A Novel Nomenclature for Repeat Motifs in the Thymidylate Synthase Enhancer Region and Its Relevance for Pharmacogenetic Studies

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. PCR and Sequence Analysis
2.3. Statistical Analyses
3. Results
3.1. Polymorphisms in the TSER
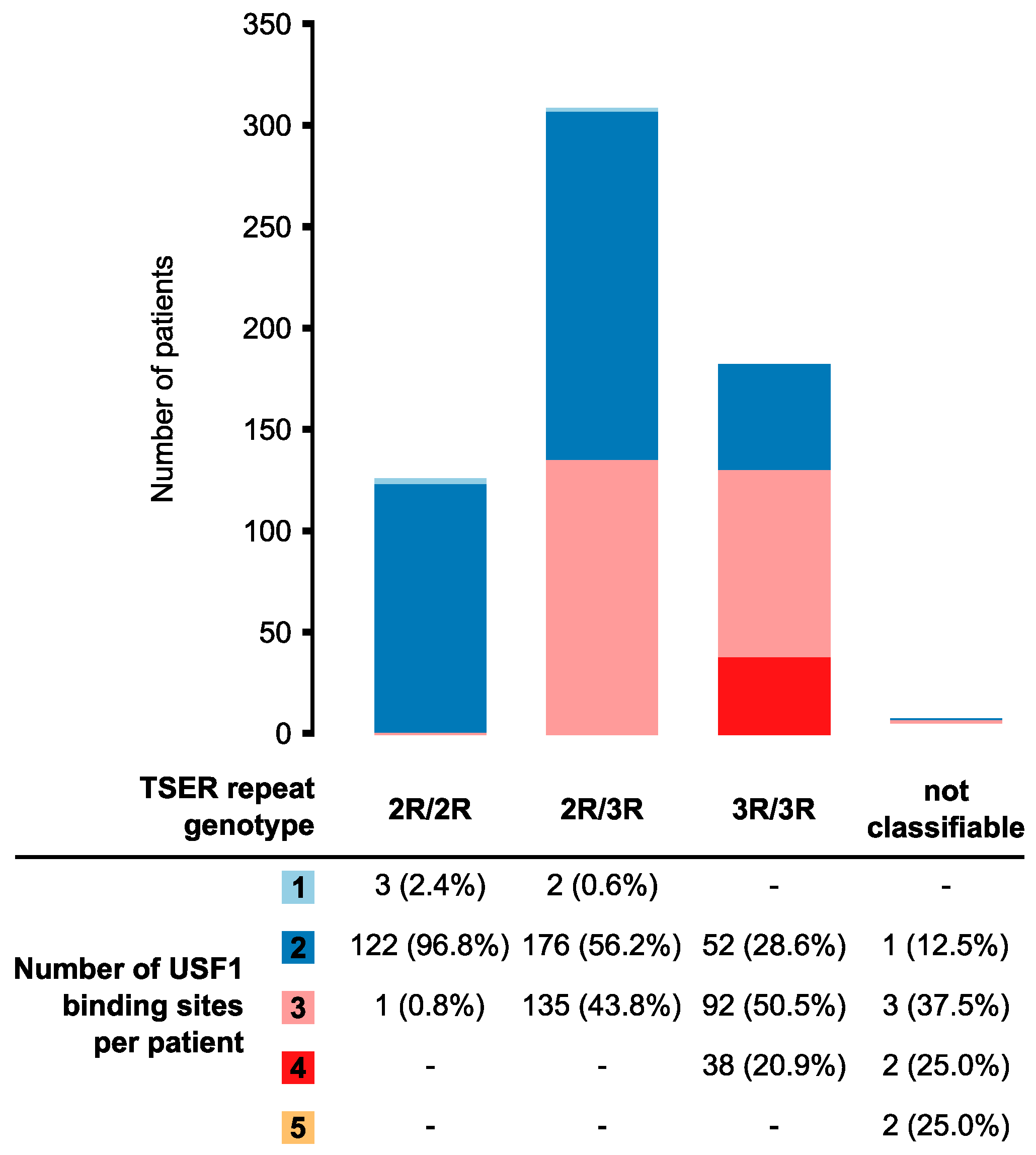
3.2. Distribution of the Number of USF1-Binding Sites Between Different TSER-Repeat Genotypes
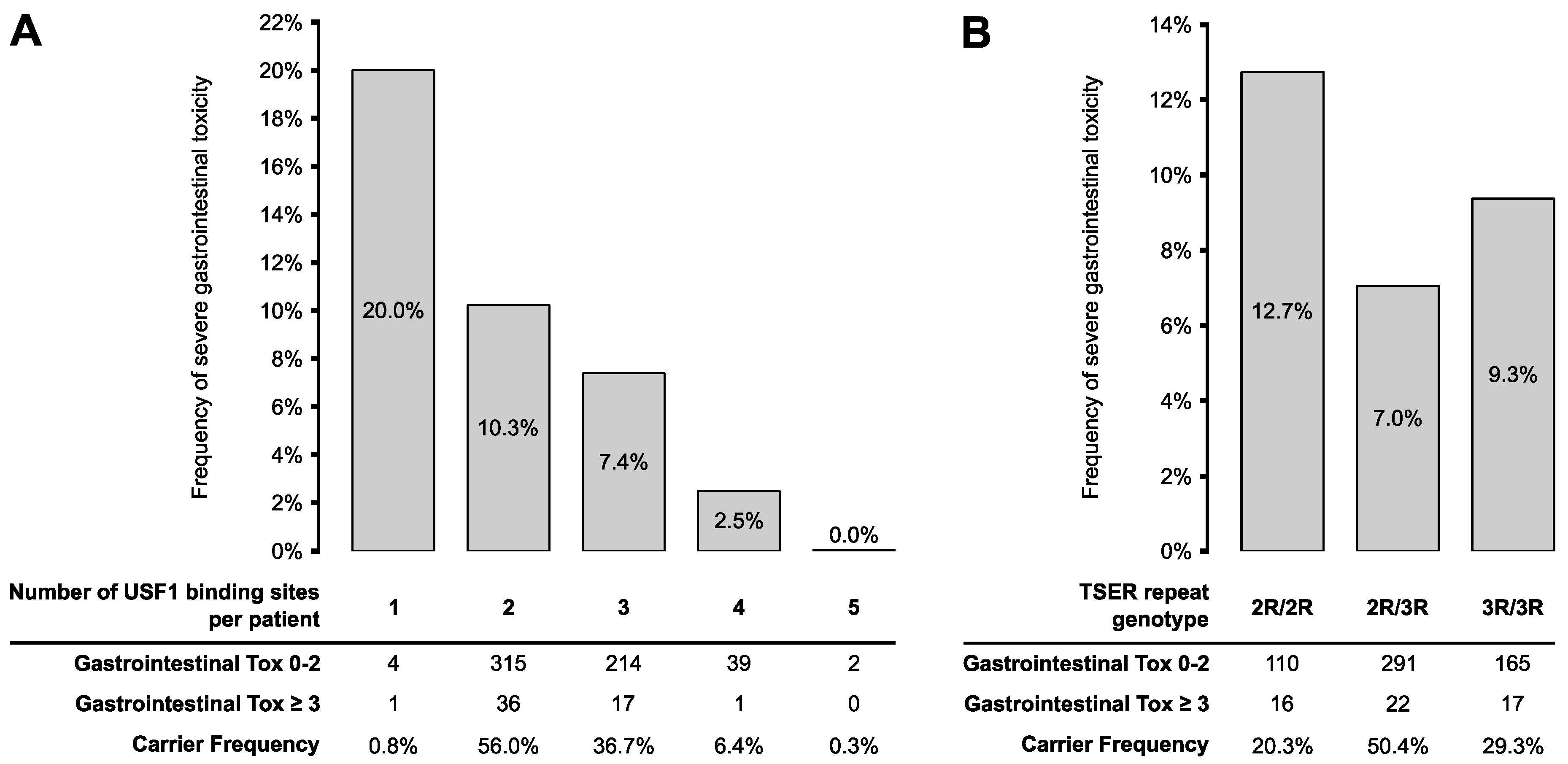
3.3. Association Analyses of TYMS TSER-Variants with Severe 5FU Toxicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Navalgund, L.G.; Rossana, C.; Muench, A.J.; Johnson, L.F.; Gollakota, L.; Rossana, C.; Muench, A.J.; Johnson, L.F. Cell cycle regulation of thymidylate synthetase gene expression in cultured mouse fibroblasts. J. Biol. Chem. 1980, 255, 7386–7390. [Google Scholar] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Meyerhardt, J.A.; Mayer, R.J. Systemic therapy for colorectal cancer. N. Engl. J. Med. 2005, 352, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Horie, N.; Takeishi, K. Identification of functional elements in the promoter region of the human gene for thymidylate synthase and nuclear factors that regulate the expression of the gene. J. Biol. Chem. 1997, 272, 18375–18381. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, S.; Nalbantoglu, J.; Takeishi, K.; Shimizu, K.; Gotoh, O.; Seno, T.; Ayusawa, D. Structural and functional analysis of the human thymidylate synthase gene. J. Biol. Chem. 1990, 265, 20277–20284. [Google Scholar]
- Horie, N.; Takeishi, K.; Aiba, H.; Oguro, K.; Hojo, H. Functional analysis and DNA polymorphism of the tandemly repeated sequences in the 5′-Terminal Regulatory Region of the human gene for thymidylate synthase. Cell Struct. Funct. 1995, 20, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Rosmarin, D.; Palles, C.; Church, D.; Domingo, E.; Jones, A.; Johnstone, E.; Wang, H.; Love, S.; Julier, P.; Scudder, C.; et al. Genetic markers of toxicity from capecitabine and other fluorouracil-based regimens: Investigation in the QUASAR2 study, systematic review, and meta-analysis. J. Clin. Oncol. 