The Bluegreen Algae (AFA) Consumption over 48 h Increases the Total Number of Peripheral CD34+ Cells in Healthy Patients: Effect of Short-Term and Long-Term Nutritional Supplementation (Curcumin/AFA) on CD34+ Levels (Blood)

Abstract
:1. Introduction
Aim
2. Material and Methods
2.1. Clinical Protocol for Patients
2.1.1. Inclusion/Exclusion Criteria
- -
- AFA bluegreen algae-treated healthy subjects over 48 consecutive hours (n = 5).
- -
- Powder curcumin-treated subjects over 48 consecutive hours (n = 5).
- -
- Liquid curcumin-treated subjects over 48 consecutive hours (n = 5)
- -
- Placebo-treated group (n = 7).
- -
- The liquid curcumin was administered over seven consecutive days (cur 7 days) or 38 consecutive days of administration in healthy subjects (cur 38 days, n = 5). The administration of curcumin take place in the same patient until reaching 38 days of supplementation. Thus, blood samples were collected at both selected times (seven days as well as 38 days, n = 5).
- -
- Placebo-treated group (n = 7).
- -
- Glycosinolate of sulforaphane (NutriSGS without C vitamin) from Cienporciennatural company (Madrid, Spain, see protocol Figure 1).
2.1.2. Exclusion Criteria
2.2. Protocol for Nutritional Supplementation in Healthy Subjects
2.2.1. Short-Term Nutritional Supplementation
2.2.2. Long-Term Nutritional Supplementation
2.3. Protocol for Peripheral Blood Extraction
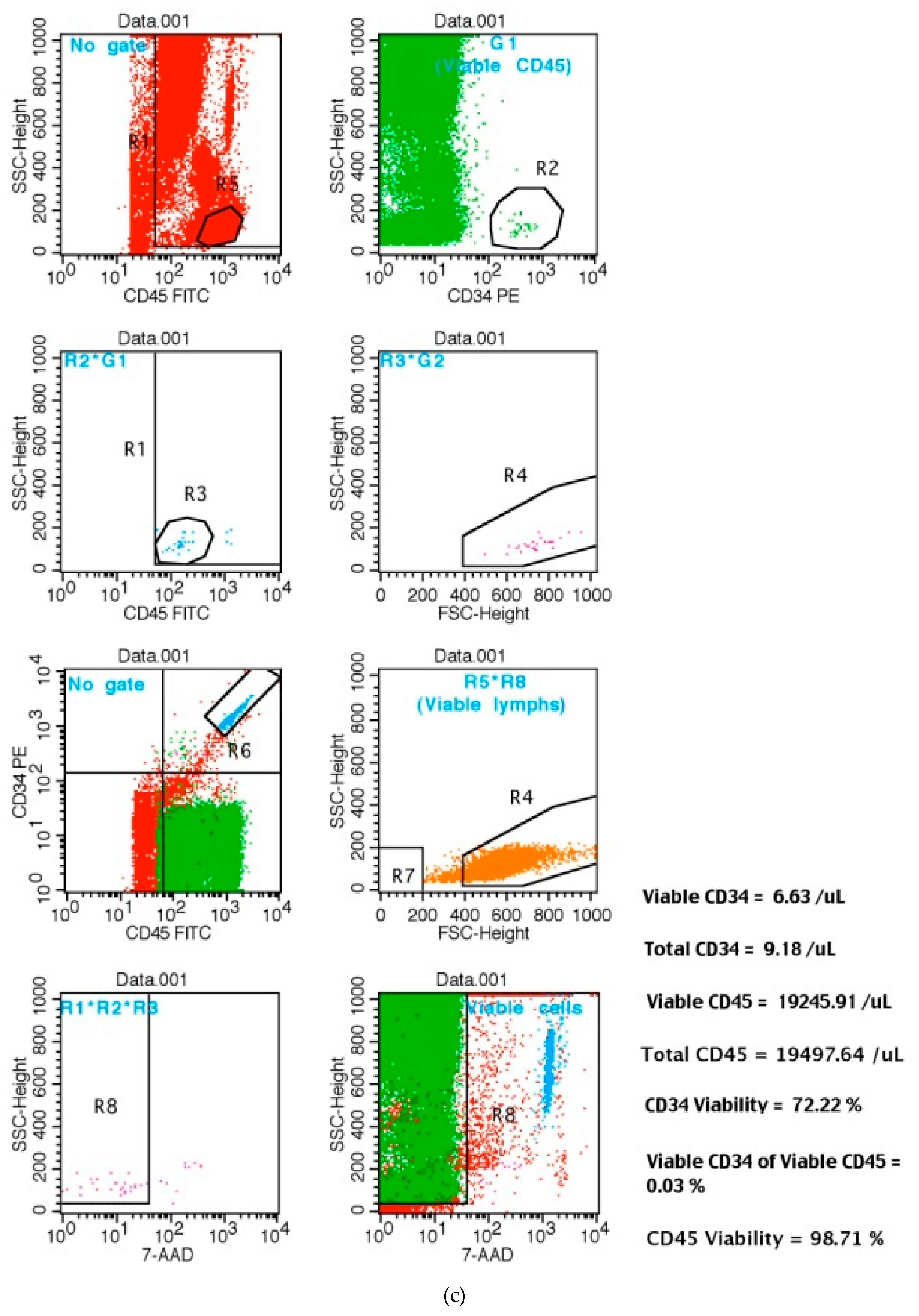
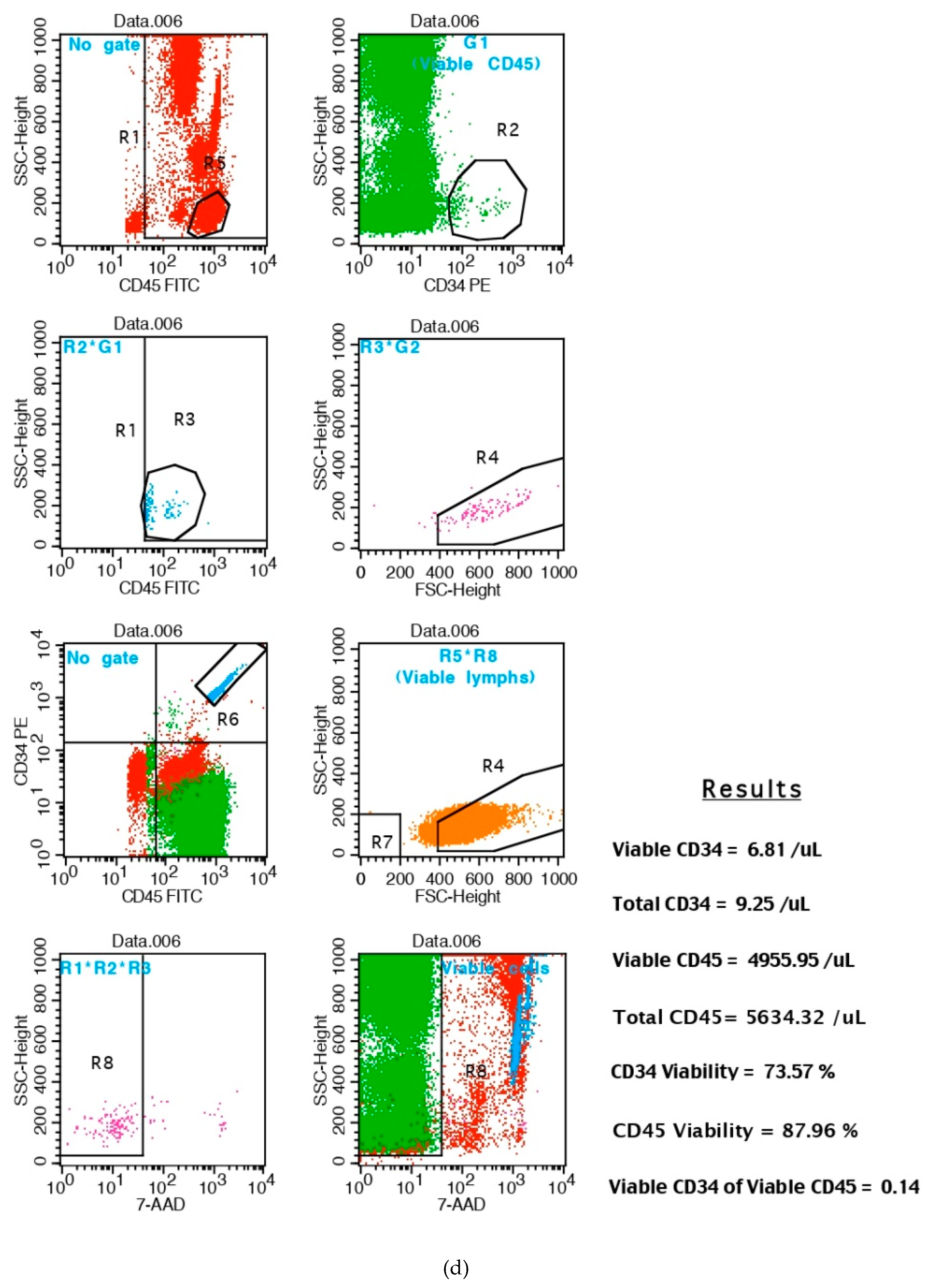
2.4. Quantification of Total Peripheral CD34+ Positive Cells by Flow Cytometry: CD34+ Is a Marker of Hematopoietic Stem Cells (HSC) Mobilization in Patients
2.5. Statistical Analysis
3. Results: CD34+ Hematopoyetic Stem Cell (HSC) Mobilization in Healthy Subjects
3.1. Protocol for Short-Term Nutritional Supplementation in Healthy Patients (Figure 2)
3.1.1. AFA Bluegreen Algae Extract Consumption Increased the Total Number of Peripheral CD34+-HSC Cells without Affecting Their Circulating Levels in Curcumin–Treated Subjects
3.1.2. Curcumin Treatment (Powder or Liquid Forms) Did Not Increase the Total Number of CD34+ Peripheral Cells in Healthy Subjects
3.2. The Long-Term Nutritional Supplementation with Curcumin, Glycosinolate of Sulforaphane, and AFA Bluegreen Algae Extract Increased the Total Number of CD34+ Peripheral Cells in Healthy Subjects
3.3. Increased Number of CD34+ That Are Also CD45+ Cells, after 48 h of AFA Consumption or Curcumin (Powder Form) Administration over 24 h in Healthy Subjects
3.4. The Number of Nucleated Cells (TNC) or White Blood Cells (WBC) Did Not Differ between Healthy-Treated Subjects
