
Multi-Omics Analyses to Identify FCGBP as a Potential Predictor in Head and Neck Squamous Cell Carcinoma


Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression Feature Analysis
2.2. Survival Prognosis Analysis
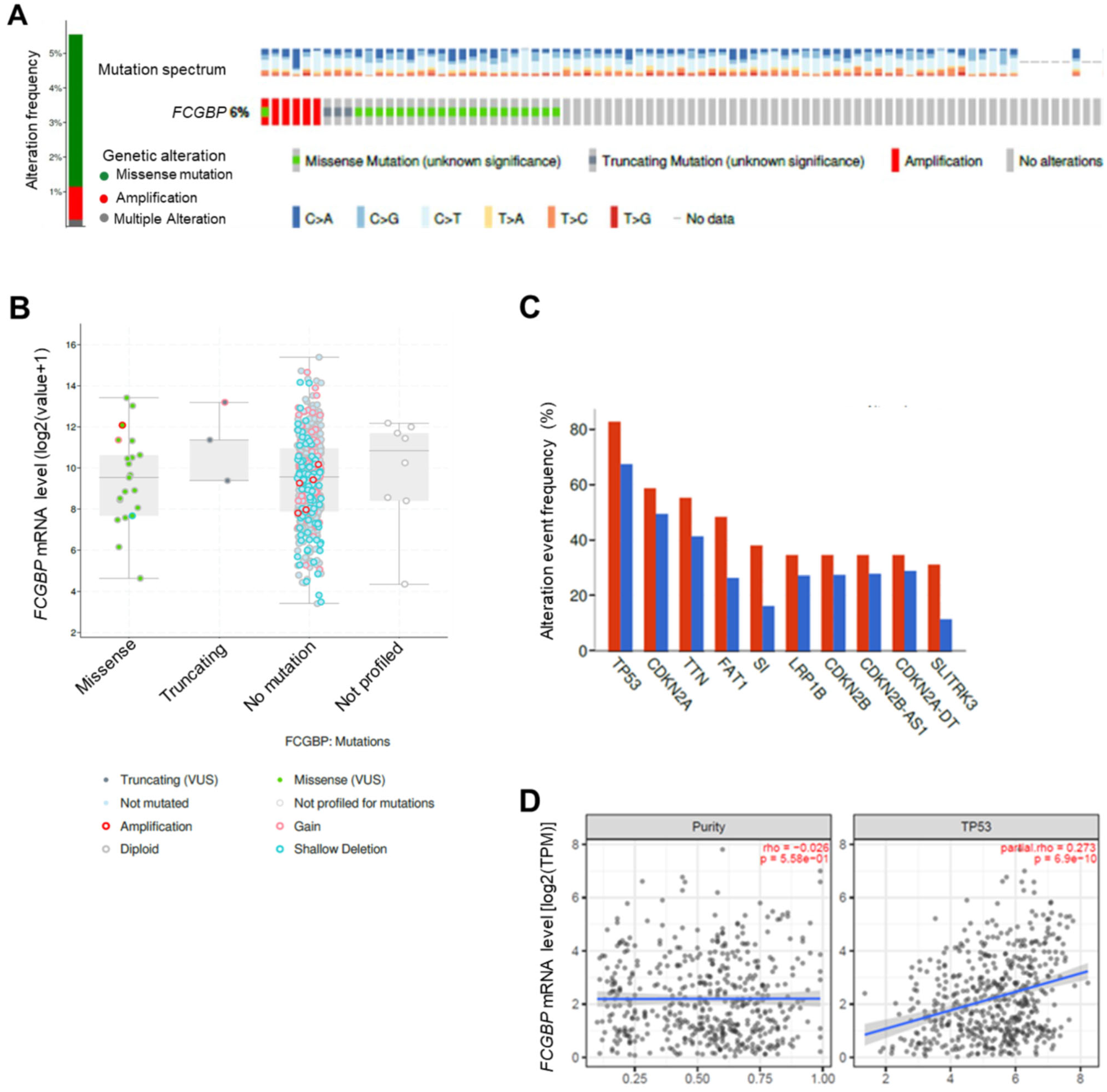
2.3. Gene Alteration Analysis
2.4. Molecular Interaction Analysis
2.5. Gene Set Enrichment Analysis
2.6. Immune Infiltration
3. Results
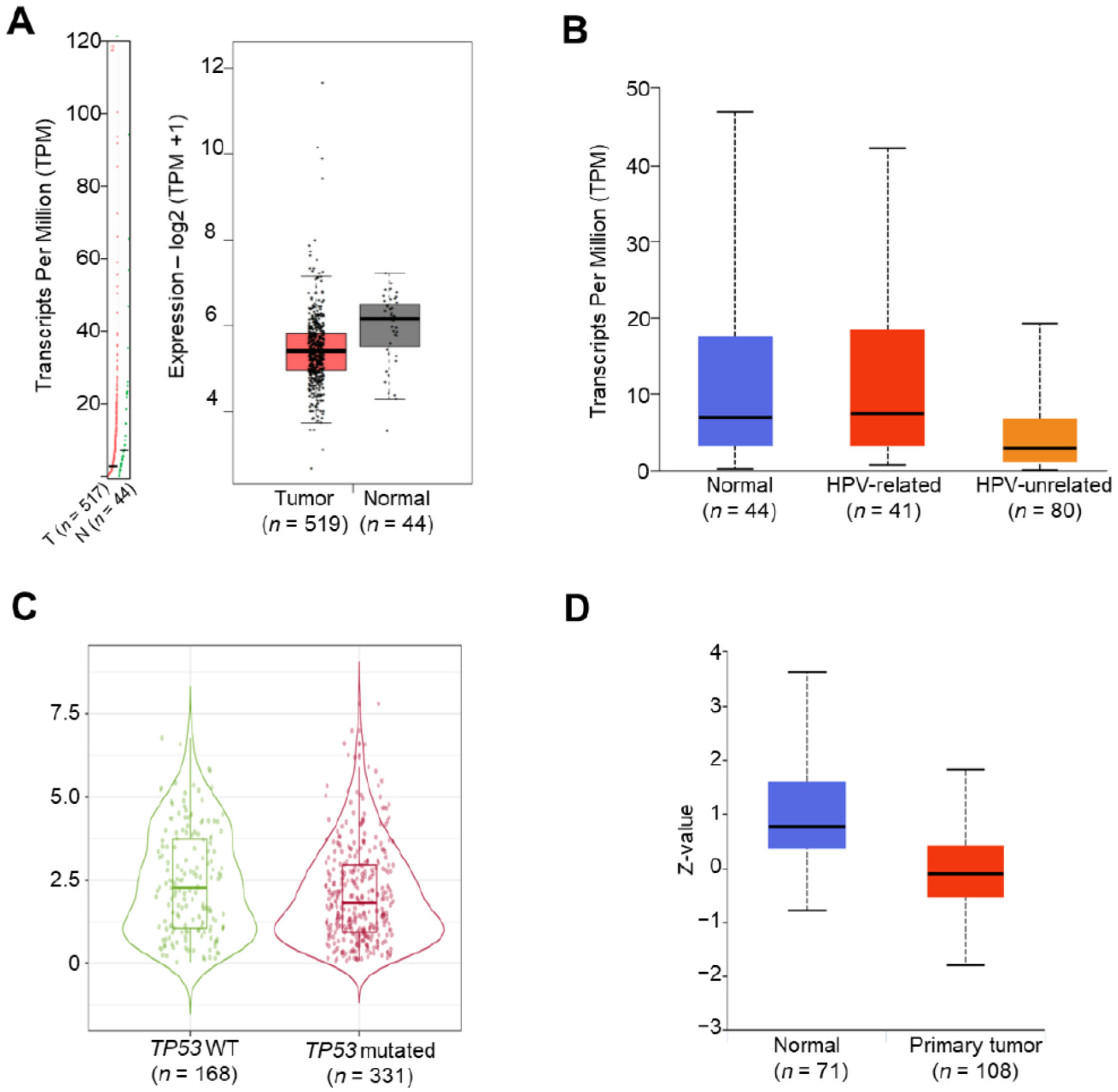
3.1. Downregulation of the FCGBP Level in Head and Neck Squamous Cell Carcinoma
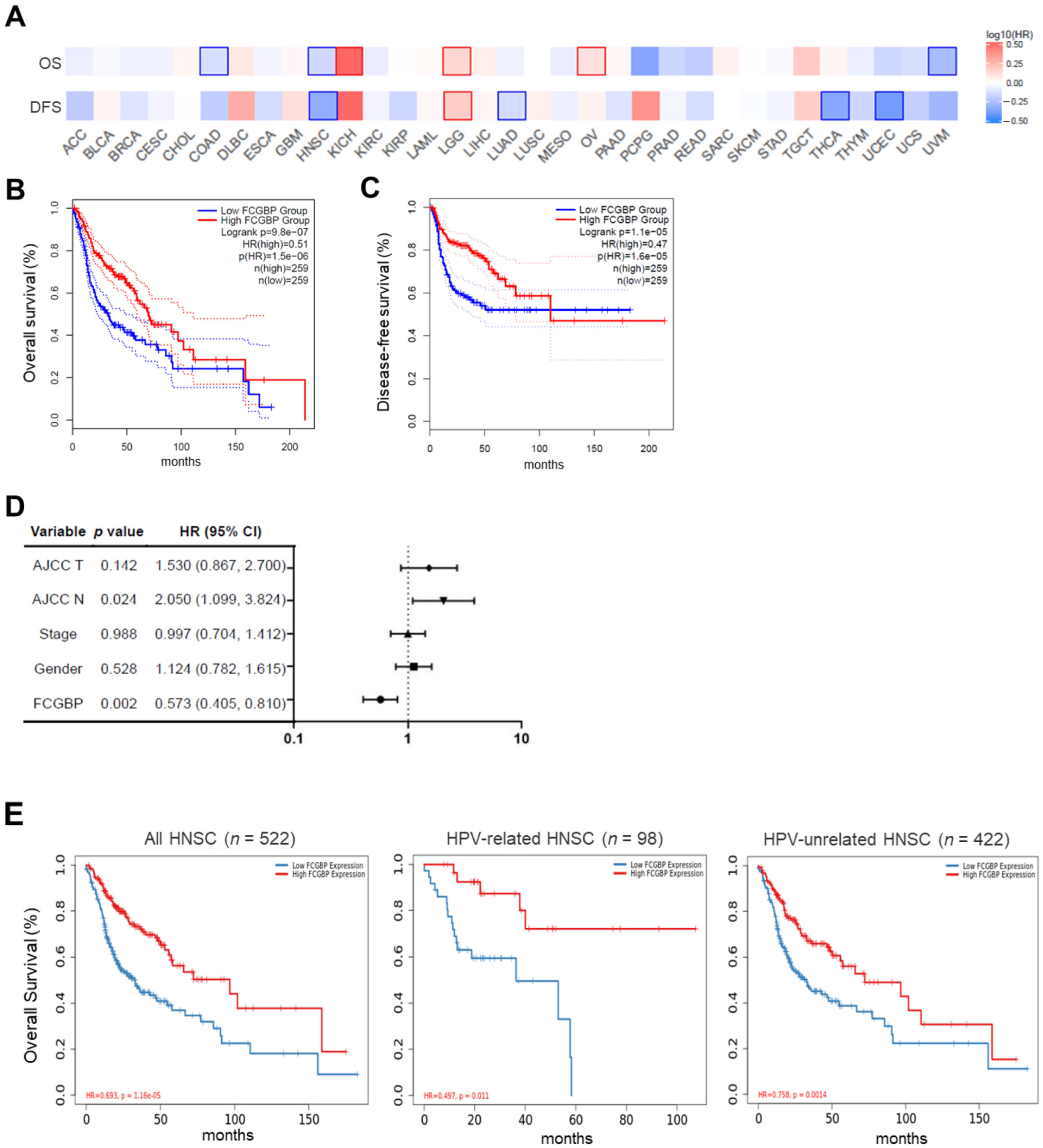
3.2. High FCGBP Level as an Independent Prognostic Factor for Favorable Survival in HNSC
3.3. The FCGBP mRNA Level Positively Correlates to TP53 WT in HNSC Patients
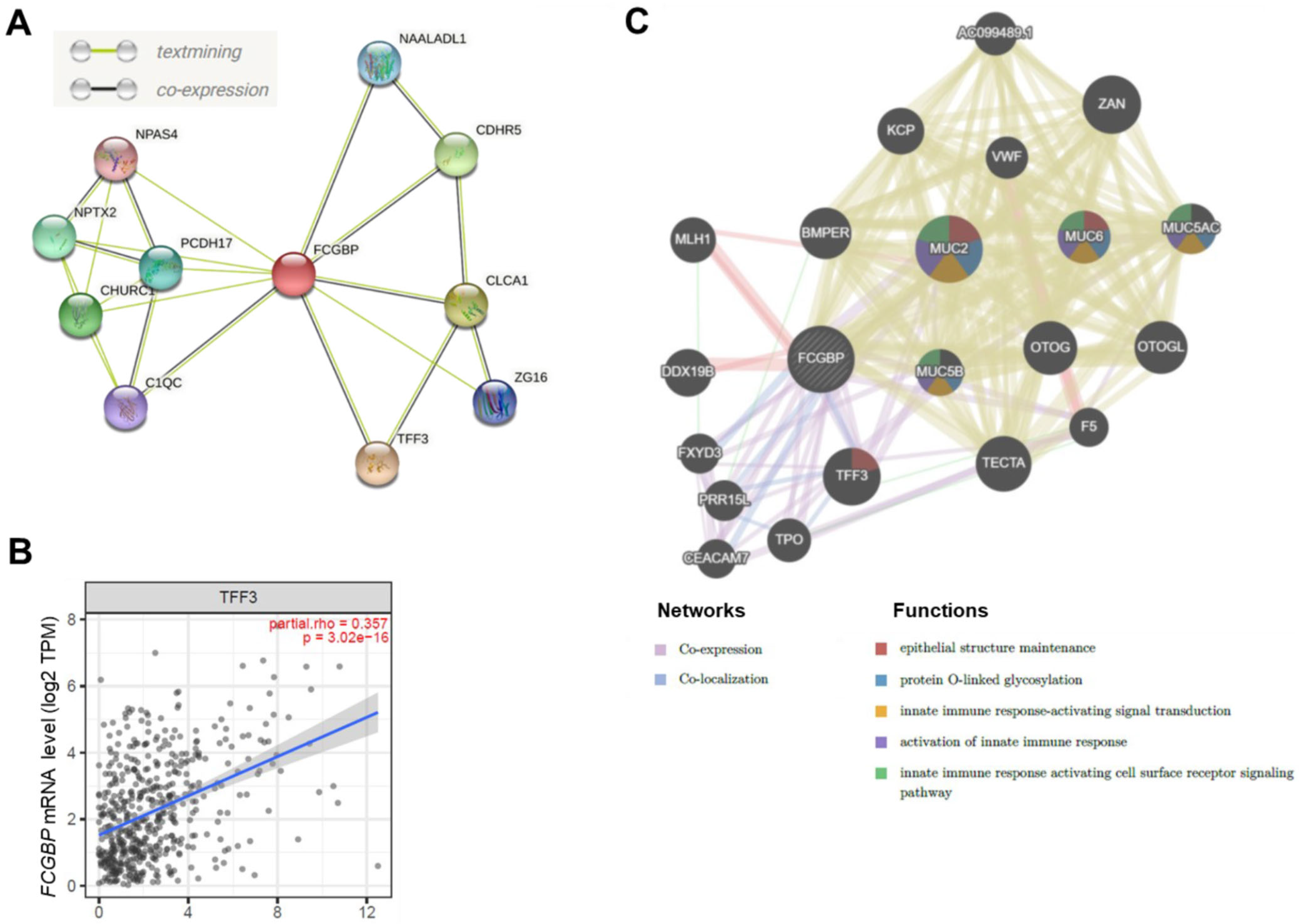
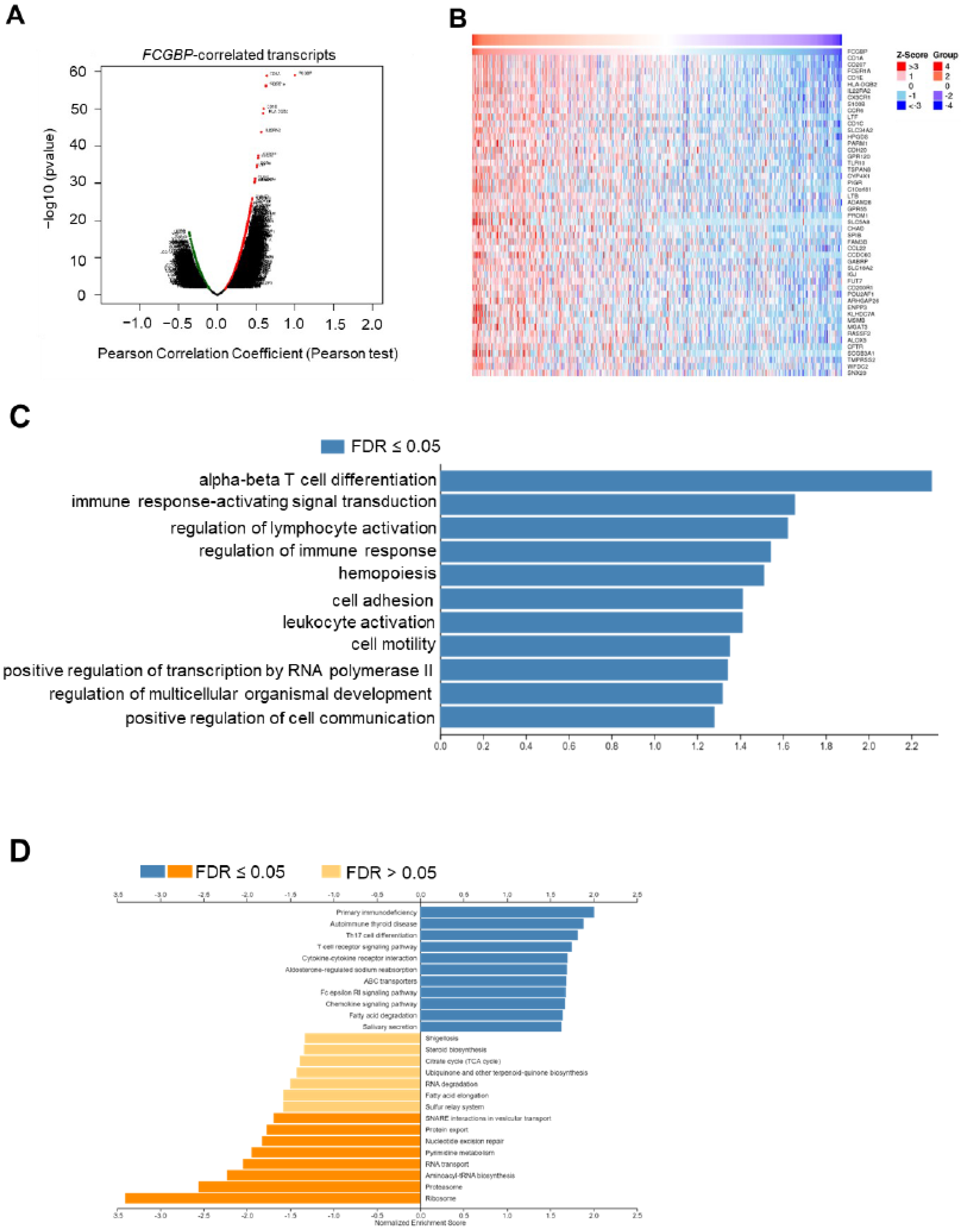
3.4. The FCGBP Correlated with Several Proteins in HNSC Patients
3.5. Functional Enrichment Analysis of FCGBP and Co-Expressed Genes in HNSC
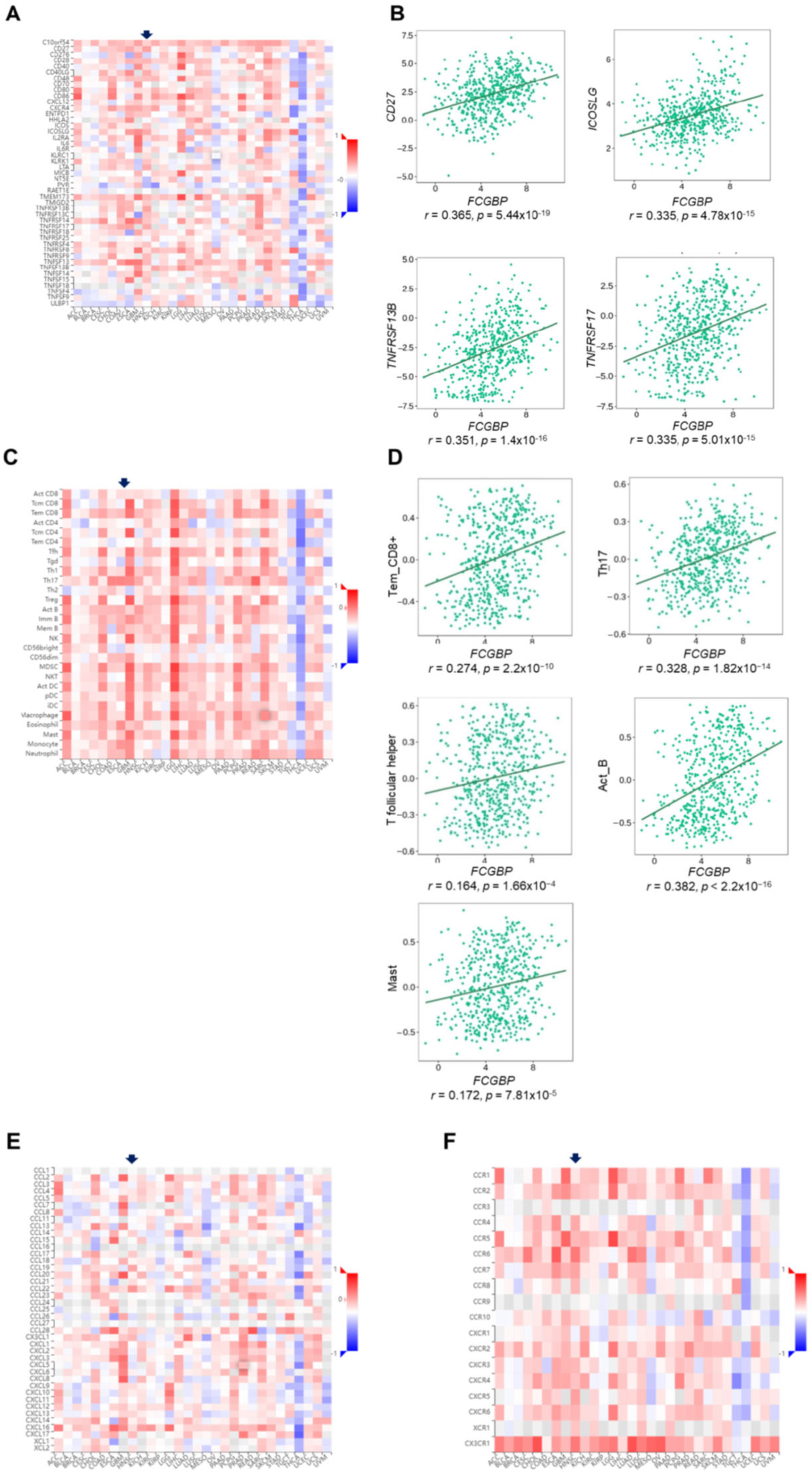
3.6. The FCGBP Level Correlates to Immunomodulators and Immune-Related Molecules, and the Abundance of Infiltration of Immune Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, G.; Westra, W.H.; Wang, S.J.; van Zante, A.; Wentz, A.; Kluz, N.; Rettig, E.; Ryan, W.R.; Ha, P.K.; Kang, H.; et al. Differences in the Prevalence of Human Papillomavirus (HPV) in Head and Neck Squamous Cell Cancers by Sex, Race, Anatomic Tumor Site, and HPV Detection Method. JAMA Oncol. 2017, 3, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Grulich, A.E.; Jin, F.; Conway, E.L.; Stein, A.N.; Hocking, J. Cancers attributable to human papillomavirus infection. Sex. Health 2010, 7, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Wiest, T.; Schwarz, E.; Enders, C.; Flechtenmacher, C.; Bosch, F.X. Involvement of intact HPV16 E6/E7 gene expression in head and neck cancers with unaltered p53 status and perturbed pRb cell cycle control. Oncogene 2002, 21, 1510–1517. [Google Scholar] [CrossRef] [Green Version]
