
Serum Phosphorus, Serum Bicarbonate, and Renal Function in Relation to Liver CYP1A2 Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Assessment of General Characteristics through Self Reports
2.3. General Body Measurements
2.4. Caffeine Intake Assessment
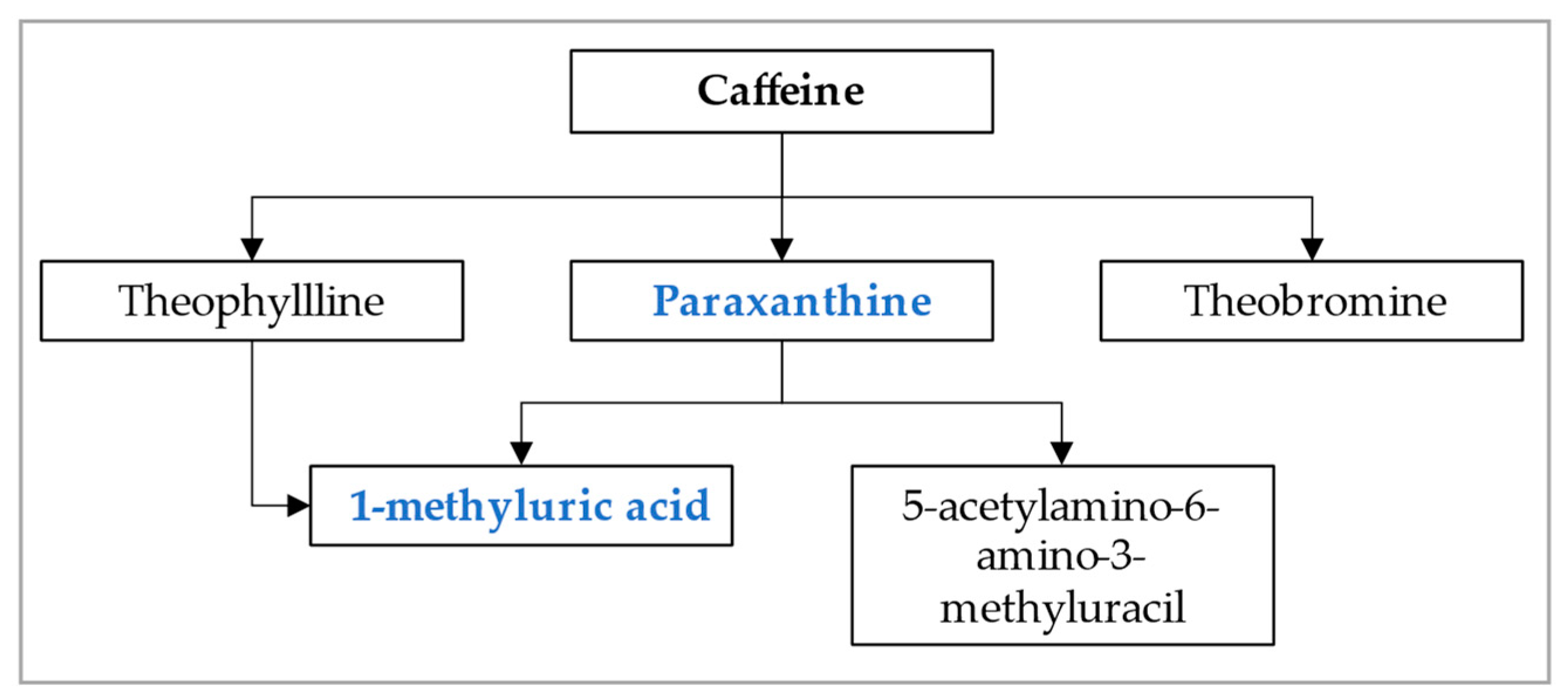
2.5. Assessment of Urinary Caffeine Metabolite and CYP1A2 Activity
2.6. Serum Bicarbonate and Serum Phosphate Assessment
2.7. Estimated Glomerular Filtration Rate (eGFR)
2.8. Hepatic Steatosis Index
2.9. Statistical Analyses
3. Results
3.1. Demographic Characteristics in the Whole Data Set (Table 1) and by HSI Status (Table 2)

{kind=link}
{kind=link}
{kind=link}
| Characteristic | Total Cohort (n = 1381) |
|---|---|
| Gender | |
| Female | 707 (51%) |
| Male | 674 (49%) |
| Age | |
| 18 to 39 | 458 (33%) |
| 40 to 59 | 493 (36%) |
| 60 or older | 430 (31%) |
| Ethnicity | |
| Mexican American | 246 (18%) |
| Other Hispanic | 143 (10%) |
| White Non-Hispanic | 732 (53%) |
| Black Non-Hispanic | 205 (15%) |
| Other | 55 (4%) |
| Smoking intensity, total cigarettes over lifetime | |
| 0 | 688 (53%) |
| 1 to 50,400 | 252 (19%) |
| >50,400 | 364 (28%) |
| BMI, kg/m² | |
| <25 | 394 (29%) |
| 25 to 29.9 | 460 (33%) |
| >29.9 | 527 (38%) |
| Blood pressure status | |
| Normotensive | 659 (48%) |
| Elevated | 255 (18%) |
| Hypertension 1 | 246 (18%) |
| Hypertension 2 | 221 (16%) |
| Diabetes | |
| Yes | 133 (10%) |
| No | 1212 (88%) |
| Borderline | 33 (2%) |
| eGFR, mL/min/1.73 cm² | |
| >90 | 902 (65%) |
| 60 to 90 | 395 (29%) |
| <60 | 84 (6%) |
| Physical activity, MET-hours/week | |
| 0 | 373 (27%) |
| >0 to 4 | 93 (7%) |
| >4 to 40 | 496 (36%) |
| >40 | 419 (30%) |
| Paraxanthine/caffeine ratio | 0.15 (0.07, 0.30) |
