
Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus
 , ,
, ,
Abstract
:1. Introduction
2. Case Presentation
3. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Serag, H.B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 2012, 142, 1264–1273.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trépo, C.; Chan, H.L.Y.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- Kao, J. Molecular Epidemiology of Hepatitis B Virus. Korean J. Intern. Med. 2011, 26, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.; Tanaka, Y.; Kurbanov, F.; Sugauchi, F.; Mano, S.; Maeshiro, T.; Nakayoshi, T.; Wakuta, M.; Miyakawa, Y.; Mizokami, M. A genetic variant of hepatitis B virus divergent from known human and ape genotypes isolated from a Japanese patient and provisionally assigned to new genotype J. J. Virol. 2009, 83, 10538–10547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, S. Hepatitis B virus taxonomy and hepatitis B virus genotypes. World J. Gastroenterol. 2007, 13, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Zehender, G.; Ebranati, E.; Gabanelli, E.; Sorrentino, C.; Lo Presti, A.; Tanzi, E.; Ciccozzi, M.; Galli, M. Enigmatic origin of hepatitis B virus: An ancient travelling companion or a recent encounter? World J. Gastroenterol. 2014, 20, 7622–7634. [Google Scholar] [CrossRef]
- Patel, J.; Borucki, R.; Werth, V.P. An Update on the Pathogenesis of Cutaneous Lupus Erythematosus and Its Role in Clinical Practice. Curr. Rheumatol. Rep. 2020, 22, 69. [Google Scholar] [CrossRef]
- Garelli, C.J.; Refat, M.A.; Nanaware, P.P.; Ramirez-Ortiz, Z.G.; Rashighi, M.; Richmond, J.M. Current Insights in Cutaneous Lupus Erythematosus Immunopathogenesis. Front. Immunol. 2020, 11, 1353. [Google Scholar] [CrossRef]