2014, 32, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.R.; Ozawa, S.; Saito, Y.; Kurose, K.; Kaniwa, N.; Kamatani, N.; Hamaguchi, T.; Shirao, K.; Muto, M.; Ohtsu, A.; et al. Fourteen novel genetic variations and haplotype structures of the TYMS gene encoding human thymidylate synthase (TS). Drug Metab. Pharmacokinet. 2006, 21, 509–516. [Google Scholar] [CrossRef]
- Marsh, S.; Ameyaw, M.M.; Githang’a, J.; Indalo, A.; Ofori-Adjei, D.; McLeod, H.L. Novel thymidylate synthase enhancer region alleles in African populations. Hum. Mutat. 2000, 16, 528. [Google Scholar] [CrossRef]
- Hamzic, S.; Kummer, D.; Froehlich, T.K.; Joerger, M.; Aebi, S.; Palles, C.; Thomlinson, I.; Meulendijks, D.; Schellens, J.H.M.; García-González, X.; et al. Evaluating the role of ENOSF1 and TYMS variants as predictors in fluoropyrimidine-related toxicities: An IPD meta-analysis. Pharmacol. Res. 2020, 152, 104594. [Google Scholar] [CrossRef]
- Jennings, B.A.; Kwok, C.S.; Willis, G.; Matthews, V.; Wawruch, P.; Loke, Y.K. Functional polymorphisms of folate metabolism and response to chemotherapy for colorectal cancer, a systematic review and meta-analysis. Pharmacogenet. Genom. 2012, 22, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Watanabe, G. Identification and functional analysis of single nucleotide polymorphism in the tandem repeat sequence of thymidylate synthase gene. Cancer Res. 2003, 63, 6004–6007. [Google Scholar] [PubMed]
- Mandola, M.V.; Stoehlmacher, J.; Muller-Weeks, S.; Cesarone, G.; Yu, M.C.; Lenz, H.-J.J.; Ladner, R.D. A novel single nucleotide polymorphism within the 5′ tandem repeat polymorphism of the thymidylate synthase gene abolishes USF-1 binding and alters transcriptional activity. Cancer Res. 2003, 63, 2898–2904. [Google Scholar] [PubMed]
- Gusella, M.; Bolzonella, C.; Crepaldi, G.; Ferrazzi, E.; Padrini, R. A novel G/C single-nucleotide polymorphism in the double 28-bp repeat thymidylate synthase allele. Pharmacogenomics J. 2006, 6, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Meulendijks, D.; Jacobs, B.A.W.; Aliev, A.; Pluim, D.; Van Werkhoven, E.; Deenen, M.J.; Beijnen, J.H.; Cats, A.; Schellens, J.H.M. Increased risk of severe fluoropyrimidine-associated toxicity in patients carrying a G to C substitution in the first 28-bp tandem repeat of the thymidylate synthase 2R allele. Int. J. Cancer 2016, 138, 245–253. [Google Scholar] [CrossRef]
