
Host-Pathogen Adhesion as the Basis of Innovative Diagnostics for Emerging Pathogens

 , , , ,
, , , ,  , , , ,
, , , ,
Abstract
:1. Introduction
1.1. Microbial Adhesion, Colonization and Host Infection
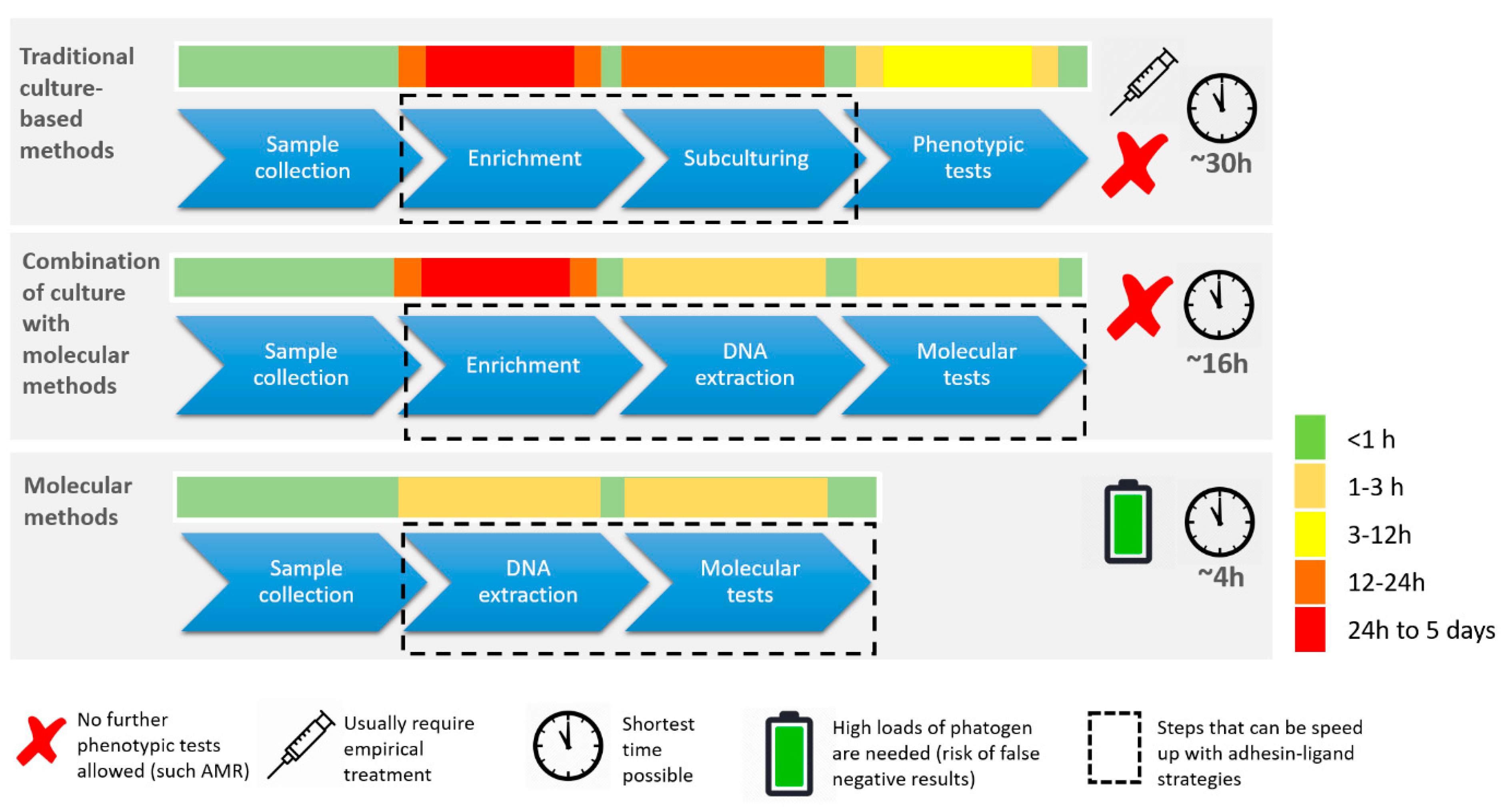
1.2. Current State of Infectious Disease Diagnostics
1.3. Mandatory Improvement of IVD
2. Microbial Adhesin–Receptor Pairs
2.1. Viral Adhesion Processes
2.2. Modes of Bacterial Adhesion
2.3. Adhesion Diversity and Evolution
3. Structural Analysis of Adhesin–Ligand Pairs
Technological and Methodological Developments
4. Microbial Adhesion and Future High-Throughput Diagnostic Microbiology Technology
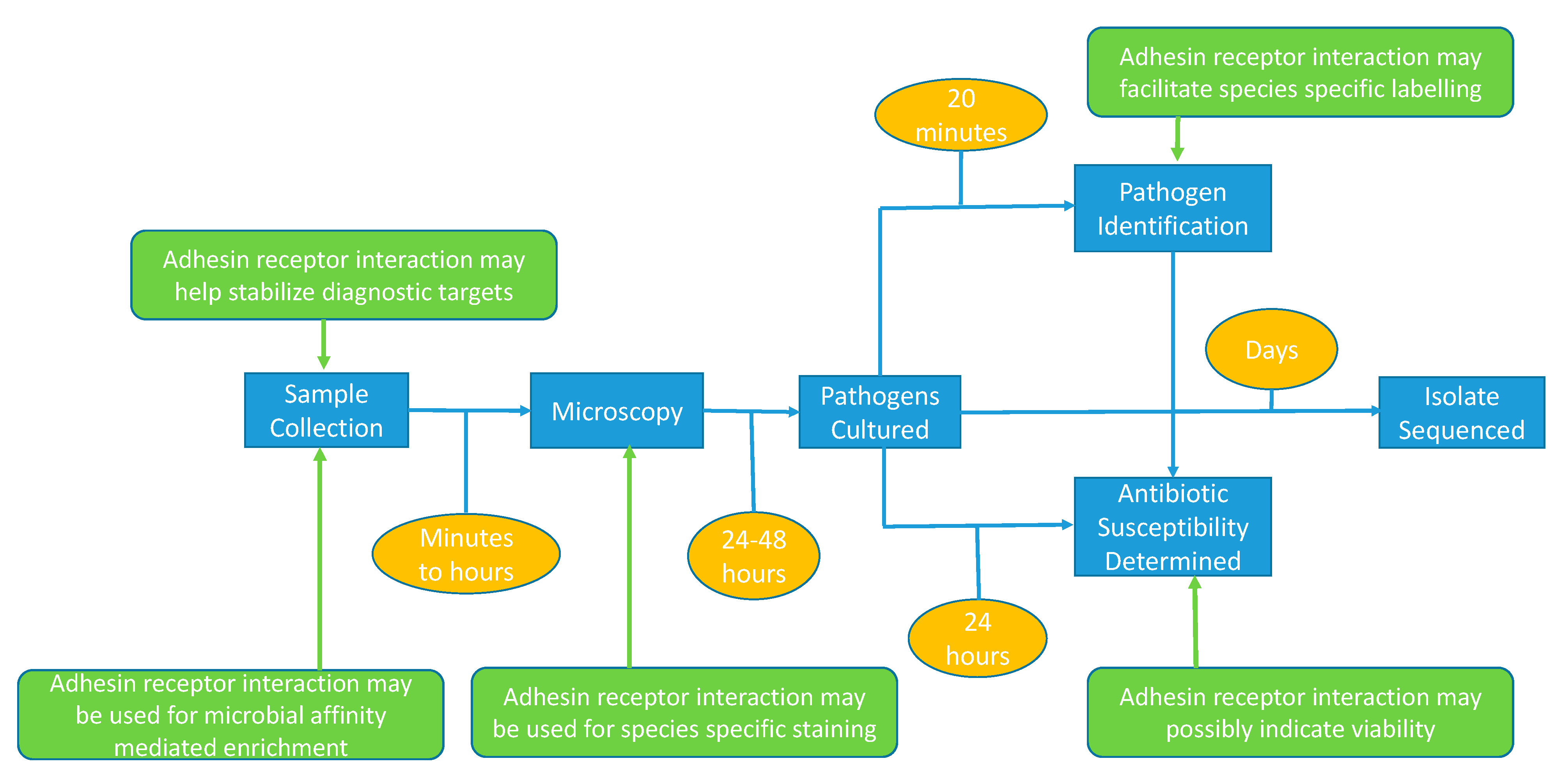
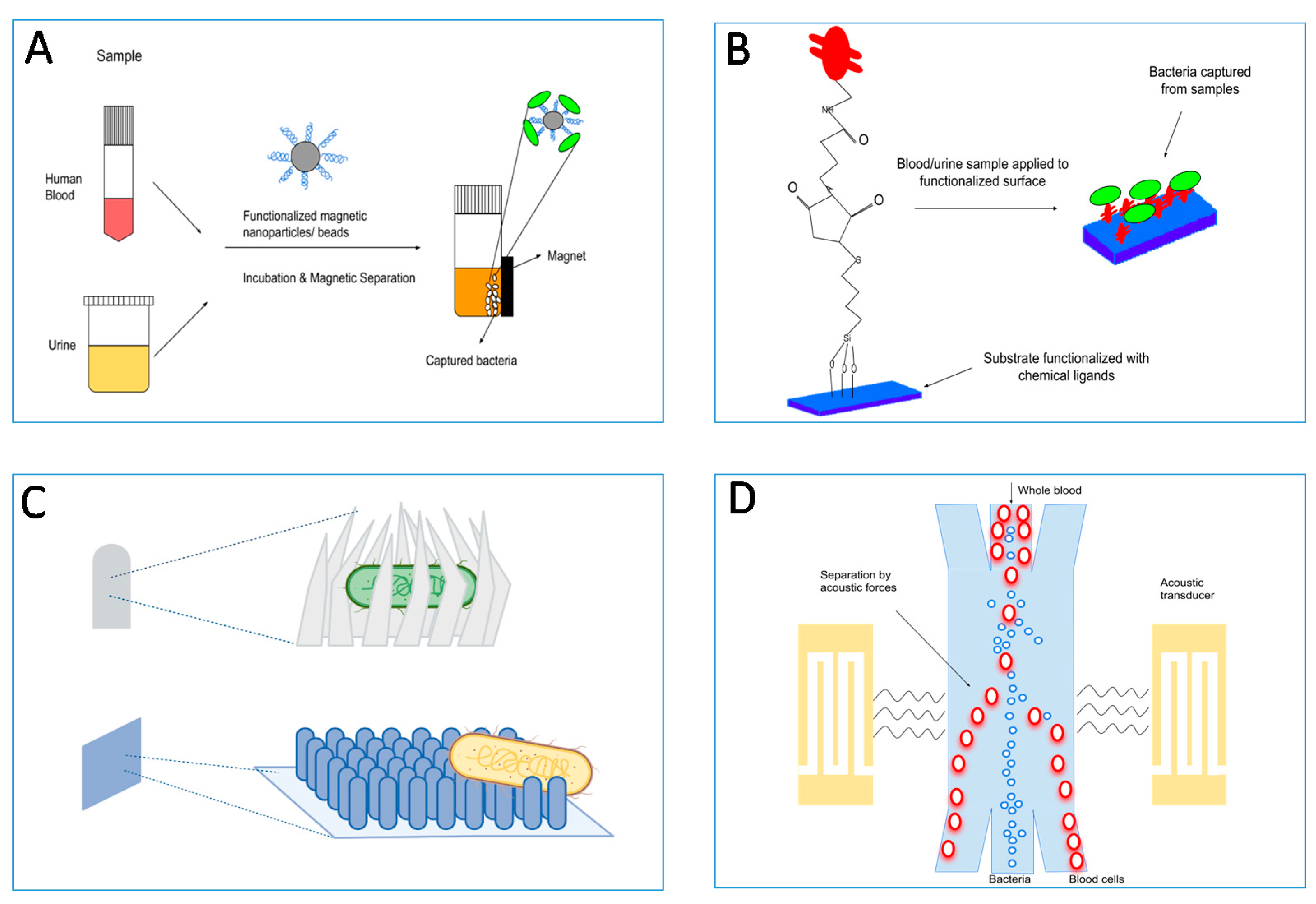
4.1. Adhesin-Based Sample Processing in Microbial Diagnostics
4.2. Target Enrichment Technology
5. Clinical–Diagnostic Application of Pathogen Adhesion Tests
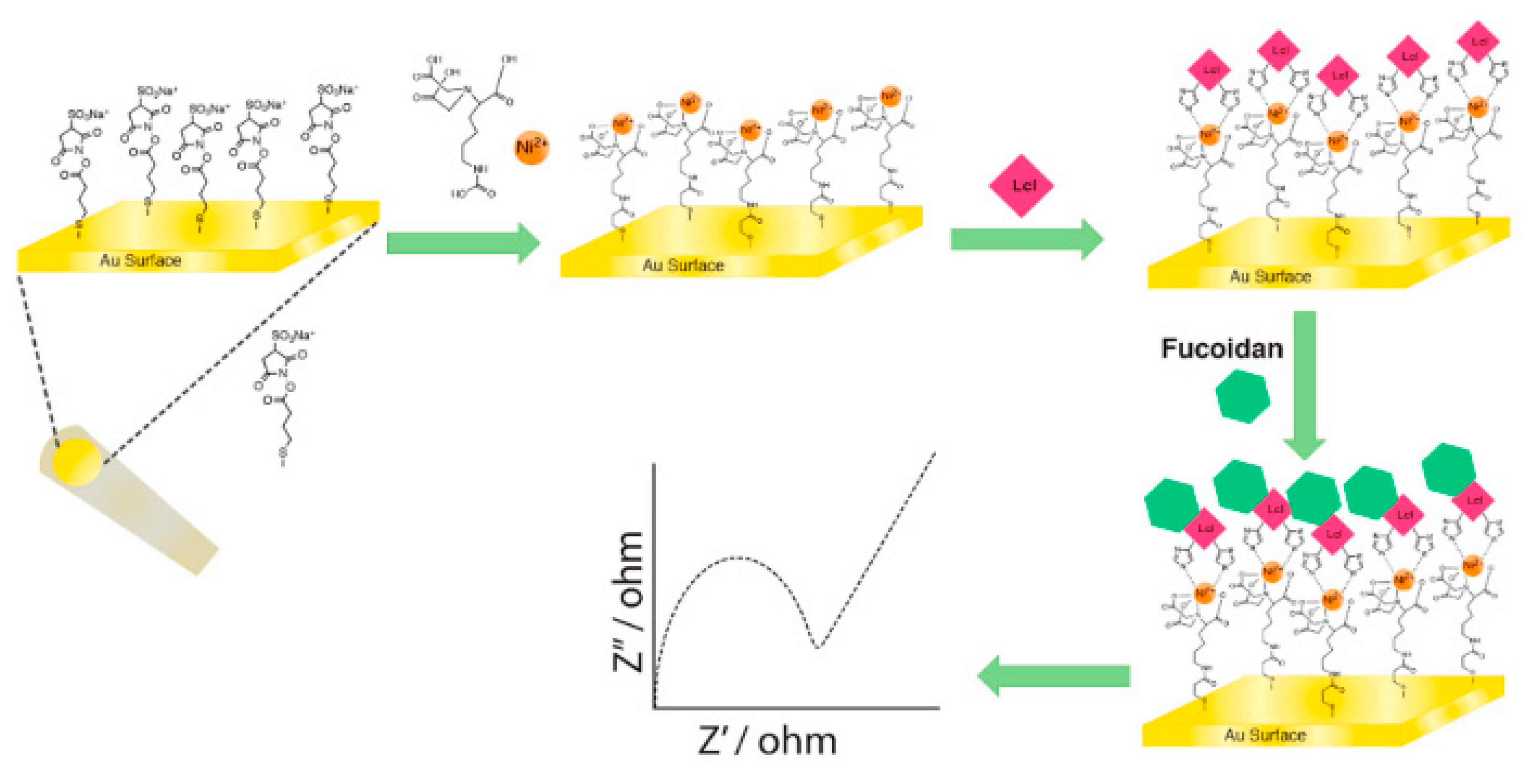
5.1. Biosensor-Based Pathogen Detection
5.2. Next Generation Test Formats—Bioactive Surfaces and Materials
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Markwell, A.; Mitchell, R.; Wright, A.L.; Brown, A.F. Clinical and ethical challenges for emergency departments during communicable disease outbreaks: Can lessons from Ebola Virus Disease be applied to the COVID-19 pandemic? Emerg. Med. Australas. 