4. Discussion
5. Conclusions and Further Considerations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Osawa, M.; Hanada, K.; Hamada, H.; Nakauchi, H. Long-term lymphohematopoietic reconstitution by a single CD34-low/negative hematopoietic stem cell. Science 1996, 273, 242–245. [Google Scholar] [CrossRef] [Green Version]
- Laurenti, E.; Gottgens, B. From haematopoietic stem cells to complex differentiation landscapes. Nature 2018, 553, 418–426. [Google Scholar] [CrossRef]
- Drapeau, C.; Eugemio, G.; Mazzoni, P.; Toth, G.D.; Stranber, S. The therapeutical potential of stimulating endogenoud stem cell mobilization. In Tech Open. Basic Biol. Clin. Appl. 2012, 8, 167–202. [Google Scholar]
- Viswanathan, C.; Kulkarni, R.; Bopardikar, A.; Ramdasi, S. Significance of CD34 negative hematopoietic stem cells and CD34 positive mesenchymal stem cells—A valuable dimension to the current understanding. Curr. Stem Cell Res. Ther. 2017, 12, 476–483. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, Y.; Miao, W.; Cheng, T. Reactive Oxygen Species and Nrf2: Functional and Transcriptional Regulators of Hematopoiesis. Oxid. Med. Cell. Longev. 2019, 18. [Google Scholar] [CrossRef]
- Lemery, S.; Hsieh, M.M.; Smith, A.; Rao, S.; Khuu, H.M.; Donohue, T.; Viano, J.M.; Cook, L.; Goodwin, R.; Boss, C.; et al. A pilot study evaluating the safety and CD34+ cell mobilizing activity of escalating doses of plerixafor in healthy volunteers. Br. J. Hematol. 2011, 153, 66–75. [Google Scholar] [CrossRef]
- Rauscher, F.M.; Goldschmidt-Clermont, P.J.; Davis, B.H.; Wang, T.; Gregg, D.; Ramaswami, P.; Pippen, A.M.; Annex, B.H.; Dong, C.; Taylor, D.A. Aging, progenitor cell exhaustion, and atherosclerosis. Circulation 2003, 108, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Fadini, G.P.; Ciciliot, S.; Albiero, M. Concise review: Perspective and clinical implications of bone marrow and circulating stem cells defects in diabetes. Stem Cells 2017, 35, 106–116. [Google Scholar] [CrossRef]
- Mazzanti, G.; Di Giacom, S. Curcumin and resveratrol in the management of cognitive disorders: What is the clinical evidence? Molecules 2016, 21, 1243. [Google Scholar] [CrossRef] [Green Version]
- Weissman, I.L.; Anderson, D.J.; Gage, F. Stem and progenitor cells: Origins, phenotypes, lineage commitments, and transdifferentiations. Annu. Rev. Cell Dev. Biol. 2001, 17, 387–403. [Google Scholar] [CrossRef] [Green Version]
