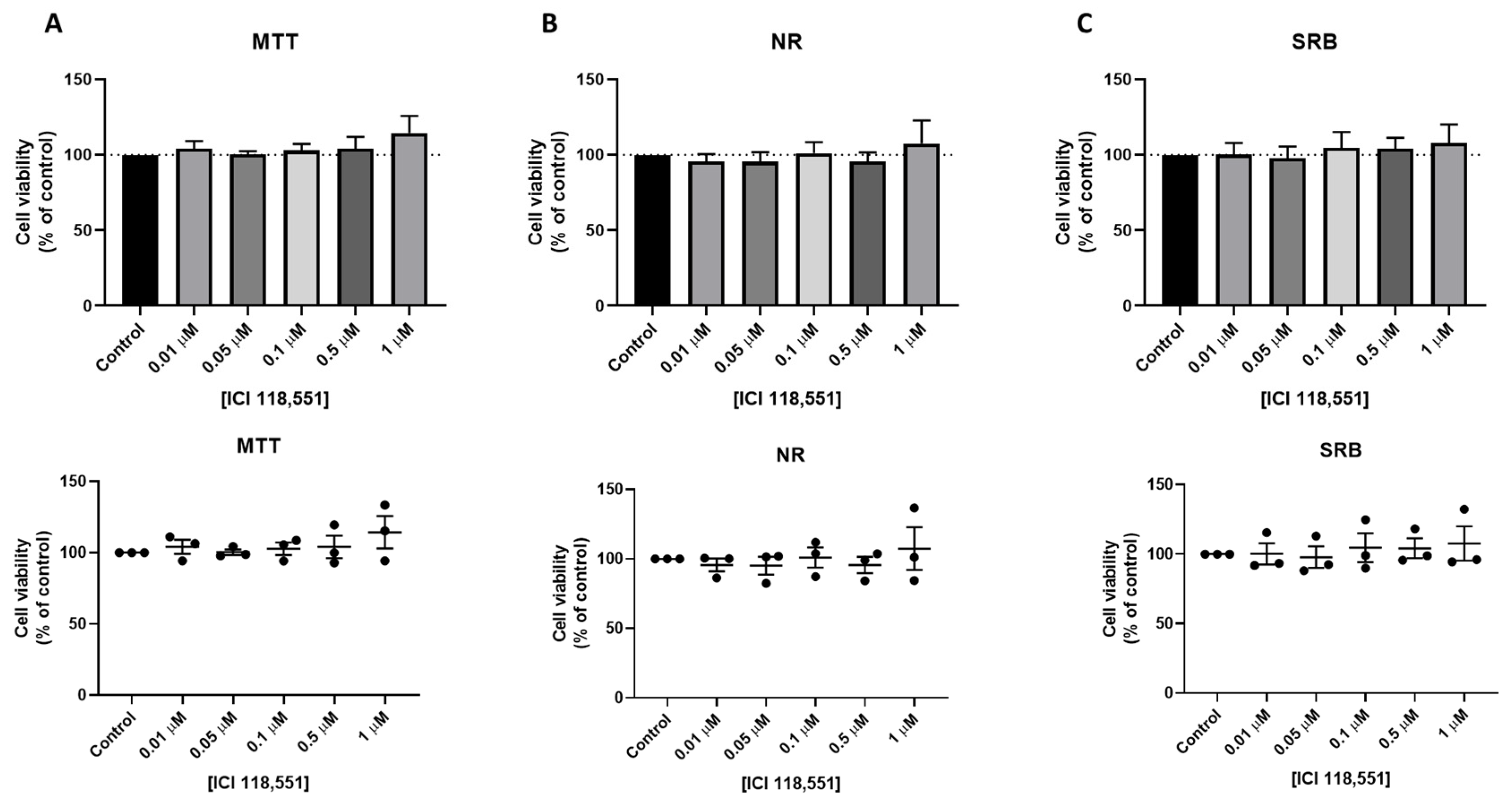
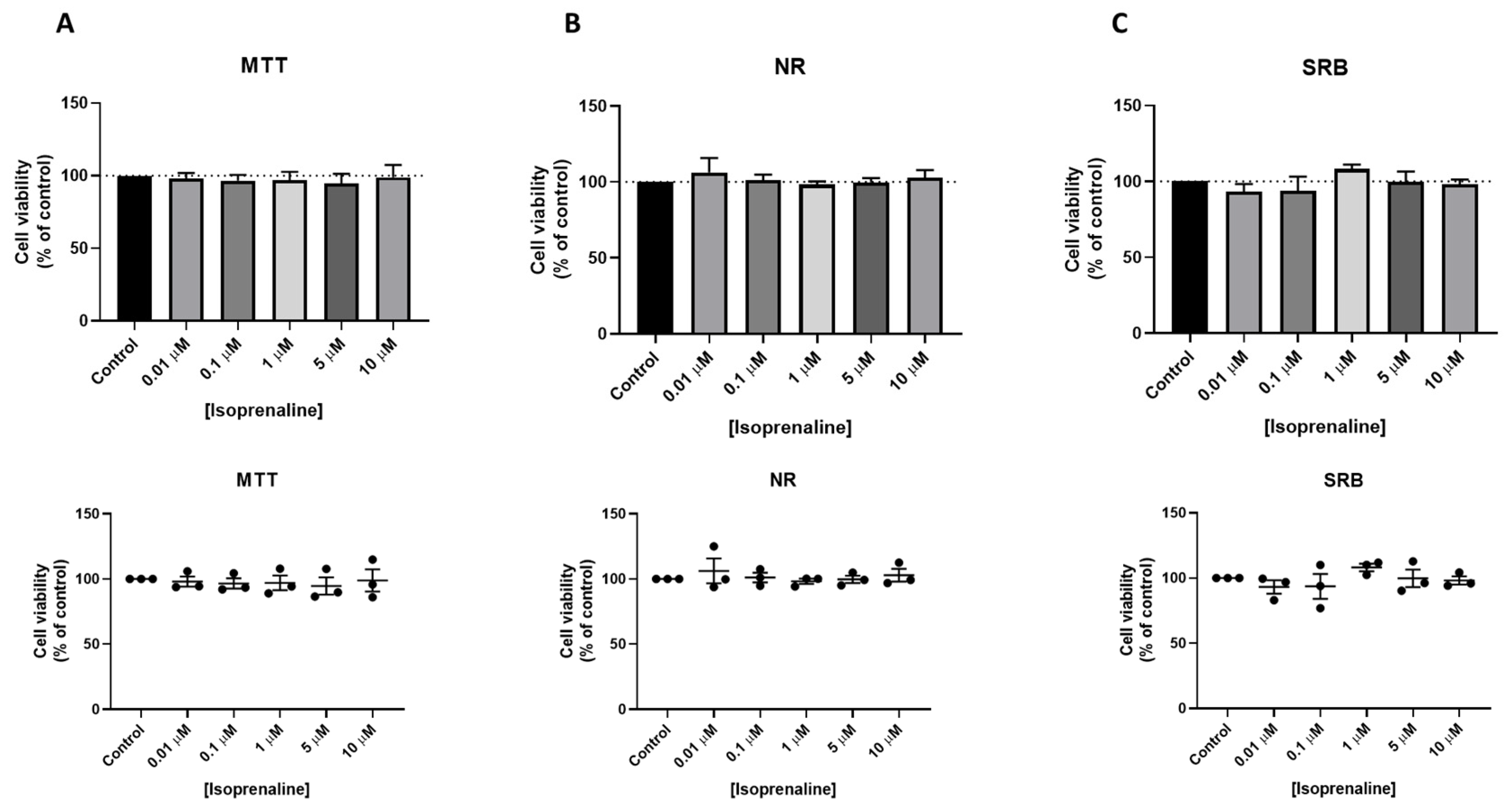
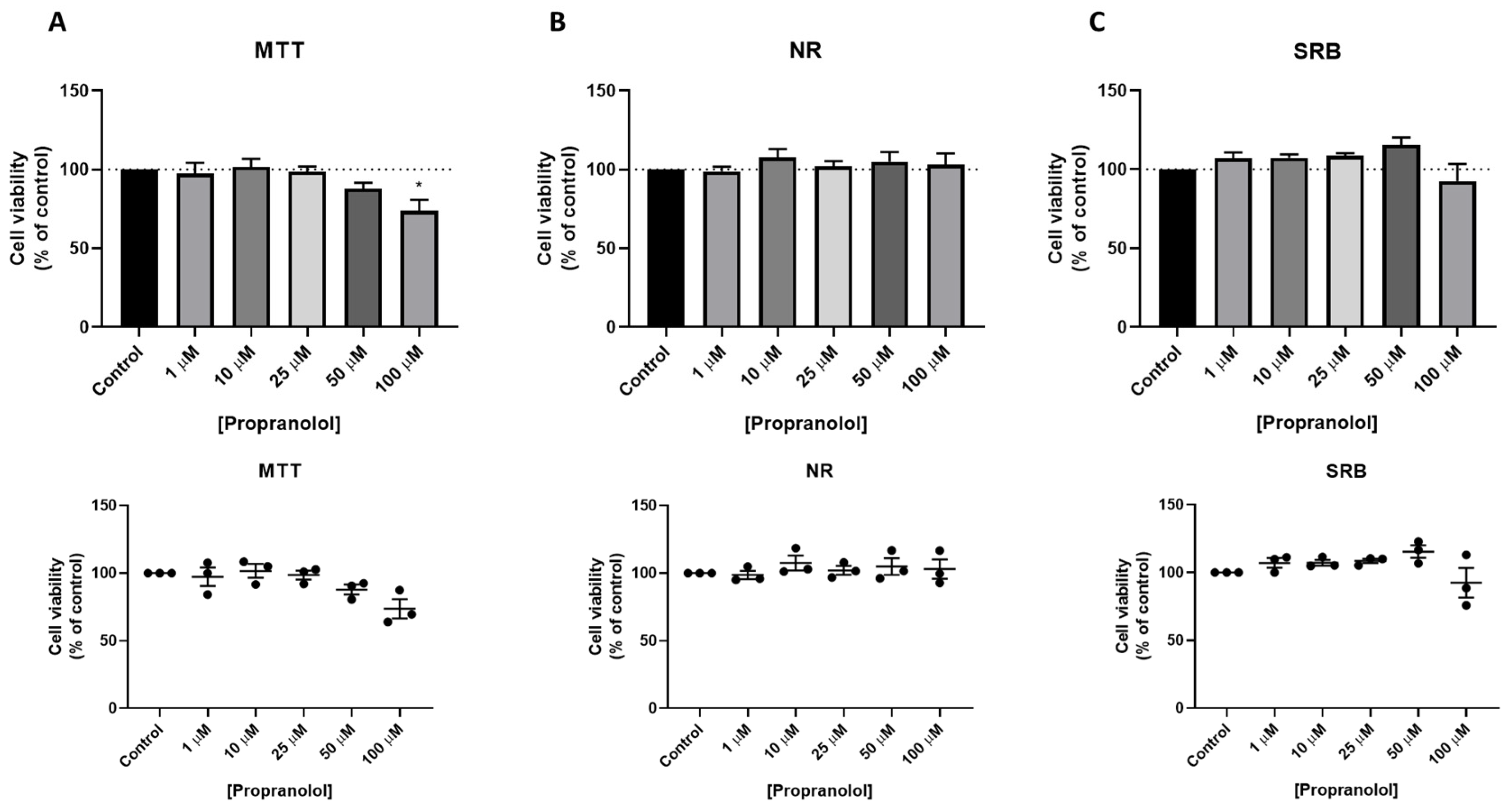
3.3. Effect of Isoprenaline, Propranolol, and ICI 118,551 on MCF-7 Cellular Viability
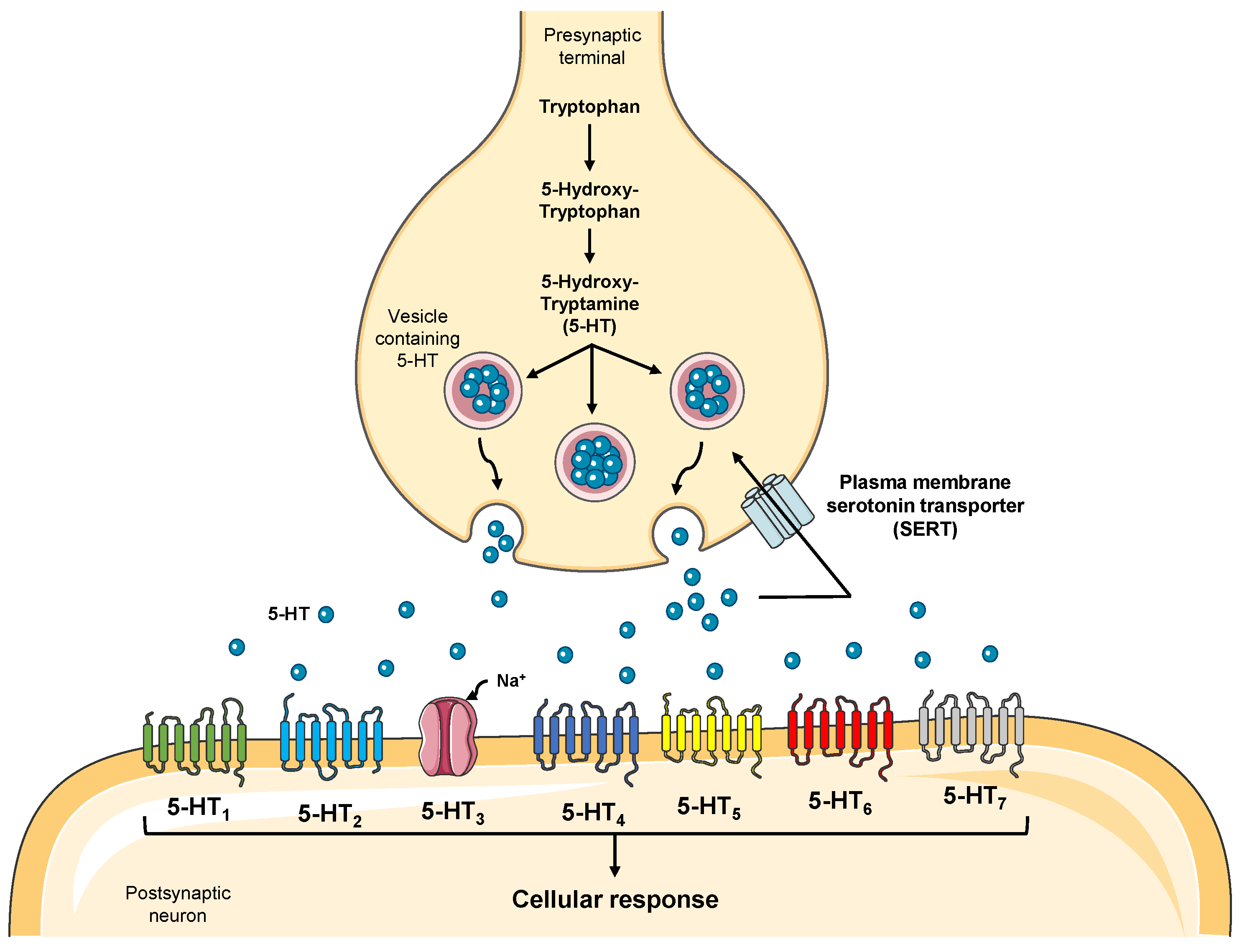
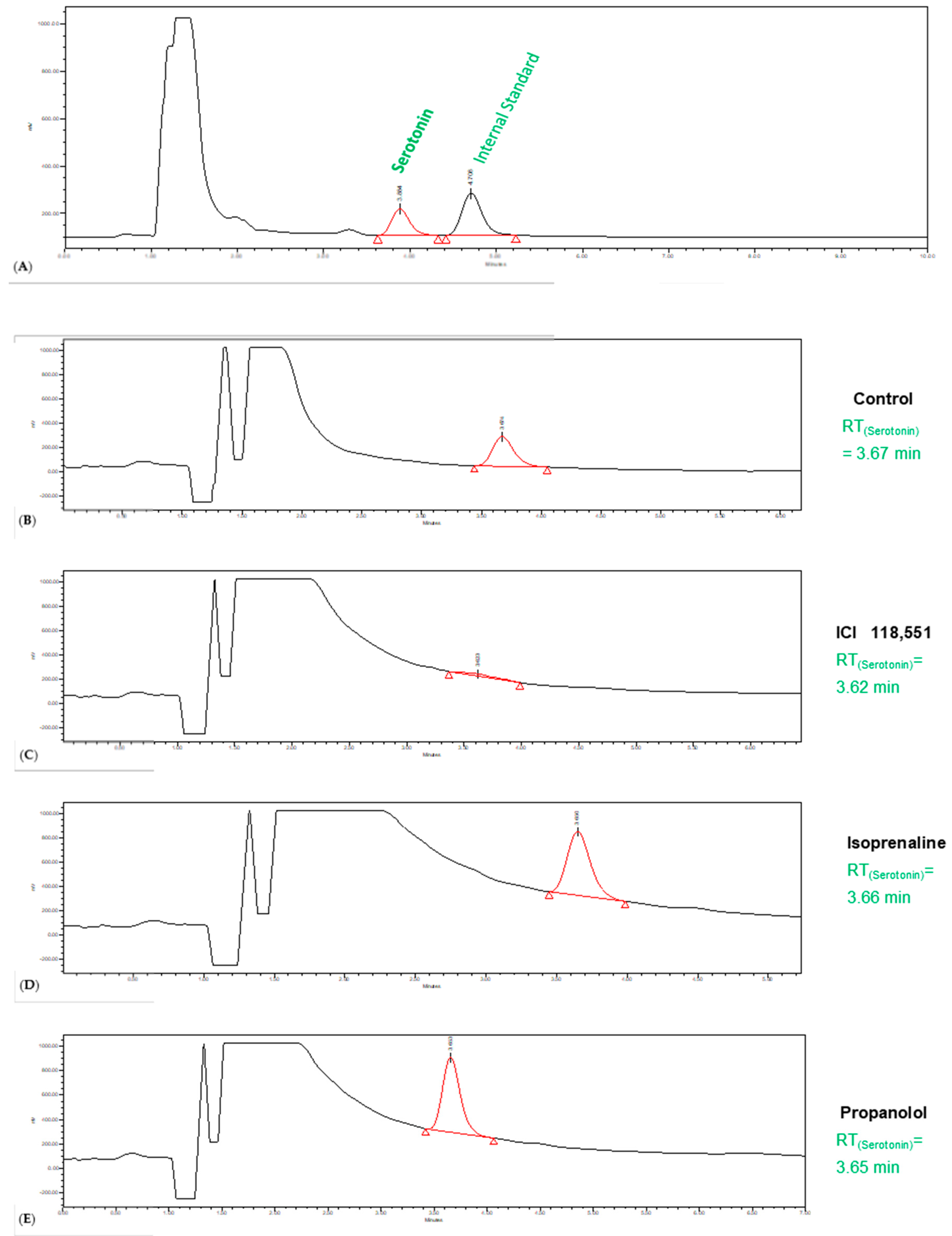
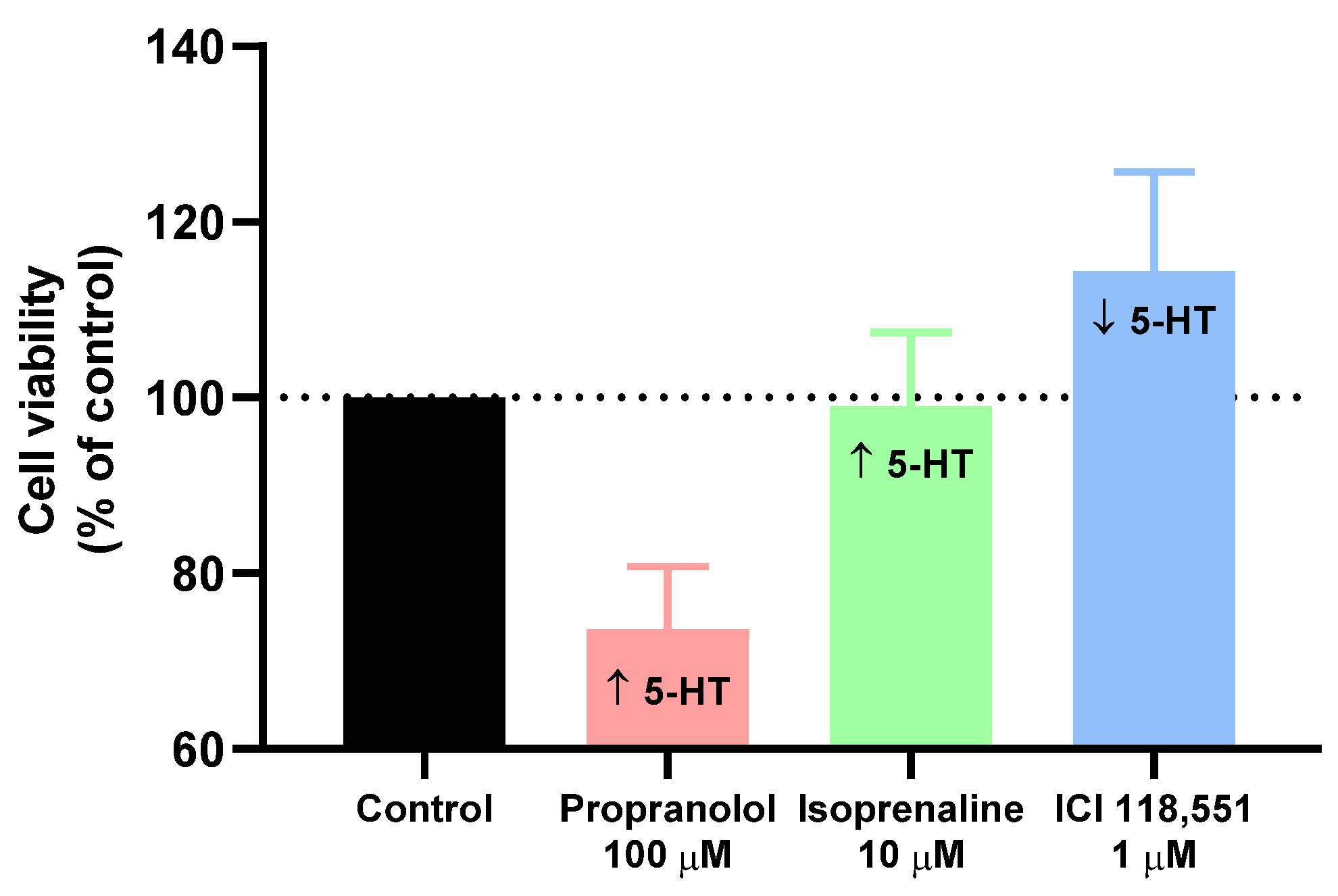
3.4. Effect of Isoprenaline, Propranolol, and ICI 118,551 on MCF-7 Serotonin Production
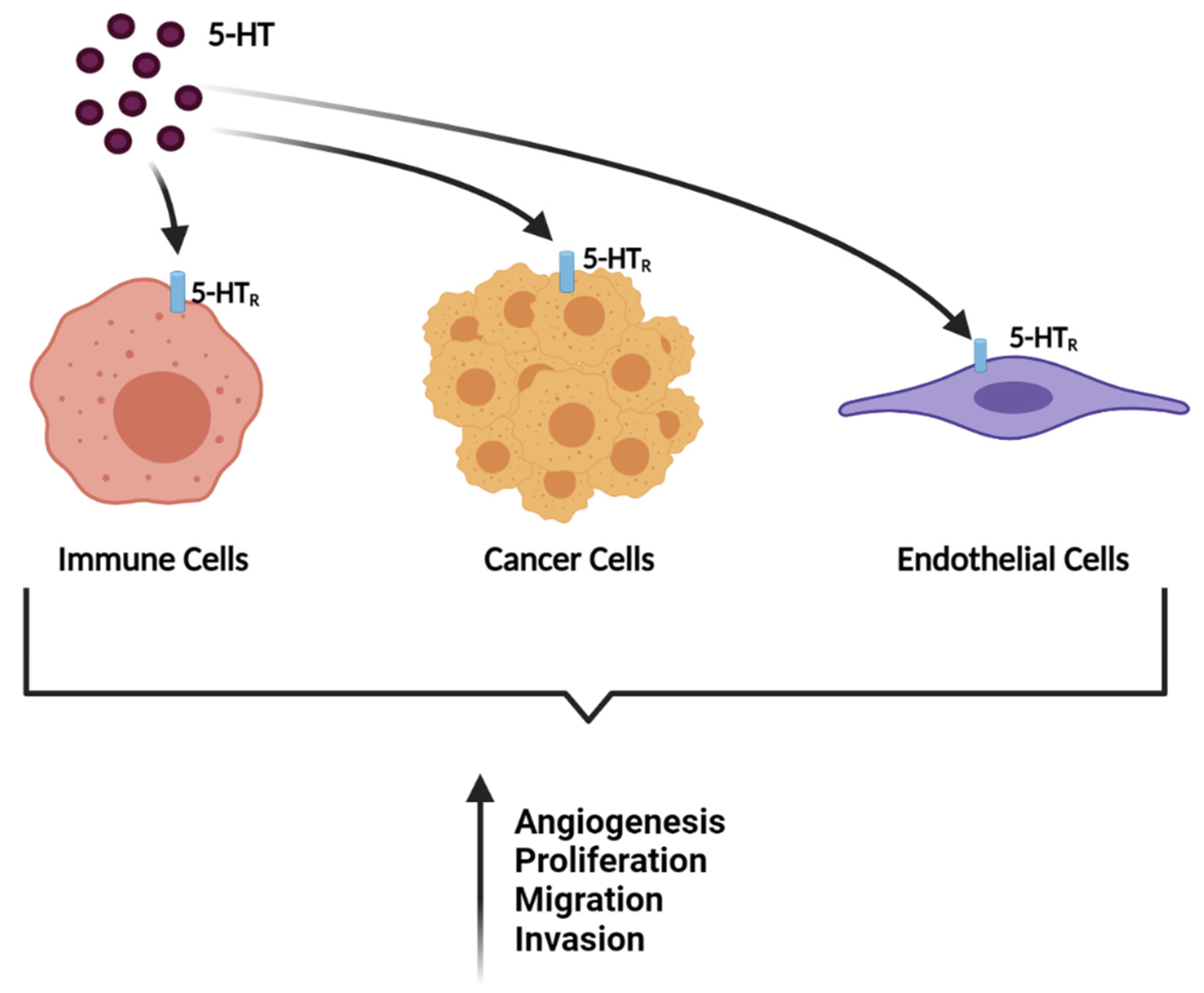
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarrouilhe, D.; Mesnil, M. Serotonin and human cancer: A critical view. Biochimie 2019, 161, 46–50. [Google Scholar] [CrossRef]