- De Bock, C.E.; Garg, M.B.; Scott, N.; Sakoff, J.A.; Scorgie, F.E.; Ackland, S.P.; Lincz, L.F. Association of thymidylate synthase enhancer region polymorphisms with thymidylate synthase activity in vivo. Pharmacogenomics J. 2011, 11, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ruzzo, A.; Graziano, F.; Loupakis, F.; Rulli, E.; Canestrari, E.; Santini, D.; Catalano, V.; Ficarelli, R.; Maltese, P.; Bisonni, R.; et al. Pharmacogenetic profiling in patients with advanced colorectal cancer treated with first-line FOLFOX-4 chemotherapy. J. Clin. Oncol. 2007, 25, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- McLeod, H.L.; Sargent, D.J.; Marsh, S.; Green, E.M.; King, C.R.; Fuchs, C.S.; Ramanathan, R.K.; Williamson, S.K.; Findlay, B.P.; Thibodeau, S.N.; et al. Pharmacogenetic predictors of adverse events and response to chemotherapy in metastatic colorectal cancer: Results from North American gastrointestinal intergroup trial N9741. J. Clin. Oncol. 2010, 28, 3227–3233. [Google Scholar] [CrossRef]
- Lecomte, T.; Ferraz, J.M.; Zinzindohoué, F.; Loriot, M.A.; Tregouet, D.A.; Landi, B.; Berger, A.; Cugnenc, P.H.; Jian, R.; Beaune, P.; et al. Thymidylate synthase gene polymorphism predicts toxicity in colorectal cancer patients receiving 5-fluorouracil-based chemotherapy. Clin. Cancer Res. 2004, 10, 5880–5888. [Google Scholar] [CrossRef] [Green Version]
- Nief, N.; Le Morvan, V.; Robert, J. Involvement of gene polymorphisms of thymidylate synthase in gene expression, protein activity and anticancer drug cytotoxicity using the NCI-60 panel. Eur. J. Cancer 2007, 43, 955–962. [Google Scholar] [CrossRef]