- Rettig, E.M.; D’Souza, G. Epidemiology of head and neck cancer. Surg. Oncol. Clin. N. Am. 2015, 24, 379–396. [Google Scholar] [CrossRef]
- Kobayashi, K.; Ogata, H.; Morikawa, M.; Iijima, S.; Harada, N.; Yoshida, T.; Brown, W.R.; Inoue, N.; Hamada, Y.; Ishii, H.; et al. Distribution and partial characterisation of IgG Fc binding protein in various mucin producing cells and body fluids. Gut 2002, 51, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Denny, P.; Hagen, F.K.; Hardt, M.; Liao, L.; Yan, W.; Arellanno, M.; Bassilian, S.; Bedi, G.S.; Boontheung, P.; Cociorva, D.; et al. The proteomes of human parotid and submandibular/sublingual gland salivas collected as the ductal secretions. J. Proteome Res. 2008, 7, 1994–2006. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Wang, R.; Su, B.; Luo, Y.; Terhune, J.; Beck, B.; Peatman, E. Evasion of mucosal defenses during Aeromonas hydrophila infection of channel catfish (Ictalurus punctatus) skin. Dev. Comp. Immunol. 2013, 39, 447–455. [Google Scholar] [CrossRef]
- Albert, T.K.; Laubinger, W.; Müller, S.; Hanisch, F.G.; Kalinski, T.; Meyer, F.; Hoffmann, W. Human intestinal TFF3 forms disulfide-linked heteromers with the Mucus-Associated FCGBP protein and is released by hydrogen sulfide. J. Proteome Res. 2010, 9, 3108–3117. [Google Scholar] [CrossRef]
- Kim, M.; Lee, S.; Yang, S.K.; Song, K.; Lee, I. Differential expression in histologically normal crypts of ulcerative colitis suggests primary crypt disorder. Oncol. Rep. 2006, 16, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Risques, R.A.; Lai, L.A.; Himmetoglu, C.; Ebaee, A.; Li, L.; Feng, Z.; Bronner, M.P.; Al-Lahham, B.; Kowdley, K.V.; Lindor, K.D.; et al. Ulcerative colitis-associated colorectal cancer arises in a field of short telomeres, senescence, and inflammation. Cancer Res. 2011, 71, 1669–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Chen, H.; Han, L.; Xue, F.; Wang, A.; Liang, Y.J. Screening of genes related to lung cancer caused by smoking with RNA-Seq. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 117–125. [Google Scholar] [PubMed]