| 1-methyluric acid/caffeine ratio | 0.59 (0.33, 1.07) |
| Bicarbonate level, mmol/L | 26 (24, 27) |
| Phosphorus level, mmol/L | 26 (24, 27) |
| Characteristic | HSI < 30 (n = 197) | 30 ≤ HSI < 36 (n = 393) | 36 ≤ HSI < 42 (n = 432) | HSI ≥ 42 (n = 359) |
|---|---|---|---|---|
| Gender | ||||
| Female | 104 (53%) | 179 (46%) | 215 (50%) | 209 (58%) |
| Male | 93 (47%) | 214 (54%) | 217 (50%) | 150 (42%) |
| Age | ||||
| 18 to 39 | 88 (45%) | 141 (36%) | 116 (27%) | 113 (31%) |
| 40 to 59 | 54 (27%) | 127 (32%) | 154 (36%) | 158 (44%) |
| 60 or older | 55 (28%) | 125 (32%) | 162 (37%) | 88 (25%) |
| Ethnicity | ||||
| Mexican American | 16 (8%) | 64 (16%) | 87 (20%) | 79 (22%) |
| Other Hispanic | 20 (10%) | 40 (10%) | 45 (10%) | 38 (11%) |
| White Non-Hispanic | 113 (57%) | 220 (56%) | 228 (53%) | 171 (48%) |
| Black Non-Hispanic | 29 (15%) | 50 (13%) | 63 (15%) | 63 (18%) |
| Other | 19 (10%) | 19 (5%) | 9 (2%) | 8 (2%) |
| Smoking intensity, total cigarettes over lifetime | ||||
| 0 | 89 (50%) | 189 (51%) | 220 (54%) | 190 (55%) |
| 1 to 50,400 | 40 (23%) | 79 (22%) | 72 (17%) | 61 (17%) |
| >50,400 | 48 (27%) | 101 (27%) | 119 (29%) | 96 (28%) |
| BMI, kg/m² | ||||
| <25 | 195 (98%) | 187 (47%) | 12 (3%) | 0 (0%) |
| 25 to 29.9 | 1 (1%) | 199 (51%) | 252 (58%) | 8 (2%) |
| >29.9 | 1 (1%) | 7 (2%) | 168 (39%) | 351 (98%) |
| Blood pressure status | ||||
| Normotensive | 124 (63%) | 204 (52%) | 186 (43%) | 145 (40%) |
| Elevated | 18 (9%) | 84 (21%) | 88 (20%) | 65 (18%) |
| Hypertension 1 | 30 (15%) | 58 (15%) | 69 (16%) | 89 (25%) |
| Hypertension 2 | 25 (13%) | 47 (12%) | 89 (21%) | 60 (17%) |
| Diabetes | ||||
| Yes | 3 (2%) | 18 (5%) | 54 (12%) | 58 (16%) |