- Marcuello, E.; Altés, A.; Del Rio, E.; César, A.; Menoyo, A.; Baiget, M. Single nucleotide polymorphism in the 5’ tandem repeat sequences of Thymidylate synthase gene predicts for response to fluorouracil-based chemotherapy in advanced colorectal cancer patients. Int. J. Cancer 2004, 112, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Hoskins, J.M.; Rivory, L.P.; Zucknick, M.; London, R.; Liddle, C.; Clarke, S.J. Thymidylate synthase and methylenetetrahydrofolate reductase gene polymorphisms and toxicity to capecitabine in advanced colorectal cancer patients. Clin. Cancer Res. 2008, 14, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, A.; Nielsen, J.N.; Gyldenkerne, N.; Lindeberg, J. Thymidylate synthase and methylenetetrahydrofolate reductase gene polymorphism in normal tissue as predictors of fluorouracil sensitivity. J. Clin. Oncol. 2005, 23, 1365–1369. [Google Scholar] [CrossRef] [PubMed]
- Pullarkat, S.T.; Stoehlmacher, J.; Ghaderi, V.; Xiong, Y.-P.; Ingles, S.A.; Sherrod, A.; Warren, R.; Tsao-Wei, D.; Groshen, S.; Lenz, H.-J.J. Thymidylate synthase gene polymorphism determines response and toxicity of 5-FU chemotherapy. Pharmacogenomics J. 2001, 1, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joerger, M.; Huitema, A.D.R.R.; Boot, H.; Cats, A.; Doodeman, V.D.; Smits, P.H.M.M.; Vainchtein, L.; Rosing, H.; Meijerman, I.; Zueger, M.; et al. Germline TYMS genotype is highly predictive in patients with metastatic gastrointestinal malignancies receiving capecitabine-based chemotherapy. Cancer Chemother. Pharmacol. 2015, 75, 763–772. [Google Scholar] [CrossRef]
- Wu, Q.; Dolnick, B.J. Detection of thymidylate synthase modulators by a novel screening assay. Mol. Pharmacol. 2003, 63, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Froehlich, T.K.; Amstutz, U.; Aebi, S.; Joerger, M.; Largiadèr, C.R. Clinical importance of risk variants in the dihydropyrimidine dehydrogenase gene for the prediction of early-onset fluoropyrimidine toxicity. Int. J. Cancer 2015, 136, 730–739. [Google Scholar] [CrossRef]
- NCI USNCI Cancer Therapy Evaluation Program: Common Terminology Criteria for Adverse Events (CTCAE) v3.0. Available online: https://ctep.cancer.gov/protocoldevelopment/electronic_applications/docs/CTCAE_v5_Quick_Reference_8.5x11.pdf (accessed on 4 February 2020).