- Wang, A.; Wu, L.; Lin, J.; Han, L.; Bian, J.; Wu, Y.; Robson, S.C.; Xue, L.; Ge, Y.; Sang, X.; et al. Whole-exome sequencing reveals the origin and evolution of hepato-cholangiocarcinoma. Nat. Commun. 2018, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Gazi, M.H.; He, M.; Cheville, J.C.; Young, C.Y. Downregulation of IgG Fc binding protein (Fc gammaBP) in prostate cancer. Cancer Biol. Ther. 2008, 7, 70–75. [Google Scholar] [CrossRef] [Green Version]
- O’Donovan, N.; Fischer, A.; Abdo, E.M.; Simon, F.; Peter, H.J.; Gerber, H.; Buergi, U.; Marti, U. Differential expression of IgG Fc binding protein (FcgammaBP) in human normal thyroid tissue, thyroid adenomas and thyroid carcinomas. J. Endocrinol. 2002, 174, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Wen, Y.; Miao, X.; Yang, Z. NT5E and FcGBP as key regulators of TGF-1-induced epithelial-mesenchymal transition (EMT) are associated with tumor progression and survival of patients with gallbladder cancer. Cell Tissue Res. 2014, 355, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Zhao, Z.; Hu, H.; Zhu, Y.; Zhang, W.; Tang, Q.; Huang, R.; Gao, F.; Zou, C.; Wang, G.; et al. IgG Fc Binding Protein (FCGBP) is Down-Regulated in Metastatic Lesions and Predicts Survival in Metastatic Colorectal Cancer Patients. Onco Targets Ther. 2021, 14, 967–977. [Google Scholar] [CrossRef]
- Choi, C.H.; Choi, J.J.; Park, Y.A.; Lee, Y.Y.; Song, S.Y.; Sung, C.O.; Song, T.; Kim, M.K.; Kim, T.J.; Lee, J.W.; et al. Identification of differentially expressed genes according to chemosensitivity in advanced ovarian serous adenocarcinomas: Expression of GRIA2 predicts better survival. Br. J. Cancer 2012, 107, 91–99. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Liu, H.; Zhang, Q.; Song, H.; Tang, J.; Fu, J.; Wang, X. FcGBP was upregulated by HPV infection and correlated to longer survival time of HNSCC patients. Oncotarget 2017, 8, 86503–86514. [Google Scholar] [CrossRef]