- Klein, R.S.; Morganroth, P.A.; Werth, V.P. Cutaneous lupus and the Cutaneous Lupus Erythematous Disease Area and Severity Index instrument. Rheum. Dis. Clin. N. Am. 2010, 36, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, A.; Ceccarelli, F.; Dorrucci, M.; Farchi, F.; Pirone, C.; Garufi, C.; Valdarchi, C.; Spinelli, F.R.; Alessandri, C.; Chionne, P.; et al. Hepatitis B and C Virus Infection in Patients with Systemic and Cutaneous Lupus Erythematosus. New Microbiol. 2022. accepted. In Press on 1/2023 of New Microbiologica. [Google Scholar]
- Nelson, P.; Rylance, P.; Roden, D.; Trela, M.; Tugnet, N. Viruses as potential pathogenic agents in systemic lupus erythematosus. Lupus 2014, 23, 596–605. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Tsokos, G.C. Systemic lupus erythematosus. N. Engl. J. Med. 2022, 365, 2110–2121. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, F.; Perricone, C.; Borgiani, P.; Ciccacci, C.; Rufini, S.; Cipriano, E.; Alessandri, C.; Spinelli, F.R.; Scavalli, A.S.; Novelli, G.; et al. Genetic Factors in Systemic Lupus Erythematosus: Contribution to Disease Phenotype. J. Immunol. Res. 2015, 2015, 745647. [Google Scholar] [CrossRef] [Green Version]
- Rigante, D.; Mazzoni, M.B.; Esposito, S. The cryptic interplay between systemic lupus erythematosus and infections. Autoimmun. Rev. 2014, 13, 96–102. [Google Scholar] [CrossRef]
- Rigante, D.; Esposito, S. Infections and Systemic Lupus Erythematosus: Binding or Sparring Partners? Int. J. Mol. Sci. 2015, 16, 17331–17343. [Google Scholar] [CrossRef] [Green Version]
- Rajoriya, N.; Combet, C.; Zoulim, F.; Janssen, H.L.A. How viral genetic variants and genotypes influence disease and treatment outcome of chronic hepatitis B. Time for an individualised approach? J. Hepatol. 2017, 67, 1281–1297. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, Y.; Xu, M.; Li, X.; Zhang, Z. Distribution of hepatitis B virus genotypes and subgenotypes. Medicine 2021, 100, e27941. [Google Scholar] [CrossRef] [PubMed]