- Sarrouilhe, D.; Clarhaut, J.; Defamie, N.; Mesnil, M. Serotonin and Cancer: What Is the Link? Curr. Mol. Med. 2015, 15, 62–77. [Google Scholar] [CrossRef]
- Best, J.; Nijhout, H.F.; Reed, M. Serotonin synthesis, release and reuptake in terminals: A mathematical model. Theor. Biol. Med. Model. 2010, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.K. Serotonin: The Central Link between Bone Mass and Energy Metabolism. In Translational Endocrinology of Bone; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 51–62. ISBN 9780124157842. [Google Scholar]
- Jiang, S.H.; Hu, L.P.; Wang, X.; Li, J.; Zhang, Z.G. Neurotransmitters: Emerging targets in cancer. Oncogene 2020, 39, 503–515. [Google Scholar] [CrossRef]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin regulates pancreatic beta cell mass during pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Chabbi-Achengli, Y.; Coudert, A.E.; Callebert, J.; Geoffroy, V.; Côté, F.; Collet, C.; De Vernejoul, M.C. Decreased osteoclastogenesis in serotonin-deficient mice. Proc. Natl. Acad. Sci. USA 2012, 109, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Dizeyi, N.; Bjartell, A.; Nilsson, E.; Hansson, J.; Gadaleanu, V.; Cross, N.; Abrahamsson, P.A. Expression of Serotonin Receptors and Role of Serotonin in Human Prostate Cancer Tissue and Cell Lines. Prostate 2004, 59, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, E.J.; Shabbir, M.A.; Mikhailidis, D.P.; Mumtaz, F.H.; Thompson, C.S. The