| No | 191 (98%) | 371 (94%) | 370 (86%) | 280 (78%) |
| Borderline | 0 (0%) | 4 (1%) | 8 (2%) | 21 (6%) |
| eGFR, mL/min/1.73 cm² | ||||
| >90 | 125 (63%) | 252 (64%) | 274 (63%) | 251 (70%) |
| 60 to 90 | 64 (32%) | 115 (29%) | 120 (28%) | 96 (27%) |
| <60 | 8 (4%) | 26 (7%) | 38 (9%) | 12 (3%) |
| Physical activity, MET-hours/week | ||||
| 0 | 39 (20%) | 83 (21%) | 128 (30%) | 123 (34%) |
| >0 to 4 | 16 (8%) | 25 (6%) | 28 (6%) | 24 (7%) |
| >4 to 40 | 77 (39%) | 155 (39%) | 150 (35%) | 114 (32%) |
| >40 | 65 (33%) | 130 (33%) | 126 (29%) | 98 (27%) |
| Paraxanthine/caffeine ratio | 0.16 (0.06, 0.32) | 0.15 (0.07, 0.31) | 0.15 (0.07, 0.30) | 0.15 (0.07, 0.28) |
| 1-Methyluric acid/caffeine ratio | 0.66 (0.40, 1.14) | 0.58 (0.33, 1.07) | 0.57 (0.32, 1.06) | 0.55 (0.32, 0.99) |
| Bicarbonate level, mmol/L | 26 (25, 27) | 26 (25, 27) | 25 (24, 27) | 25 (24, 27) |
| Phosphorus level, mmol/L | 1.20 (1.10, 1.32) | 1.23 (1.10, 1.32) | 1.20 (1.07, 1.32) | 1.16 (1.07, 1.29) |
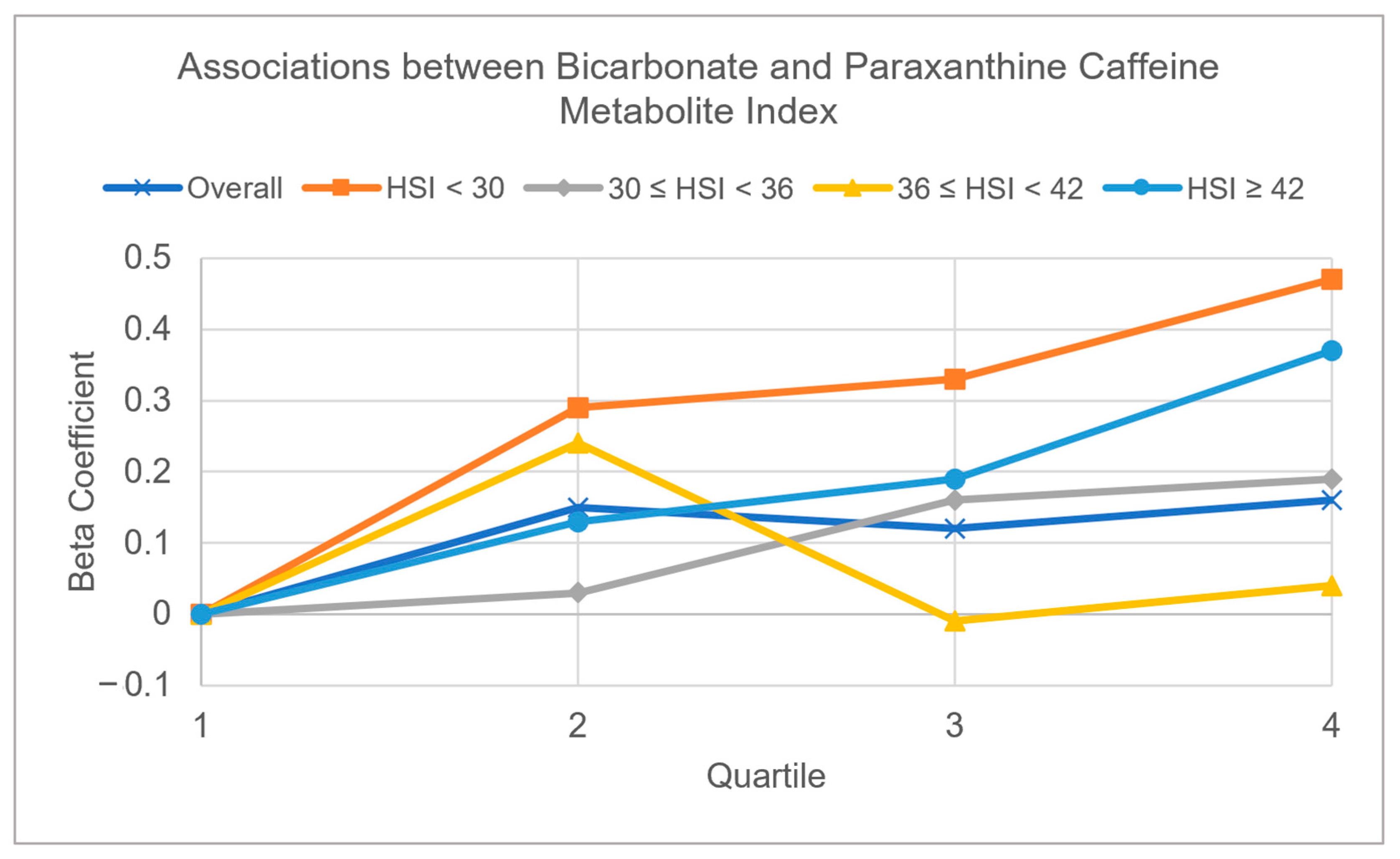
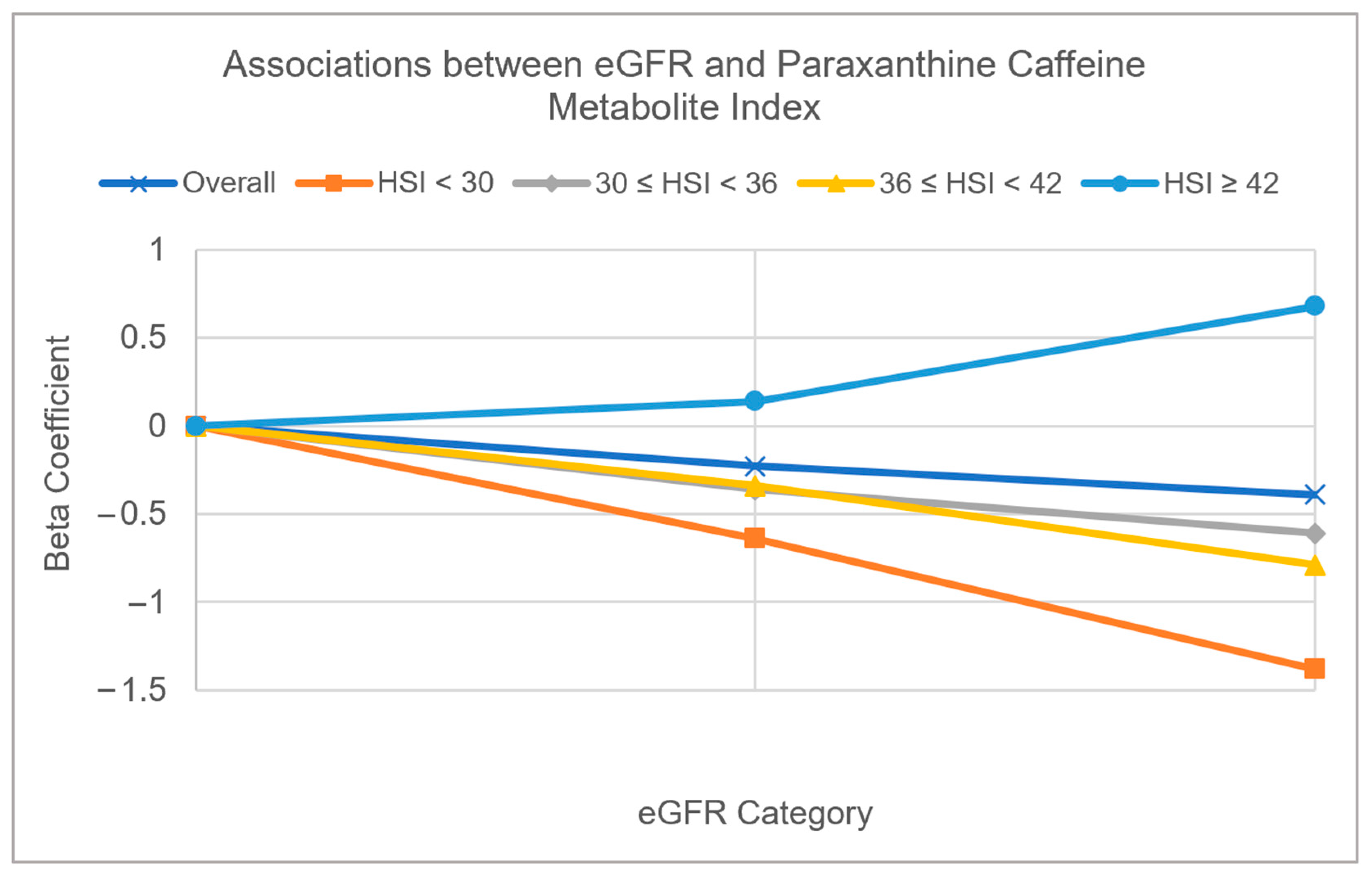
3.2. Multivariable Adjusted Associations of Serum Bicarbonate, Serum Phosphorus, and eGFR with CYP1A2 Activity Measured by Caffeine Metabolite Indices (Table 3 and Figure 2 and Figure 3)
| Overall (n = 1381) | HSI < 30 (n = 197) | 30 ≤ HSI < 36 (n = 393) | 36 ≤ HSI < 42 (n = 432) | HSI ≥ 42 (n = 359) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Paraxanthine | 1-MUA | Paraxanthine | 1-MUA | Paraxanthine | 1-MUA | Paraxanthine | 1-MUA | Paraxanthine | 1-MUA | |
| β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | β (p-Value) | |
| Phosphorus, mmol/L | ||||||||||
| Quartile 1 (0.55–1.10) | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref |
| Quartile 2 (1.13–1.20) | −0.01 (0.94) | −0.02 (0.69) | 0.07 (0.80) | −0.09 (0.65) | 0.01 (0.96) | −0.02 (0.90) | −0.19 (0.26) | −0.15 (0.25) | 0.13 (0.58) | 0.11 (0.47) |
| Quartile 3 (1.22–1.36) | −0.04 (0.58) | 0.03 (0.61) | −0.19 (0.51) | 0.07 (0.69) | 0.04 (0.75) | −0.01 (0.94) | −0.03 (0.87) | 0.08 (0.46) | −0.11 (0.48) | 0.07 (0.54) |
| Quartile 4 (1.38–2.62) | 0.13 (0.15) | 0.11 (0.22) | 0.05 (0.90) | 0.01 (0.96) | 0.29 (0.10) | 0.25 (0.04) | 0.05 (0.82) | 0.11 (0.51) | 0.14 (0.43) | 0.03 (0.87) |
| Bicarbonate, mmol/L | ||||||||||
| Quartile 1 (15–24) | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref |
| Quartile 2 (25–25) | 0.15 (0.21) | 0.01 (0.88) | 0.29 (0.24) | 0.06 (0.72) | 0.03 (0.92) | −0.05 (0.74) | 0.24 (0.20) | 0.20 (0.14) | 0.13 (0.58) | −0.12 (0.43) |
| Quartile 3 (26–26) | 0.12 (0.24) | 0.12 (0.10) | 0.33 (0.25) | 0.12 (0.65) | 0.16 (0.31) | 0.15 (0.20) | −0.01 (0.96) | 0.08 (0.59) | 0.19 (0.36) | 0.17 (0.24) |
| Quartile 4 (27–38) | 0.16 (0.06) | 0.13 (0.12) | 0.47 (0.19) | 0.12 (0.67) | 0.19 (0.28) | 0.23 (0.19) | 0.04 (0.82) | 0.14 (0.33) | 0.37 (0.09) | 0.15 (0.34) |
| eGFR | ||||||||||
| >90 | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref |
| 60 to 90 | −0.23 (0.0031) | −0.11 (0.08) | −0.64 (0.06) | −0.52 (0.03) | −0.36 (0.10) | −0.16 (0.24) | −0.34 (0.04) | −0.19 (0.28) | 0.14 (0.32) | 0.04 (0.78) |
| <60 | −0.39 (0.0010) | 0.25 (0.02) | −1.38 (0.04) | −0.62 (0.21) | −0.61 (0.03) | 0.15 (0.49) | −0.79 (0.0018) | 0.27 (0.14) | 0.68 (0.11) | 0.52 (0.23) |