- Small, G.W.; Siddarth, P.; Li, Z.; Miller, K.J.; Ercoli, L.; Emerson, N.; Martinez, J.; Koon-Pong, W.; Liu, J.; Merril, D.A.; et al. Memory and brain amyloid and tau effects of a bioavailable form of curcumin in non-demented adults: A double blind, placebo-controlled 18-month trial. Am. J. Geriatr. Psychiatry 2018, 26, 266–277. [Google Scholar] [CrossRef]
- Braak, E.H.; Braak, E. Neuropathological staging of Alzheimer-related changes. Acta Neuropathol. (Berl) 1991, 82, 239–259. [Google Scholar] [CrossRef]
- Benedict Andrea, L.; Mountney, A.; Hurtado, A.; Kelley, E.; Bryan Ronald, L.; Schnaar Albena, T.; Dinkova-Kostova Talalay, P. Neuroprotective Effects of Sulforaphane after Contusive Spinal Cord Injury. J. Neurotrauma 2012, 29, 2586. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Hirao, A.; Arai, F.; Matsuoka, S.; Takubo, K.; Hamaguchi, I.; Nomiyama, K.; Hosokawa, K.; Sakurada, K.; Nakagata, N.; et al. Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature 2004, 431, 997–1002. [Google Scholar] [CrossRef]
- Tothova, Z.; Kollipara, R.; Huntly, B.J.; Lee, B.H.; Castrillon, D.H.; Cullen, D.E.; McDowell, E.P.; Lazo-Kallanian, S.; Williams, I.R.; Cristopher, S.; et al. FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 2007, 128, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Chai, X.; Li, D.; Cao, X.M.; Zhang, Y.; Mu, J.; Lu, W.; Xiao, X.; Li, C.; Meng, J.; Chen, J.; et al. ROS-mediated iron overload injures the hematopoiesis of bone marrow by damaging hematopoietic stem/progenitor cells in mice. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhai, Z.; Wang, Y.; Zhang, J.; Wu, H.; Wang, Y.; Li, C.; Li, D.; Lu, L.; Wang, X.; et al. Resveratrol ameliorates ionizing irradiation-induced long-term hematopoietic stem cell injury in mice. Free Radic. Biol. Med. 2013, 54, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Tian, Z.; Tang, W.M.; Zhang, J.; Lu, L.; Sun, Z.; Zhou, Z.; Fan, F. The Protective Effects of 5-Methoxytryptamine-α-lipoic Acid on Ionizing Radiation-Induced Hematopoietic Injury. Int. J. Mol. Sci. 2016, 17, 935. [Google Scholar] [CrossRef] [Green Version]
- Suryavanshi, S.; Sharma, D.; Checker, R.; Maikho, T.; Vikram, G.; Santosh, K.S.; Sainis, K.B. Amelioration of radiation-induced hematopoietic syndrome by an antioxidant chlorophyllin through increased stem cell activity and modulation of hematopoiesis. Free Radic. Biol. Med. 2015, 85, 56–70. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Y.L.; Zhang, H.; Yan, H.; Wu, X.J.; Zhang, C.Z. Administration of the resveratrol analogues isorhapontigenin and heyneanol—A protects mice hematopoietic cells against irradiation injuries. BioMed Res. Int. 2014, 2014, 282657. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cai, D.; Yao, X.; Zhang, Y.; Chen, L.; Jing, P.; Wang, L.; Wang, Y. Protective effect of ginsenoside Rg1 on hematopoietic stem/progenitor cells through attenuating oxidative stress and the Wnt/β-catenin signaling pathway in a mouse model of d-galactose-induced aging. Int. J. Mol. Sci. 2016, 17, 849. [Google Scholar] [CrossRef] [PubMed]
- Drapeau, C.; Benson, K.F.; Jensen, G.S. Rapid and selective mobilization of specific stem cell types after consumption of a polyphenol-rich extract from sea buckthorn berries (Hippophae) in healthy human subjects. Clin. Interv. Aging 2019, 14, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Cheng, H.; Gao, Y.; Ming, S.; Yanfeng, L.; Zheng, H.; Jing, X.; Lugui, Q.; Weiping, Y.; Leung, A.Y.-H.; et al. Antioxidant N-acetyl-L-cysteine increases engraftment of human hematopoietic stem cells in immune-deficient mice. Blood 2014, 124, e45–e48. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Wang, Y.; Zhang, Y.Y.; Shi, M.-M.; Mo, X.-D.; Sun, Y.-Q.; Chang, Y.-J.; Xu, L.-P.; Zhang, X.-H.; Liu, K.-Y.; et al. Prophylactic oral NAC reduced poor hematopoietic reconstitution by improving endothelial cells after haploidentical transplantation. Blood Adv. 2019, 3, 1303–1317. [Google Scholar] [CrossRef]
- Xian, Y.; Lin, Y.; Cao, C.; Li, L.; Wang, J.; Niu, J.; Guo, Y.; Sun, Y.; Wang, Y.; Wang, W. Protective effect of umbilical cord mesenchimal stem cells combined with resveratrol against renal podocyte damage in NOD mice. Diabetes Res. Clin. Pract. 2011, 156, 107755. [Google Scholar] [CrossRef]
- Rimmele, P.; Lofek-Czubek, S.; Ghaffari, S. Resveratrol increases the bone marrow hematopoietic stem and progenitor cell capacity. Am. J. Hematol. 2014, 89, E235–E238. [Google Scholar] [CrossRef] [Green Version]
- Rainey-Smith, S.R.; Brown, B.M.; Sohrabi, H.R.; Shah, T.; Goozee, K.G.; Gupta, V.B.; Martins, R.N. Curcumin and cognition: A randomised, placebo-controlled, double-blind study of community-dwelling older adults. Br. J. Nutr. 2016, 115, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.H.; Pipingas, A.; Scholey, A.B. Investigation of the effects of solid lipid curcumin on cognition and mood in a healthy older population. J. Psychopharmacol. 2015, 29, 642–651. [Google Scholar] [CrossRef]
- Sasaki, H.; Sunagawa, Y.; Takahashi, K.; Imaizumi, A.; Fukuda, H.; Hashimoto, T.; Wada, H.; Katanasaka, Y.; Kakeya, H.; Fujita, M.; et al. Innovative preparation of curcumin for improved oral bioavailability. Biol. Pharm. Bull. 2011, 34, 660–665. [Google Scholar] [CrossRef] [Green Version]