2020, 32, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Suetens, C.; Latour, K.; Kärki, T.; Ricchizzi, E.; Kinross, P.; Moro, M.L.; Jans, B.; Hopkins, S.; Hansen, S.; Lyytikäinen, O.; et al. Prevalence of healthcare-associated infections, estimated incidence and composite antimicrobial resistance index in acute care hospitals and long-term care facilities: Results from two European point prevalence surveys, 2016 to 2017. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef] [Green Version]
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global Contributors to Antibiotic Resistance. J. Glob. Infect. Dis. 2019, 11, 36–42. [Google Scholar] [CrossRef]
- Zumla, A.; Memish, Z.A.; Maeurer, M.; Bates, M.; Mwaba, P.; Al-Tawfiq, J.A.; Denning, D.W.; Hayden, F.G.; Hui, D.S. Emerging novel and antimicrobial-resistant respiratory tract infections: New drug development and therapeutic options. Lancet. Infect. Dis. 2014, 14, 1136–1149. [Google Scholar] [CrossRef]
- Sloots, T.P.; Nissen, M.D.; Ginn, A.N.; Iredell, J.R. Rapid identification of pathogens using molecular techniques. Pathology 2015, 47, 191–198. [Google Scholar] [CrossRef]
- Pinsky, B.A.; Hayden, R.T. Cost-Effective Respiratory Virus Testing. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Yamazaki, K.; Kashimoto, T.; Kado, T.; Akeda, Y.; Yoshioka, K.; Kodama, T.; Yamamoto, M.; Okamura, M.; Kakuda, T.; Ueno, S. Chemotactic invasion in deep soft tissue by Vibrio vulnificus is essential for the progression of necrotic lesions. Virulence 2020, 11, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Phukan, U.J. Interaction of host and Staphylococcus aureus protease-system regulates virulence and pathogenicity. Med. Microbiol. Immunol. 2019, 208, 585–607. [Google Scholar] [CrossRef] [PubMed]