4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ando, Y.; Jou, J.H. Nonalcoholic Fatty Liver Disease and Recent Guideline Updates. Clin. Liver Dis. 2021, 17, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.-F.; Schattenberg, J.M.; et al. A New Definition for Metabolic Dysfunction-Associated Fatty Liver Disease: An International Expert Consensus Statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Estes, C.; Anstee, Q.M.; Arias-Loste, M.T.; Bantel, H.; Bellentani, S.; Caballeria, J.; Colombo, M.; Craxi, A.; Crespo, J.; Day, C.P.; et al. Modeling NAFLD Disease Burden in China, France, Germany, Italy, Japan, Spain, United Kingdom, and United States for the Period 2016–2030. J. Hepatol. 2018, 69, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Baumgartner, K.; Bositis, C. Cirrhosis: Diagnosis and Management. Am. Fam. Physician 2019, 100, 759–770. [Google Scholar]
- Piazzolla, V.A.; Mangia, A. Noninvasive Diagnosis of NAFLD and NASH. Cells 2020, 9, 1005. [Google Scholar] [CrossRef]
- Baumgartner, K.; Cooper, J.; Smith, A.; St Louis, J. Liver Disease: Cirrhosis. FP Essent. 2021, 511, 36–43. [Google Scholar]
- Kisseleva, T.; Brenner, D. Molecular and Cellular Mechanisms of Liver Fibrosis and Its Regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Kalra, R.; Patel, N.; Arora, P.; Arora, G. Cardiovascular Health and Disease Among Asian-Americans (from the National Health and Nutrition Examination Survey). Am. J. Cardiol. 2019, 124, 270–277. [Google Scholar] [CrossRef]
- Ambühl, P.M. Protein Intake in Renal and Hepatic Disease. Int. J. Vitam. Nutr. Res. Int. Z. Vitam. Ernahrungsforschung J. Int. Vitaminol. Nutr. 2011, 81, 162–172. [Google Scholar] [CrossRef]
- Hakkola, J.; Hukkanen, J.; Turpeinen, M.; Pelkonen, O. Inhibition and Induction of CYP Enzymes in Humans: An Update. Arch. Toxicol. 2020, 94, 3671–3722. [Google Scholar] [CrossRef]