- Merino, J.J.; Parmigiani-Izquierdo, J.M.; Toledano Gasca, A.; Cabaña-Muñoz, M.E. The long-term algae extract (Chlorella and Fucus) and aminosulphurate supplumentation modulate SOD-1 activity and decrease heavy metal (Hg++, Sn) in patients with long-term dental titanium implant and amalgam fillings. Antioxidants (Basel) 2019, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Xu, Q.; Li, C.; Zhao, H. Effects of sulforaphane on neural stem cell proliferation and differentiation. Genesis 2017, 55, 1–6. [Google Scholar] [CrossRef]
- Ping, Z.; Liu, W.W.; Kang, Z.M.; Cai, J.M.; Wang, Q.S.; Chebg, N.; Sun, X.J. Sulforaphane protects brains against hypoxic-ischemic injury through induction of Nrf-2-dependent phase 2 enzyme. Brain Res. 2010, 1343, 178–185. [Google Scholar] [CrossRef]
- Ren, L.; Zhan, P.; Wang, Q.; Wang, C.; Liu, Y.; Yu, Z.; Zhang, S. Curcumin upregulates Nrf-2 system by repressing inflammatory signaling-mediated Keap1 expression in insulin-resistant conditions. Biochem. Biophys. Res. Commun. 2019, 514, 691–698. [Google Scholar] [CrossRef]
- Kubo, E.; Chhunchha, B.; Singh, P.; Sasaki, H.; Singh, D.P. Sulforaphane reactivates cellular antioxidant defense by unducing Nrf-22/ARE/Prdx6 activity over aging and oxidative stress. Sci. Rep. 2017, 7, 14120. [Google Scholar] [CrossRef] [Green Version]
- Pugh, N.; Ross, S.A.; ElSohly, H.N.; ElSohly, M.A.; Pasco, D.S. Isolation of three high molecular weigth polysaccharide preparations with potent immunostimulatory activity from Spirulina platensis in Aphanizomen flos-aquae and Chlorella pyronoidosa. Planta Med. 2001, 67, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Grzanna, R.; Polotsky, A.; Phan, P.V.; Pugh, N.; Pasco, D.; Frondoza, C.G. Immolina, a high-molecular-weigth polysaccharide fraction of spirulina, enhances chemokine expression in human monocytic thp-1 cells. J. Altern. Complement. Med. 2006, 12, 429–435. [Google Scholar] [CrossRef]
- Merino, J.J.; Belver-Landette, V.; Oset-Gasque, M.J.; Cubelos, B. CXCR4/CXCR7 molecular involvement in neuronal and neural progenitor migration: Focus in CNS repair. J. Cell Physiol. 2015, 230, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Lapidot, T.; Petit, I. Current understanding of stem cells mobilization: The roles of chemokines, proteolytic enzymes, adhesion molecules, cytokines and stromal cells. Exp. Hematol. 2002, 30, 14153–14160. [Google Scholar]