- Sagnelli, C.; Ciccozzi, M.; Pisaturo, M.; Zehender, G.; Lo Presti, A.; Alessio, L.; Starace, M.; Lovero, D.; Sagnelli, E.; Coppola, N. Molecular epidemiology of hepatitis B virus genotypes circulating in acute hepatitis B patients in the Campania region. J. Med. Virol. 2014, 86, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- De Maddalena, C.; Giambelli, C.; Tanzi, E.; Colzani, D.; Schiavini, M.; Milazzo, L.; Bernini, F.; Ebranati, E.; Cargnel, A.; Bruno, R.; et al. High level of genetic heterogeneity in S and P genes of genotype D hepatitis B virus. Virology 2007, 365, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chachá, S.G.F.; Gomes-Gouvêa, M.S.; de Mello Malta, F.; da Costa Ferreira, S.; Villanova, M.G.; Souza, F.F.; Teixeira, A.C.; da Costa Passos, A.D.; Pinho, J.R.R.; de Lourdes Candolo Martinelli, A. Distribution of HBV subgenotypes in Ribeirão Preto, Southeastern Brazil: A region with history of intense Italian immigration. Braz. J. Infect. Dis. 2017, 21, 424–432. [Google Scholar] [CrossRef]
- Sheldon, J.; Rodès, B.; Zoulim, F.; Bartholomeusz, A.; Soriano, V. Mutations affecting the replication capacity of the hepatitis B virus. J. Viral Hepat. 2006, 13, 427–434. [Google Scholar] [CrossRef]
- Torresi, J. The virological and clinical significance of mutations in the overlapping envelope and polymerase genes of hepatitis B virus. J. Clin. Virol. 2002, 25, 97–106. [Google Scholar] [CrossRef]
- Zhang, M.; Ge, G.; Yang, Y.; Cai, X.; Fu, Q.; Cai, J.; Huang, Z. Decreased antigenicity profiles of immune-escaped and drug-resistant hepatitis B surface antigen. Virol. J. 2013, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Lazarevic, I.; Banko, A.; Miljanovic, D.; Cupic, M. Immune-Escape Hepatitis B Virus Mutations Associated with Viral Reactivation upon Immunosuppression. Viruses 2019, 11, 778. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Leung, P.S.C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.M. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123. [Google Scholar] [CrossRef]