- Wijnen, P.A.H.M.; Op Den Buijsch, R.A.M.; Drent, M.; Kuipers, P.M.J.C.; Neef, C.; Bast, A.; Bekers, O.; Koek, G.H. Review Article: The Prevalence and Clinical Relevance of Cytochrome P450 Polymorphisms. Aliment. Pharmacol. Ther. 2007, 26, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhu, X.; Badawy, S.; Ihsan, A.; Liu, Z.; Xie, C.; Wang, X. Metabolism and Mechanism of Human Cytochrome P450 Enzyme 1A2. Curr. Drug Metab. 2021, 22, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Gonzalez, F.J.; Kalow, W.; Tang, B.K. Biotransformation of Caffeine, Paraxanthine, Theobromine and Theophylline by cDNA-Expressed Human CYP1A2 and CYP2E1. Pharmacogenetics 1992, 2, 73–77. [Google Scholar] [CrossRef]
- Garduno, A.; Wu, T. Tobacco Smoke and CYP1A2 Activity in a US Population with Normal Liver Enzyme Levels. Int. J. Environ. Res. Public. Health 2021, 18, 2225. [Google Scholar] [CrossRef]
- Hopkins, E.; Sanvictores, T.; Sharma, S. Physiology, Acid Base Balance. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Katopodis, P.; Pappas, E.M.; Katopodis, K.P. Acid-Base Abnormalities and Liver Dysfunction. Ann. Hepatol. 2022, 27, 100675. [Google Scholar] [CrossRef] [PubMed]
- Sacks, G.S. The ABC’s of Acid-Base Balance. J. Pediatr. Pharmacol. Ther. 2004, 9, 235–242. [Google Scholar] [CrossRef]
- Scheiner, B.; Lindner, G.; Reiberger, T.; Schneeweiss, B.; Trauner, M.; Zauner, C.; Funk, G.-C. Acid-Base Disorders in Liver Disease. J. Hepatol. 2017, 67, 1062–1073. [Google Scholar] [CrossRef]
- Seifter, J.L.; Chang, H.-Y. Disorders of Acid-Base Balance: New Perspectives. Kidney Dis. 2016, 2, 170–186. [Google Scholar] [CrossRef]
- NHANES. NHANES 2009–2010: Lab Manual. Available online: https://wwwn.cdc.gov/nchs/data/nhanes/2009-2010/manuals/lab.pdf (accessed on 7 December 2022).