- Larochelle, A.; Krouse, A.; Metger, M.; Orlic, D.; Donahue, R.E.; Ficker, S.; Bridger, G.; Dunbar, C.E.; Hematti, P. AMD3100 mobilizates hematopoietic stem cells with long-term repopulating capacity in nonhuman primates. Blood 2006, 107, 3773–3778. [Google Scholar] [CrossRef] [Green Version]
- Son, B.; Marquez-Curtis, L.A.; Kucia, M.; Wysoczynski, M.; Turner, A.R.; Ratajczak, M.Z. Migration of bone marrow and cord blood mesenchymal stem cells in vitro is regulated by stromal-derived factor-1-CXCR4 and hepatocyte growth factor-c-met axis and involves matrix metalloproteinases. Stem Cells 2006, 24, 1254–1264. [Google Scholar] [CrossRef]
- Jensen, G.S.; Hart, A.N.; Zaske, L.A.M.; Drapeau, C.; Gupta, N.; Schaeffer, D.J.; Cruickshank, J.A. Mobilization of human CD34+ CD 133+ and CD34+ CD133– stem cells in vivo by consumption of an extract from Aphanizomenon flos-aquae-related to modulation of CXCR4 expression by an L-selectin ligand? Cardiovasc. Revasc. Med. 2007, 8, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Van Zante, A.; Rosen, S.D. Sulphated endothelial ligands for L-selectin in lymphocyte homing and inflammation. Biochem. Soc. Trans. 2003, 31, 313–317. [Google Scholar] [CrossRef]
- Shytle, D.R.; Tan, J.; Ehrhart, J.; Smith, A.J.; Sanberg, C.D.; Sanberg, P.R.; Anderson, J.; Bickford, P.C. Effects of blue-green algae extracts on the proliferation of human adult stem cells in vitro: A preliminary study. Med. Sci. Monit. 2010, 16, BR1–BR5. [Google Scholar]
- Sarifi, S.; Vahed, S.Z.; Ahmadian, E.; Dizaj, S.M.; Abedi, A.; Khatibi, S.M.H.; Samiel, M. Stem cell teraphy: Curcumin does the trick. Phytother. Res. 2019, 38, 2927–2937. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Chang, C.L.; Lee, H.-C.; Chi, C.; Pan, J.P.; Yang, W.C. Curcumin induces the apoptosis of monocytic leukemia THP-1 cells via activation of JNK/ERK pathways. Complement. Altern. Med. 2012, 12, 22. [Google Scholar] [CrossRef] [Green Version]
- Shekhani, M.; Jayanthy, A.S.; Maddodi, N.; Setaburi, V. Cancer stem cells and tumor transdifferentiation for novel therapeutic strategies. Am. J. Stem Cells 2013, 2, 52–61. [Google Scholar]
- Shytle, R.D.; Ehrhart, J.; Tan, J.; Vila, J.; Cole, M.; Sanberg, C.D.; Sanberg, P.R.; Bickford, P.C. Oxidative stress of neural, hematopoietic, and stem cells: Protection by natural compounds. Rejuvemation Res. 2007, 10, 173–178. [Google Scholar] [CrossRef]
- Rocha, V.; Gamier, F.; Ionescu, I.; Gluckman, E. Hematopoioetic stem-cell transplantation using umbilical-cord blood cells. Rev. Investig. Clin. 2005, 57, 314–323. [Google Scholar]