- Comunian, S.; Dongo, D.; Milani, C.; Palestini, P. Air Pollution and COVID-19: The Role of Particulate Matter in the Spread and Increase of COVID-19’s Morbidity and Mortality. Int. J. Environ. Res. Public. Health. 2020, 17, 4487. [Google Scholar] [CrossRef] [PubMed]
- Speziale, P.; Arciola, C.R.; Pietrocola, G. Fibronectin and Its Role in Human Infective Diseases. Cells 2019, 8, 1516. [Google Scholar] [CrossRef] [Green Version]
- Duell, B.L.; Su, Y.C.; Riesbeck, K. Host-pathogen interactions of nontypeable Haemophilus influenzae: From commensal to pathogen. FEBS Lett. 2016, 590, 3840–3853. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Pietrocola, G.; Arciola, C.R.; Rindi, S.; Montanaro, L.; Speziale, P. Streptococcus agalactiae Non-Pilus, Cell Wall-Anchored Proteins: Involvement in Colonization and Pathogenesis and Potential as Vaccine Candidates. Front. Immunol. 2018, 9, 602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, R.J. Receptor mimicry as novel therapeutic treatment for biothreat agents. Bioeng. Bugs. 2010, 1, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.B.; Atrzadeh, F.; Burnham, C.A.; Cavalieri, S.; Dunn, J.; Jones, S.; Mathews, C.; McNult, P.; Meduri, J.; Newhouse, C.; et al. Clinical Utility of Advanced Microbiology Testing Tools. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Dubourg, G.; Lamy, B.; Ruimy, R. Rapid phenotypic methods to improve the diagnosis of bacterial bloodstream infections: Meeting the challenge to reduce the time to result. Clin. Microbiol. Infect. 