effect of serotonin and serotonin antagonists on bladder cancer cell proliferation. BJU Int. 2006, 97, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Nocito, A.; Dahm, F.; Jochum, W.; Jae, H.J.; Georgiev, P.; Bader, M.; Graf, R.; Clavien, P.-A.P.A.; Jang, J.H.; Georgiev, P.; et al. Serotonin Regulates Macrophage-Mediated Angiogenesis in a Mouse Model of Colon Cancer Allografts. Cancer Res. 2008, 68, 5152–5158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, V.P.; Marshall, A.M.; Hernandez, L.L.; Buckley, A.R.; Horseman, N.D. Altered serotonin physiology in human breast cancers favors paradoxical growth and cell survival. Breast Cancer Res. 2009, 11, R81. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Sola-Penna, M.; Paixão, L.P.; Branco, J.R.; Ochioni, A.C.; Albanese, J.M.; Mundim, D.M.; Baptista-de-Souza, D.; Figueiredo, C.P.; Coelho, W.S.; Marcondes, M.C.; et al. Serotonin activates glycolysis and mitochondria biogenesis in human breast cancer cells through activation of the Jak1/STAT3/ERK1/2 and adenylate cyclase/PKA, respectively. Br. J. Cancer 2020, 122, 194–208. [Google Scholar] [CrossRef]
- Sonier, B.; Arseneault, M.; Lavigne, C.; Ouellette, R.J.; Vaillancourt, C. The 5-HT2A serotoninergic receptor is expressed in the MCF-7 human breast cancer cell line and reveals a mitogenic effect of serotonin. Biochem. Biophys. Res. Commun. 2006, 343, 1053–1059. [Google Scholar] [CrossRef]