- Rousset, F. Genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Res. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Amstutz, U.; Henricks, L.M.; Offer, S.M.; Barbarino, J.; Schellens, J.H.M.; Swen, J.J.; Klein, T.E.; McLeod, H.L.; Caudle, K.E.; Diasio, R.B.; et al. Clinical pharmacogenetics implementation consortium (CPIC) guideline for dihydropyrimidine dehydrogenase genotype and fluoropyrimidine dosing: 2017 update. Clin. Pharmacol. Ther. 2018, 103, 210–216. [Google Scholar] [CrossRef]
- Marsh, S.; Collie-Duguid, E.S.R.; Li, T.; Liu, X.; McLeod, H.L. Ethnic variation in the thymidylate synthase enhancer region polymorphism among Caucasian and Asian populations. Genomics 1999, 58, 310–312. [Google Scholar] [CrossRef]
- Luo, H.R.; Lü, X.M.; Yao, Y.G.; Horie, N.; Takeishi, K.; Jorde, L.B.; Zhang, Y.P. Length polymorphism of thymidylate synthase regulatory region in Chinese populations and evolution of the novel alleles. Biochem. Genet. 2002, 40, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Loganayagam, A.; Arenas Hernandez, M.; Corrigan, A.; Fairbanks, L.; Lewis, C.M.; Harper, P.; Maisey, N.; Ross, P.; Sanderson, J.D.; Marinaki, A.M. Pharmacogenetic variants in the DPYD, TYMS, CDA and MTHFR genes are clinically significant predictors of fluoropyrimidine toxicity. Br. J. Cancer 2013, 108, 2505–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, M.; Zanger, U.M.; Marx, C.; Schaeffeler, E.; Klein, K.; Dippon, J.; Kerb, R.; Blievernicht, J.; Fischer, J.; Hofmann, U.; et al. Role of genetic and nongenetic factors for fluorouracil treatment-related severe toxicity: A prospective clinical trial by the German 5-FU toxicity study group. J. Clin. Oncol. 2008, 26, 2131–2138. [Google Scholar] [CrossRef]
- Petrelli, F.; Cabiddu, M.; Barni, S. 5-Fluorouracil or capecitabine in the treatment of advanced colorectal cancer: A pooled-analysis of randomized trials. Med. Oncol. 2012, 29, 1020–1029. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |




{kind=link}
{kind=link}
{kind=link}
| Grade 3–5 | Grade 0–2 | Total | |||
|---|---|---|---|---|---|
| N | % | N | % | N | |
| Total | |||||
| Total cohort | 106 | 17 | 523 | 83 | 629 |
| Collection | |||||
| Prospectively | 92 | 15 | 522 | 85 | 614 |
| Retrospectively | 14 | 93 | 1 | 7 | 15 |
| Sex | |||||
| Female | 55 | 22 | 192 | 78 | 247 |
| Male | 51 | 13 | 331 | 87 | 382 |
| Ancestry | |||||
| Caucasian | 104 | 17 | 516 | 83 | 620 |
| Arab | 0 | 0 | 3 | 100 | 3 |
| African | 1 | 50 | 1 | 50 | 2 |
| Asian | 0 | 0 | 2 | 100 | 2 |
| Unknown | 1 | 50 | 1 | 50 | 2 |
| Treatment | |||||
| FOLFOX, FOLFIRI | 22 | 10 | 192 | 90 | 214 |
| 5FU +/−LV | 12 | 11 | 100 | 89 | 112 |
| 5FU, CPL +/−D,E | 36 | 30 | 86 | 70 | 122 |
| Other 5FU regimen | 10 | 28 | 26 | 72 | 36 |
| Cp | 26 | 18 | 119 | 82 | 145 |
| Toxicity category | |||||
| Hematologic toxicity | 58 | 9 | 571 | 91 | 629 |
| Gastrointestinal toxicity | 55 | 9 | 574 | 91 | 629 |
| Infection | 24 | 4 | 605 | 96 | 629 |
| Dermatologic toxicity | 15 | 2 | 614 | 98 | 629 |