2018, 24, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, O.; Durand, G.; Hallin, M.; Diefenbach, A.; Gant, V.; Murray, P.; Kozlakidis, Z.; van Belkum, A. Consolidation of Clinical Microbiology Laboratories and Introduction of Transformative Technologies. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in virus diagnostics: From ELISA to next generation sequencing. Virus. Res. 2014, 186, 20–31. [Google Scholar] [CrossRef]
- Han, D.; Li, Z.; Li, R.; Tan, P.; Zhang, R.; Li, J. mNGS in clinical microbiology laboratories: On the road to maturity. Crit. Rev. Microbiol. 2019, 45, 668–685. [Google Scholar] [CrossRef]
- van Belkum, A.; Bachmann, T.T.; Lüdke, G.; Lisby, J.G.; Kahlmeter, G.; Mohess, A.; Becker, K.; Hays, J.P.; Woodford, N.; Mitsakakis, K.; et al. Developmental roadmap for antimicrobial susceptibility testing systems. Nat. Rev. Microbiol. 2019, 17, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, O.; Martiny, D.; Rochas, O.; van Belkum, A.; Kozlakidis, Z. Considerations for diagnostic COVID-19 tests. Nat. Rev. Microbiol. 2021, 19, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.L.; St George, K.; Rhoads, D.D.; Butler-Wu, S.M.; Dharmarha, V.; McNult, P.; Miller, M.B. Understanding, Verifying, and Implementing Emergency Use Authorization Molecular Diagnostics for the Detection of SARS-CoV-2 RNA. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef]