- Chen, S.M.Y.; Krinsky, A.L.; Woolaver, R.A.; Wang, X.; Chen, Z.; Wang, J.H. Tumor immune microenvironment in head and neck cancers. Mol. Carcinog. 2020, 59, 766–774. [Google Scholar] [CrossRef]
- Chow, L.Q.M.; Haddad, R.; Gupta, S.; Mahipal, A.; Mehra, R.; Tahara, M.; Berger, R.; Eder, J.P.; Burtness, B.; Lee, S.H.; et al. Antitumor Activity of Pembrolizumab in Biomarker-Unselected Patients with Recurrent and/or Metastatic Head and Neck Squamous Cell Carcinoma: Results from the Phase Ib KEYNOTE-012 Expansion Cohort. J. Clin. Oncol. 2016, 34, 3838–3845. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Rodriguez, I.P.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Chen, F.; Chandrashekar, D.S.; Varambally, S.; Creighton, C.J. Pan-cancer molecular subtypes revealed by mass-spectrometry-based proteomic characterization of more than 500 human cancers. Nat. Commun. 2019, 10, 5679. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef] [Green Version]
- Ru, B.; Wong, C.N.; Tong, Y.; Zhong, J.Y.; Zhong, S.S.W.; Wu, W.C.; Chu, K.C.; Wong, C.Y.; Lau, C.Y.; Chen, I.; et al. TISIDB: An integrated repository portal for tumor-immune system interactions. Bioinformatics 2019, 35, 4200–4202. [Google Scholar] [CrossRef]
- Vousden, K.H.; Prives, C. P53 and prognosis: New insights and further complexity. Cell 2005, 120, 7–10. [Google Scholar] [PubMed] [Green Version]
- Chu, B.; Kon, N.; Chen, D.; Li, T.; Liu, T.; Jiang, L.; Song, S.; Tavana, O.; Gu, W. ALOX12 is required for p53-mediated tumour suppression through a distinct ferroptosis pathway. Nat. Cell Biol. 2019, 21, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.P., IV; Yashinskie, J.J.; Koche, R.; Chandwani, R.; Tian, S.; Chen, C.C.; Baslan, T.; Marinkovic, Z.S.; Sánchez-Rivera, F.J.; Leach, S.D.; et al. α-Ketoglutarate links p53 to cell fate during tumour suppression. Nature 2019, 573, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Spike, B.T.; Wahl, G.M. p53, Stem Cells, and Reprogramming: Tumor Suppression beyond Guarding the Genome. Genes Cancer 2011, 2, 404–419. [Google Scholar] [CrossRef]