- Mishra, R.; Bhattacharya, S.; Rawat, B.S.; Kumar, A.; Kumar, A.; Niraj, K.; Chande, A.; Gandhi, P.; Khetan, D.; Aggarwal, A.; et al. MicroRNA-30e-5p has an Integrated Role in the Regulation of the Innate Immune Response during Virus Infection and Systemic Lupus Erythematosus. iScience 2020, 23, 101322. [Google Scholar] [CrossRef]



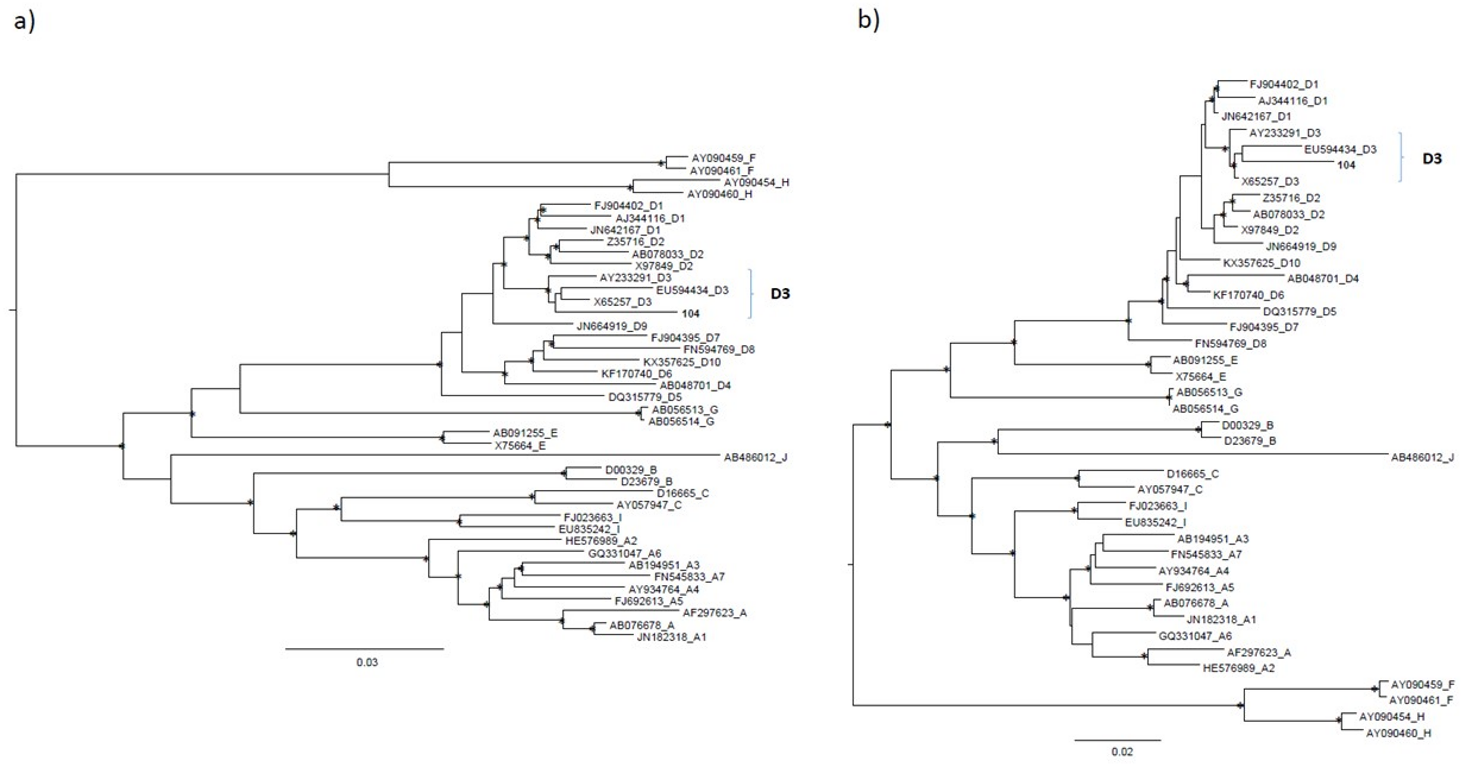{kind=link}
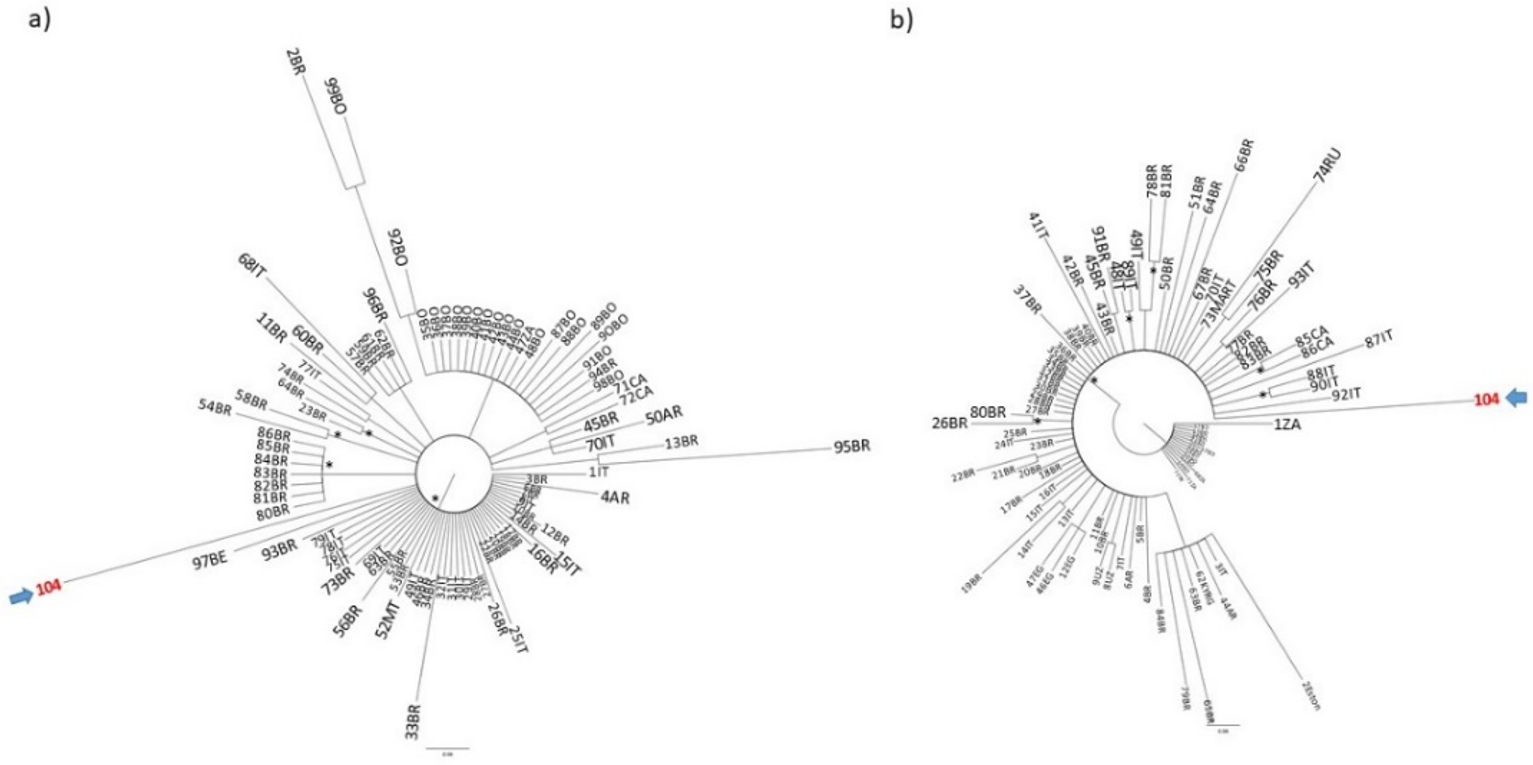{kind=link}
| Isolate 104 | Amino Acid in Reference Acc Number: X65257 | |
|---|---|---|
| (a) | ||
| polymerase | 339 H | Y |
| 351G # | W | |
| 402H | Y | |
| 468G | R | |
| 526Y ° | F | |
| 578C § | S | |
| HBsAg | 168M # | I |
| 266 T | I | |
| 268 S | L | |
| 344 I ° | F | |
| 381 N | S | |
| 394 S § | F | |
| 396 V | L | |
| (b) | ||
| polymerase | 351G | W |
| 526Y | F | |
| 578C | S | |
| 168M | I | |
| 268 S | L | |
| HBsAg | 344 I | F |
| 394 S | F | |
| 396 V | L | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villano, U.; Mataj, E.; Dorrucci, M.; Farchi, F.; Pirone, C.; Valdarchi, C.; Equestre, M.; Madonna, E.; Bruni, R.; Pisani, G.; et al. Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus. Diagnostics 2022, 12, 2866. https://doi.org/10.3390/diagnostics12112866
Villano U, Mataj E, Dorrucci M, Farchi F, Pirone C, Valdarchi C, Equestre M, Madonna E, Bruni R, Pisani G, et al. Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus. Diagnostics. 2022; 12(11):2866. https://doi.org/10.3390/diagnostics12112866
Chicago/Turabian StyleVillano, Umbertina, Elida Mataj, Maria Dorrucci, Francesca Farchi, Carmelo Pirone, Catia Valdarchi, Michele Equestre, Elisabetta Madonna, Roberto Bruni, Giulio Pisani, and et al. 2022. "Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus" Diagnostics 12, no. 11: 2866. https://doi.org/10.3390/diagnostics12112866