- Amaro, F.; Silva, D.; Reguengo, H.; Oliveira, J.C.; Quintas, C.; Vale, N.; Gonçalves, J.; Fresco, P. β-adrenoceptor activation in breast mcf-10a cells induces a pattern of catecholamine production similar to that of tumorigenic mcf-7 cells. Int. J. Mol. Sci. 2020, 21, 7968. [Google Scholar] [CrossRef]
- Cole, S.W.; Sood, A.K. Molecular pathways: Beta-adrenergic signaling in cancer. Clin. Cancer Res. 2012, 18, 1201–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargiulo, L.; Copsel, S.; Rivero, E.M.; Galés, C.; Sénard, J.M.; Lüthy, I.A.; Davio, C.; Bruzzone, A. Differential β2-adrenergic receptor expression defines the phenotype of non-tumorigenic and malignant human breast cell lines. Oncotarget 2014, 5, 10058–10069. [Google Scholar] [CrossRef] [PubMed]
- Hothersall, J.D.; Black, J.; Caddick, S.; Vinter, J.G.; Tinker, A.; Baker, J.R. The design, synthesis and pharmacological characterization of novel β 2-adrenoceptor antagonists. Br. J. Pharmacol. 2011, 164, 317–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balakin, K.V. Pharmaceutical Data Mining: Approaches and Applications for Drug Discovery; John Wiley and Sons: Hoboken, NJ, USA, 2009; ISBN 9780470196083. [Google Scholar]
- IUPHAR/BPS Guide to PHARMACOLOGY. Available online: https://www.guidetopharmacology.org/ (accessed on 17 August 2021).