- Andryukov, B.G.; Besednova, N.N.; Romashko, R.V.; Zaporozhets, T.S.; Efimov, T.A. Label-Free Biosensors for Laboratory-Based Diagnostics of Infections: Current Achievements and New Trends. Biosensors 2020, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Bedford, R.; Tiede, C.; Hughes, R.; Curd, A.; McPherson, M.J.; Peckham, M.; Tomlinson, D.C. Alternative reagents to antibodies in imaging applications. Biophys. Rev. 2017, 9, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, X.; Fu, X.; Li, Z.; Huang, Y.; Liang, C. Advances in Aptamer-Based Biomarker Discovery. Front. Cell. Dev. Biol. 2021, 9, 659760. [Google Scholar] [CrossRef]
- Sharon, N.; Ofek, I. Identification of receptors for bacterial lectins by blotting techniques. Methods Enzymol. 1995, 253, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Kitts, G.; Giglio, K.M.; Zamorano-Sánchez, D.; Park, J.H.; Townsley, L.; Cooley, R.B.; Wucher, B.R.; Klose, K.E.; Nadell, C.D.; Yildiz, F.H.; et al. A Conserved Regulatory Circuit Controls Large Adhesins in Vibrio cholerae. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chen, W.; Xia, A.; Zhang, R.; Huang, Y.; Yang, S.; Ni, L.; Jin, F. Carbon Starvation Induces the Expression of PprB-Regulated Genes in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Sequeira, S.; Kavanaugh, D.; MacKenzie, D.A.; Šuligoj, T.; Walpole, S.; Leclaire, C.; Gunning, A.P.; Latousakis, D.; Willats, W.G.T.; Angulo, J.; et al. Structural basis for the role of serine-rich repeat proteins from Lactobacillus reuteri in gut microbe-host interactions. Proc. Natl. Acad. Sci. USA 2018, 