- Wilkinson, A.C.; Yamazaki, S. The hematopoietic stem cell diet. Prog. Hematol. 2018, 107, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Bickford, P.C.; Tan, J.; Shytle, R.D.; Sanberg, C.D.; El-Badri, N.; Sanberg, P.R. Nutraceuticals synergically promote proliferation of human stem cells. Stem Cells Dev. 2004, 3, 50–58. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD45 | Concentration 12.5 (μg/mL) |
|---|---|
| CD34 | 10 |
| 7-AAD | 45–65 |
| Study Group | Total Percentage of CD34+ Cells That Are also CD45+ Cells |
|---|---|
| Placebo (n = 7) | 0.046 ± 0.049 |
| Control (liquid, n = 5) | 0.046 ± 0.051 |
| Control (powder, n = 5) | 0.056 ± 0.0068 |
| Cur 48 h (powder, n = 5) | 0.066 ± 0.0068 * |
| Cur 48 h (liquid, n = 5) | 0.068 ± 0.01 |
| Cur 7 days (n = 5) | 0.048 ± 0.0031 |
| Cur 38 days (n = 5) | 0.066 ± 0.0068 |
| Cont (Algae, n = 5) | 0.053 ± 0.043 |
| AFA (48 h, n = 5) | 0.1 ± 0.0005 * # * p < 0.05 vs. control (algae) |
| * p < 0.05 vs. control (powder) # p < 0.05 vs. placebo |
| Study Group | TNC (×108) | WBC (×106/mL) |
|---|---|---|
| Placebo (n = 7) | 5.32 ± 0.98 | 3.08 ± 0.37 |
| Control (powder n = 5) | 5.8 ± 0.35 | 3.3 ± 0.3 |
| Control (liquid, n = 5) | 6.2 ± 0.75 | 3.08 ± 0.17 |
| Cur 48 h (powder, n = 5) | 6.21 ± 0.3 | 2.9 ± 0.43 |
| Cur 48 h (liquid, n = 5) | 6.32 ± 0.28 | 2.54 ± 0.43 |
| Cur 7 days (n = 5) | 5.52 ± 0.29 | 3.07 ± 0.17 |
| Cur 38 days (n = 5) | 5.89 ± 0.19 | 3.16 ± 0.35 |
| Cont (Algae, n = 5) | 5.7 ± 0.42 | 3.46 ± 0.13 |
| AFA (48 h, n = 5) | 6.21 ± 0.3 | 2.96 ± 0.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merino, J.J.; Cabaña-Muñoz, M.E.; Pelaz, M.J. The Bluegreen Algae (AFA) Consumption over 48 h Increases the Total Number of Peripheral CD34+ Cells in Healthy Patients: Effect of Short-Term and Long-Term Nutritional Supplementation (Curcumin/AFA) on CD34+ Levels (Blood). J. Pers. Med. 2020, 10, 49. https://doi.org/10.3390/jpm10020049
Merino JJ, Cabaña-Muñoz ME, Pelaz MJ. The Bluegreen Algae (AFA) Consumption over 48 h Increases the Total Number of Peripheral CD34+ Cells in Healthy Patients: Effect of Short-Term and Long-Term Nutritional Supplementation (Curcumin/AFA) on CD34+ Levels (Blood). Journal of Personalized Medicine. 2020; 10(2):49. https://doi.org/10.3390/jpm10020049
Chicago/Turabian StyleMerino, José Joaquín, María Eugenia Cabaña-Muñoz, and María Jesús Pelaz. 2020. "The Bluegreen Algae (AFA) Consumption over 48 h Increases the Total Number of Peripheral CD34+ Cells in Healthy Patients: Effect of Short-Term and Long-Term Nutritional Supplementation (Curcumin/AFA) on CD34+ Levels (Blood)" Journal of Personalized Medicine 10, no. 2: 49. https://doi.org/10.3390/jpm10020049