- Janic, A.; Valente, L.J.; Wakefield, M.J.; Di Stefano, L.; Milla, L.; Wilcox, S.; Yang, H.; Tai, L.; Vandenberg, C.J.; Kueh, A.J.; et al. DNA repair processes are critical mediators of p53-dependent tumor suppression. Nat. Med. 2018, 24, 947–953. [Google Scholar] [CrossRef]
- Blagih, J.; Zani, F.; Chakravarty, P.; Hennequart, M.; Pilley, S.; Hobor, S.; Hock, A.K.; Walton, J.B.; Morton, J.P.; Gronroos, E.; et al. Cancer-Specific Loss of p53 Leads to a Modulation of Myeloid and T Cell Responses. Cell Rep. 2020, 30, 481–496. [Google Scholar] [CrossRef]
- Bieging-Rolett, K.T.; Kaiser, A.M.; Morgens, D.W.; Boutelle, A.M.; Seoane, J.A.; Van Nostrand, E.L.; Zhu, C.; Houlihan, S.L.; Mello, S.S.; Yee, B.A.; et al. Zmat3 Is a Key Splicing Regulator in the p53 Tumor Suppression Program. Mol. Cell 2020, 80, 452–469. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, Z.; Myers, J.N. TP53 Mutations in Head and Neck Squamous Cell Carcinoma and Their Impact on Disease Progression and Treatment Response. J. Cell. Biochem. 2016, 117, 2682–2692. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Caponio, V.C.A.; Troiano, G.; Adipietro, I.; Zhurakivska, K.; Arena, C.; Mangieri, D.; Mascitti, M.; Cirillo, N.; Lo Muzio, L. Computational analysis of TP53 mutational landscape unveils key prognostic signatures and distinct pathobiological pathways in head and neck squamous cell cancer. Br. J. Cancer 2020, 123, 1302–1314. [Google Scholar] [CrossRef]
- Perrone, F.; Bossi, P.; Cortelazzi, B.; Locati, L.; Quattrone, P.; Pierotti, M.A.; Pilotti, S.; Licitra, L. TP53 mutations and pathologic complete response to neoadjuvant cisplatin and fluorouracil chemotherapy in resected oral cavity squamous cell carcinoma. J. Clin. Oncol. 2010, 28, 761–766. [Google Scholar] [CrossRef]