- The European Collection of Authenticated Cell Cultures (ECACC) and Merck. Fundamental Techniques in Cell Culture Laboratory Handbook, 4th ed.; Sigma-Aldrich: St. Louis, MO, USA, 2016. [Google Scholar]
- Risinger, A.L.; Dybdal-Hargreaves, N.F.; Mooberry, S.L. Breast Cancer Cell Lines Exhibit Differential Sensitivities to Microtubule-targeting Drugs Independent of Doubling Time. Anticancer Res. 2015, 35, 5845–5850. [Google Scholar]
- Samaan, T.M.A.; Samec, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Paclitaxel’s Mechanistic and Clinical Effects on Breast Cancer. Biomolecules 2019, 9, 789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayamohanan, H.; Kumar, M.K.M.; Aneesh, T.P. 5-HIAA as a potential biological marker for neurological and psychiatric disorders. Adv. Pharm. Bull. 2019, 9, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Welford, R.W.D.; Vercauteren, M.; Trébaul, A.; Cattaneo, C.; Eckert, D.; Garzotti, M.; Sieber, P.; Segrestaa, J.; Studer, R.; Groenen, P.M.A.; et al. Serotonin biosynthesis as a predictive marker of serotonin pharmacodynamics and disease-induced dysregulation. Sci. Rep. 2016, 6, 30059. [Google Scholar] [CrossRef] [PubMed]
- Lenchner, J.R.; Santos, C. Biochemistry, 5 Hydroxyindoleacetic Acid; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Pantziarka, P.; Bryan, B.A.; Crispino, S.; Dickerson, E.B. Propranolol and breast cancer—A work in progress. Ecancermedicalscience 2018, 12, ed82. [Google Scholar] [CrossRef] [PubMed]
- Caparica, R.; Bruzzone, M.; Agostinetto, E.; De Angelis, C.; Fêde, Â.; Ceppi, M.; de Azambuja, E. Beta-blockers in early-stage breast cancer: A systematic review and meta-analysis. ESMO Open 2021, 6, 100066. [Google Scholar] [CrossRef] [PubMed]
- Montoya, A.; Varela-Ramirez, A.; Dickerson, E.; Pasquier, E.; Torabi, A.; Aguilera, R.; Nahleh, Z.; Bryan, B. The beta adrenergic receptor antagonist propranolol alters mitogenic and apoptotic signaling in late stage breast cancer. Biomed. J. 2019, 42, 155–165. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Chemical Structure | Target | pKi | Action |
|---|---|---|---|---|
| Isoprenaline |  | β1—Adrenoreceptor | 6.6–7.0 | Agonist |
| β2—Adrenoreceptor | 6.4 | Agonist | ||
| Propranolol |  | β1—Adrenoreceptor | 8.6–8.8 | Antagonist |
| β2—Adrenoreceptor | 9.1–9.5 | Antagonist | ||
| β3—Adrenoreceptor | 6.3–7.2 | Antagonist | ||
| 5-HT1A | 7.5 | Antagonist | ||
| 5-HT1B | 5.38 | Antagonist | ||
| ICI 118,551 |  | β1—Adrenoreceptor | 6.6 | Antagonist |
| β2—Adrenoreceptor | 9.2–9.5 | Antagonist | ||
| β3—Adrenoreceptor | 5.8–6.6 | Antagonist |
| IC50 Value/nM | Methodology |
|---|---|
| 2.0 (42) | NR Assay |
| 2.8 (50) | SRB Assay |
| 10.1 (20) | MTT Assay |
| Drug | (5-HT)/nM |
|---|---|
| Propranolol (100 µM) | +147.61 ± 6.20 |
| Isoprenaline (10 µM) | +346.86 ± 9.12 |
| ICI 118,551 (1 µM) | −115.81 ± 4.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, A.S.; Duarte, D.; Silva, I.; Reguengo, H.; Oliveira, J.C.; Vale, N. Serotonin after β-Adrenoreceptors’ Exposition: New Approaches for Personalized Data in Breast Cancer Cells. J. Pers. Med. 2021, 11, 954. https://doi.org/10.3390/jpm11100954
Correia AS, Duarte D, Silva I, Reguengo H, Oliveira JC, Vale N. Serotonin after β-Adrenoreceptors’ Exposition: New Approaches for Personalized Data in Breast Cancer Cells. Journal of Personalized Medicine. 2021; 11(10):954. https://doi.org/10.3390/jpm11100954
Chicago/Turabian StyleCorreia, Ana Salomé, Diana Duarte, Isabel Silva, Henrique Reguengo, José Carlos Oliveira, and Nuno Vale. 2021. "Serotonin after β-Adrenoreceptors’ Exposition: New Approaches for Personalized Data in Breast Cancer Cells" Journal of Personalized Medicine 11, no. 10: 954. https://doi.org/10.3390/jpm11100954