- NHANES. NHANES 2009-2010: Physical Activity—Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/nchs/nhanes/2009-2010/PAQ_F.htm (accessed on 15 November 2022).
- NHANES. NHANES 2009–2010: Smoking—Cigarette Use Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/nchs/nhanes/2009-2010/smq_f.htm (accessed on 4 March 2023).
- NHANES. NHANES 2009–2010: Blood Pressure Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/nchs/nhanes/2009-2010/BPX_F.htm (accessed on 4 March 2023).
- NHANES. NHANES 2009–2010: Body Measures Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/nchs/nhanes/2009-2010/BMX_F.htm (accessed on 4 March 2023).
- American Heart Association. High Blood Pressure|American Heart Association. Available online: https://www.heart.org/en/health-topics/high-blood-pressure (accessed on 4 March 2023).
- NHANES. NHANES 2009–2010: Dietary Interview—Individual Foods, First Day Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/Nchs/Nhanes/2009-2010/DR1IFF_F.htm (accessed on 4 March 2023).
- Nehlig, A. Interindividual Differences in Caffeine Metabolism and Factors Driving Caffeine Consumption|Pharmacological Reviews. Pharmacol. Rev. 2018, 70, 384–411. [Google Scholar] [CrossRef]
- NHANES. NHANES 2009–2010: Caffeine & Caffeine Metabolites—Urine Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/Nchs/Nhanes/2009-2010/CAFE_F.htm (accessed on 15 November 2022).
- NHANES. NHANES 2009–2010: Standard Biochemistry Profile Data Documentation, Codebook, and Frequencies. Available online: https://wwwn.cdc.gov/Nchs/Nhanes/2009-2010/BIOPRO_F.htm#Description_of_Laboratory_Methodology (accessed on 15 November 2022).
- Giglio, D. A New Equation for Estimating Glomerular Filtration Rate in Cancer Patients. Chemotherapy 2014, 60, 63–72. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, D.; Kim, H.J.; Lee, C.-H.; Yang, J.I.; Kim, W.; Kim, Y.J.; Yoon, J.-H.; Cho, S.-H.; Sung, M.-W.; et al. Hepatic Steatosis Index: A Simple Screening Tool Reflecting Nonalcoholic Fatty Liver Disease. Dig. Liver Dis. 2010, 42, 503–508. [Google Scholar] [CrossRef]
- Cohen, R.D. Roles of the liver and kidney in acid—Base regulation and its disorders. Br. J. Anaesth. 1991, 67, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Almajid, A.N.; Sugumar, K. Physiology, Bile. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Hundt, M.; Basit, H.; John, S. Physiology, Bile Secretion. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Ishiguro, H.; Yamamoto, A.; Nakakuki, M.; Yi, L.; Ishiguro, M.; Yamaguchi, M.; Kondo, S.; Mochimaru, Y. Physiology and Pathophysiology of Bicarbonate Secretion by Pancreatic Duct Epithelium. Nagoya J. Med. Sci. 2012, 74, 1–18. [Google Scholar] [PubMed]
- Rajkumar, P.; Pluznick, J.L. Acid-Base Regulation in the Renal Proximal Tubules: Using Novel pH Sensors to Maintain Homeostasis. Am. J. Physiol. Ren. Physiol. 2018, 315, F1187–F1190. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Pérez-Felpete, N.; Fernández-Fernández, C.; Donapetry-García, C.; Pazos-García, C. Liver Glucose Metabolism in Humans. Biosci. Rep. 2016, 36, e00416. [Google Scholar] [CrossRef]