- Jagla, W.; Wiede, A.; Hinz, M.; Dietzmann, K.; Gülicher, D.; Gerlach, K.L.; Hoffmann, W. Secretion of TFF-peptides by human salivary glands. Cell Tissue Res. 1999, 298, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Kouznetsova, I.; Kalinski, T.; Peitz, U.; Mönkemüller, K.E.; Kalbacher, H.; Vieth, M.; Meyer, F.; Roessner, A.; Malfertheiner, P.; Lippert, H.; et al. Localization of TFF3 peptide in human esophageal submucosal glands and gastric cardia: Differentiation of two types of gastric pit cells along the rostro-caudal axis. Cell Tissue Res. 2007, 328, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W. Trefoil Factor Family (TFF) Peptides and their Different Roles in the Mucosal Innate Immune Defense and More: An Update. Curr. Med. Chem. 2021, 28, 7387–7399. [Google Scholar] [CrossRef]
- Jahan, R.; Shah, A.; Kisling, S.G.; Macha, M.A.; Thayer, S.; Batra, S.K.; Kaur, S. Odyssey of trefoil factors in cancer: Diagnostic and therapeutic implications. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2020, 1873, 188362. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.Y.; Wang, S.J.; Song, F.; Cheng, X.; Nan, G.; Zhao, Y.; Qian, M.R.; Chen, X.; Li, J.Y.; Liu, F.L.; et al. CD147 receptor is essential for TFF3-mediated signaling regulating colorectal cancer progression. Signal Transduct. Target. Ther. 2021, 6, 268. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kim, S.Y.; Shin, S.; Jung, S.H.; Yim, S.H.; Lee, J.Y.; Lee, S.H.; Chung, Y.J. Overexpression of TFF3 is involved in prostate carcinogenesis via blocking mitochondria-mediated apoptosis. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhang, X.; Zhang, J.; Liu, Y.; Ji, T.; Mou, J.; Fang, X.; Wang, S.; Chen, J. Low expression of TFF3 in papillary thyroid carcinoma may correlate with poor prognosis but high immune cell infiltration. Future Oncol. 2022, 18, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Nisar, S.; Yousuf, P.; Masoodi, T.; Wani, N.A.; Hashem, S.; Singh, M.; Sageena, G.; Mishra, D.; Kumar, R.; Haris, M.; et al. Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 4584. [Google Scholar] [CrossRef]