| New Nomenclature | Repeat Number (RN-) Nomenclature | Repeat Number, Binding Site, SNP (RNBS-) Nomenclature | USF1-Binding Sites N | Allele N | Allele f% | Kim et al. f% | p-Value |
|---|---|---|---|---|---|---|---|
| a1+b1 | 2R | 2RG | 1 | 561 | 44.6 | 12.4 | <0.0001 |
| a1+a2b1 | 3R | 3RC | 1 | 360 | 28.6 | 42.0 | <0.0001 |
| a1+a1+b1 | 3R | 3RG | 2 | 308 | 24.5 | 42.1 | <0.0001 |
| a1+b1b1 | 3R | 3RC-ins * | 1 | 12 | 1.0 | 0.2 | ns |
| a2b1 | 2R | 2RC | 0 | 6 | 0.5 | - | - |
| a1+a1+a1+b1 | 4R | na | 3 | 4 | 0.3 | - | - |
| a1+b2 | 2R | 2RG | 2 | 1 | 0.1 | - | - |
| a1+a1+a2b1 | 4R | 4RC * | 2 | 1 | 0.1 | 0.2 | ns |
| a1+a2a2b1 | 4R | na | 1 | 1 | 0.1 | - | - |
| a1+a1+a2a2b1 | 5R | na | 2 | 1 | 0.1 | - | - |
| a1+a2b1a2b1 | 5R | na | 1 | 1 | 0.1 | - | - |
| a1+γ1 | 2R | 2RG | 1 | 1 | 0.1 | - | - |
| b1b1 | 2R | 2RC | 0 | 1 | 0.1 | - | - |
| a1+a1+a1+a2b1 | 5R | 5RC * | 3 | 0 | 0.0 | 2.1 | <0.0001 |
| Univariable | Multivariable 1 | |||||
|---|---|---|---|---|---|---|
| Outcome | OR | 95% CI | p-Value | OR | 95% CI | p-Value |
| Overall toxicity | 1.04 | 0.76–1.46 | 0.793 | 1.08 | 0.77–1.54 | 0.670 |
| Hematologic toxicity | 0.73 | 0.49–1.10 | 0.122 | 0.73 | 0.48–1.13 | 0.148 |
| Gastrointestinal toxicity | 1.66 | 1.04–2.77 | 0.043 | 1.74 | 1.06–2.99 | 0.034 |
| Infection | 1.1 | 0.59–2.20 | 0.769 | 1.14 | 0.57–2.43 | 0.723 |
| Dermatologic toxicity | 1.29 | 0.58–3.27 | 0.558 | 1.28 | 0.57–3.25 | 0.576 |
| Univariable | Multivariable 1 | |||||
|---|---|---|---|---|---|---|
| Outcome | OR | 95% CI | p-Value | OR | 95% CI | p-Value |
| Overall toxicity | 0.96 | 0.71–1.30 | 0.814 | 0.98 | 0.71–1.34 | 0.887 |
| Hematological toxicity | 0.73 | 0.49–1.08 | 0.119 | 0.75 | 0.49–1.13 | 0.174 |
| Gastrointestinal toxicity | 1.18 | 0.79–1.75 | 0.424 | 1.21 | 0.81–1.82 | 0.348 |
| Infection | 0.93 | 0.51–1.66 | 0.803 | 1.06 | 0.58–1.93 | 0.845 |
| Dermatological toxicity | 1.05 | 0.50–2.19 | 0.895 | 0.97 | 0.45–2.04 | 0.929 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaerer, D.; Froehlich, T.K.; Hamzic, S.; Offer, S.M.; Diasio, R.B.; Joerger, M.; Amstutz, U.; Largiadèr, C.R. A Novel Nomenclature for Repeat Motifs in the Thymidylate Synthase Enhancer Region and Its Relevance for Pharmacogenetic Studies. J. Pers. Med. 2020, 10, 181. https://doi.org/10.3390/jpm10040181
Schaerer D, Froehlich TK, Hamzic S, Offer SM, Diasio RB, Joerger M, Amstutz U, Largiadèr CR. A Novel Nomenclature for Repeat Motifs in the Thymidylate Synthase Enhancer Region and Its Relevance for Pharmacogenetic Studies. Journal of Personalized Medicine. 2020; 10(4):181. https://doi.org/10.3390/jpm10040181
Chicago/Turabian StyleSchaerer, Dominic, Tanja K. Froehlich, Seid Hamzic, Steven M. Offer, Robert B. Diasio, Markus Joerger, Ursula Amstutz, and Carlo R. Largiadèr. 2020. "A Novel Nomenclature for Repeat Motifs in the Thymidylate Synthase Enhancer Region and Its Relevance for Pharmacogenetic Studies" Journal of Personalized Medicine 10, no. 4: 181. https://doi.org/10.3390/jpm10040181