- Barmore, W.; Azad, F.; Stone, W.L. Physiology, Urea Cycle. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Ogobuiro, I.; Tuma, F. Physiology, Renal. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Mao, S.A.; Glorioso, J.M.; Nyberg, S.L. Liver Regeneration. Transl. Res. J. Lab. Clin. Med. 2014, 163, 352–362. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Tabibian, J.H.; Ekstedt, M.; Kechagias, S.; Hamaguchi, M.; Hultcrantz, R.; Hagström, H.; Yoon, S.K.; Charatcharoenwitthaya, P.; et al. Association of Non-Alcoholic Fatty Liver Disease with Chronic Kidney Disease: A Systematic Review and Meta-Analysis. PLoS Med. 2014, 11, e1001680. [Google Scholar] [CrossRef]
- Mantovani, A.; Zaza, G.; Byrne, C.D.; Lonardo, A.; Zoppini, G.; Bonora, E.; Targher, G. Nonalcoholic Fatty Liver Disease Increases Risk of Incident Chronic Kidney Disease: A Systematic Review and Meta-Analysis. Metabolism. 2018, 79, 64–76. [Google Scholar] [CrossRef]
- Umbro, I.; Baratta, F.; Angelico, F.; Del Ben, M. Nonalcoholic Fatty Liver Disease and the Kidney: A Review. Biomedicines 2021, 9, 1370. [Google Scholar] [CrossRef] [PubMed]
- Ryou, M.; Stylopoulos, N.; Baffy, G. Nonalcoholic Fatty Liver Disease and Portal Hypertension. Explor. Med. 2020, 1, 149–169. [Google Scholar] [CrossRef]
- Mazidi, M.; Mikhailidis, D.P.; Dehghan, A.; Jóźwiak, J.; Covic, A.; Sattar, N.; Banach, M. The Association between Coffee and Caffeine Consumption and Renal Function: Insight from Individual-Level Data, Mendelian Randomization, and Meta-Analysis. Arch. Med. Sci. AMS 2021, 18, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.C.; Schneider, J.A.; Tanguay, D.A.; Choi, J.; Acharya, T.; Stanley, S.E.; Jiang, R.; Messer, C.J.; Chew, A.; Han, J.H.; et al. Haplotype Variation and Linkage Disequilibrium in 313 Human Genes. Science 2001, 293, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Fekete, F.; Mangó, K.; Minus, A.; Tóth, K.; Monostory, K. CYP1A2 mRNA Expression Rather than Genetic Variants Indicate Hepatic CYP1A2 Activity. Pharmaceutics 2022, 14, 532. [Google Scholar] [CrossRef]
- Nelson, M.R.; Marnellos, G.; Kammerer, S.; Hoyal, C.R.; Shi, M.M.; Cantor, C.R.; Braun, A. Large-Scale Validation of Single Nucleotide Polymorphisms in Gene Regions. Genome Res. 2004, 14, 1664–1668. [Google Scholar] [CrossRef]
- Kall, M.A.; Vang, O.; Clausen, J. Effects of Dietary Broccoli on Human in Vivo Drug Metabolizing Enzymes: Evaluation of Caffeine, Oestrone and Chlorzoxazone Metabolism. Carcinogenesis 1996, 17, 793–799. [Google Scholar] [CrossRef]
- Toshikuni, N.; Tsutsumi, M.; Arisawa, T. Clinical Differences between Alcoholic Liver Disease and Nonalcoholic Fatty Liver Disease. World J. Gastroenterol. 2014, 20, 8393–8406. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, J.; Lemus, H.; Wu, T. Serum Phosphorus, Serum Bicarbonate, and Renal Function in Relation to Liver CYP1A2 Activity. Diagnostics 2023, 13, 2996. https://doi.org/10.3390/diagnostics13182996
Ito J, Lemus H, Wu T. Serum Phosphorus, Serum Bicarbonate, and Renal Function in Relation to Liver CYP1A2 Activity. Diagnostics. 2023; 13(18):2996. https://doi.org/10.3390/diagnostics13182996
Chicago/Turabian StyleIto, Joy, Hector Lemus, and Tianying Wu. 2023. "Serum Phosphorus, Serum Bicarbonate, and Renal Function in Relation to Liver CYP1A2 Activity" Diagnostics 13, no. 18: 2996. https://doi.org/10.3390/diagnostics13182996