- Fridman, W.H.; Zitvogel, L.; Sautès-Fridman, C.; Kroemer, G. The immune contexture in cancer prognosis and treatment. Nat. Rev. Clin. Oncol. 2017, 14, 717–734. [Google Scholar] [CrossRef]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | FCBGP mRNA Level | ||
|---|---|---|---|
| Low | High | p Value | |
| n = 233 | n = 209 | ||
| Gender | 1.0 | ||
| Male | 170 | 152 | |
| Female | 63 | 57 | |
| AJCC 1 T Classification | <0.001 | ||
| T1/T2 | 72 | 99 | |
| T3/T4 | 161 | 110 | |
| AJCC 1 N Classification | 0.894 | ||
| N0 | 106 | 97 | |
| N1/N2/N3 | 127 | 112 | |
| Stage | 0.015 | ||
| Stage I | 7 | 20 | |
| Stage II | 33 | 37 | |
| Stage III | 43 | 38 | |
| Stage IV | 150 | 114 | |
| Cell Type | Marker | 1 HNSC (n = 522) | 2 HPV-Unrelated HNSC (n = 422) | HPV-Related HNSC (n = 98) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 Purity | 4 None | Purity | None | Purity | None | ||||||||
| 5 Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | ||
| B | |||||||||||||
| CD19 | 0.38 | *** | 0.38 | *** | 0.32 | *** | 0.33 | *** | 0.37 | *** | 0.32 | ** | |
| CD79A | 0.42 | *** | 0.41 | *** | 0.35 | *** | 0.36 | *** | 0.43 | *** | 0.37 | *** | |
| CD86 | 0.19 | *** | 0.20 | *** | 0.18 | *** | 0.19 | *** | 0.19 | 7.13 × 10−2 | 0.15 | 1.33 × 10−1 | |
| CSF1R | 0.34 | *** | 0.33 | *** | 0.32 | *** | 0.31 | *** | 0.38 | *** | 0.33 | ** | |
| Plasma | |||||||||||||
| CD38 | 0.11 | * | 0.13 | ** | 0.08 | 1.12 × 10−1 | 0.09 | 6.82 × 10−2 | 0.23 | * | 0.13 | ** | |
| CXCR4 | 0.31 | *** | 0.32 | *** | 0.26 | *** | 0.27 | *** | 0.34 | ** | 0.35 | *** | |
| TNFRSF17 | 0.37 | *** | 0.37 | *** | 0.31 | *** | 0.32 | *** | 0.39 | *** | 0.34 | *** | |
| CD27 | 0.41 | *** | 0.40 | *** | 0.35 | *** | 0.35 | *** | 0.44 | *** | 0.38 | *** | |
| CD8+ T | |||||||||||||
| CD8A | 0.28 | *** | 0.29 | *** | 0.21 | *** | 0.23 | *** | 0.38 | *** | 0.35 | *** | |
| CD8B | 0.28 | *** | 0.29 | *** | 0.20 | *** | 0.22 | *** | 0.40 | *** | 0.41 | *** | |
| Follicular helper T | |||||||||||||
| IL21 | 0.23 | *** | 0.24 | *** | 0.16 | ** | 0.18 | *** | 0.34 | ** | 0.32 | ** | |
| BCL6 | 0.30 | *** | 0.28 | *** | 0.29 | *** | 0.26 | *** | 0.25 | 1.71 × 10−2 | 0.22 | * | |
| ICOS | 0.29 | *** | 0.29 | *** | 0.25 | *** | 0.26 | *** | 0.32 | ** | 0.32 | ** | |
| CXCR5 | 0.42 | *** | 0.41 | *** | 0.38 | *** | 0.38 | *** | 0.37 | *** | 0.33 | *** | |
| Th17 | |||||||||||||
| STAT3 | 0.37 | *** | 0.37 | *** | 0.32 | *** | 0.31 | *** | 0.50 | *** | 0.49 | *** | |
| IL-17A | 0.34 | *** | 0.34 | *** | 0.26 | *** | 0.28 | *** | 0.49 | *** | 1.74 | *** | |
| IL-21R | 0.41 | *** | 0.40 | *** | 0.35 | *** | 0.35 | *** | 0.48 | *** | 1.47 | *** | |
| IL-23R | 0.31 | *** | 0.31 | *** | 0.27 | *** | 0.27 | *** | 0.38 | *** | 0.39 | *** | |
| T cell exhaustion | |||||||||||||
| PDCD1 | 0.30 | *** | 0.31 | *** | 0.22 | *** | 0.24 | *** | 0.42 | *** | 0.39 | *** | |
| CTLA4 | 0.30 | *** | 0.30 | *** | 0.23 | *** | 0.25 | *** | 0.40 | *** | 0.39 | *** | |
| LAG3 | 0.16 | *** | 0.17 | *** | 0.09 | 6.85 × 10−2 | 0.11 | * | 0.29 | ** | 0.28 | ** | |
| HAVCR2 | 0.23 | *** | 0.22 | *** | 0.18 | *** | 0.19 | *** | 0.29 | ** | 0.23 | * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-H.; Yang, Y.-F.; Shiue, Y.-L. Multi-Omics Analyses to Identify FCGBP as a Potential Predictor in Head and Neck Squamous Cell Carcinoma. Diagnostics 2022, 12, 1178. https://doi.org/10.3390/diagnostics12051178
Lin Y-H, Yang Y-F, Shiue Y-L. Multi-Omics Analyses to Identify FCGBP as a Potential Predictor in Head and Neck Squamous Cell Carcinoma. Diagnostics. 2022; 12(5):1178. https://doi.org/10.3390/diagnostics12051178
Chicago/Turabian StyleLin, Yu-Hsuan, Yi-Fang Yang, and Yow-Ling Shiue. 2022. "Multi-Omics Analyses to Identify FCGBP as a Potential Predictor in Head and Neck Squamous Cell Carcinoma" Diagnostics 12, no. 5: 1178. https://doi.org/10.3390/diagnostics12051178