115, E2706–E2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loukachevitch, L.V.; Bensing, B.A.; Yu, H.; Zeng, J.; Chen, X.; Sullam, P.M.; Iverson, T.M. Structures of the Streptococcus sanguinis SrpA Binding Region with Human Sialoglycans Suggest Features of the Physiological Ligand. Biochemistry 2016, 55, 5927–5937. [Google Scholar] [CrossRef]
- Peacock, T.P.; Penrice-Randal, R.; Hiscox, J.A.; Barclay, W.S. SARS-CoV-2 one year on: Evidence for ongoing viral adaptation. J. Gen. Virol. 2021, 102. [Google Scholar] [CrossRef]
- Sherman, M.B.; Smith, H.Q.; Smith, T.J. The Dynamic Life of Virus Capsids. Viruses 2020, 12, 618. [Google Scholar] [CrossRef]
- Neu, U.; Bauer, J.; Stehle, T. Viruses and sialic acids: Rules of engagement. Curr. Opin. Struct. Biol. 2011, 21, 610–618. [Google Scholar] [CrossRef]
- Stroh, L.J.; Stehle, T. Glycan Engagement by Viruses: Receptor Switches and Specificity. Annu. Rev. Virol. 2014, 1, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Hiatt, A.; Bohorova, N.; Bohorov, O.; Goodman, C.; Kim, D.; Pauly, M.H.; Velasco, J.; Whaley, K.J.; Piedra, P.A.; Gilbert, B.E.; et al. Glycan variants of a respiratory syncytial virus antibody with enhanced effector function and in vivo efficacy. Proc. Natl. Acad. Sci. USA 2014, 111, 5992–5997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, T.; Okamoto, M.; Nakakita, S.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.; Roy, C.N.; Hiramatsu, H.; Sugawara, K.E.; et al. Antigenic and receptor binding properties of enterovirus 68. J. Virol. 2014, 88, 2374–2384. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, Y.; Goda, T.; Matsumoto, A.; Takeuchi, H.; Yamaoka, S.; Miyahara, Y. Direct and label-free influenza virus detection based on multisite binding to sialic acid receptors. Biosens. Bioelectron. 2017, 92, 234–240. [Google Scholar] [CrossRef]
- Naskalska, A.; Dabrowska, A.; Szczepański, A.; Milewska, A.; Jasik, K.P.; Pyrc, K. Membrane Protein of Human Coronavirus NL63 Is Responsible for Interaction with the Adhesion Receptor. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Gómez, D.; Domínguez-Soto, A.; Ancochea, J.; Jimenez-Heffernan, J.A.; Leal, J.A.; Corbí, A.L. Dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin mediates binding and internalization of Aspergillus fumigatus conidia by dendritic cells and macrophages. J. Immunol. 2004, 173, 5635–5643. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.C.; Feng, Z.H.; Nie, Q.H.; Zhou, Y.X. DC-SIGN: Binding receptors for hepatitis C virus. Chin. Med. J. 2004, 117, 1395–1400. [Google Scholar] [PubMed]
- Li, H.; Liu, L.; Zhang, D.; Xu, J.; Dai, H.; Tang, N.; Su, X.; Cao, B. SARS-CoV-2 and viral sepsis: Observations and hypotheses. Lancet 2020, 395, 1517–1520. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Jeffers, S.A.; Tusell, S.M.; Gillim-Ross, L.; Hemmila, E.M.; Achenbach, J.E.; Babcock, G.J.; Thomas, W.D., Jr.; Thackray, L.B.; Young, M.D.; Mason, R.J.; et al. CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proc. Natl. Acad. Sci. USA 2004, 101, 15748–15753. [Google Scholar] [CrossRef] [Green Version]
- Amraie, R.; Napoleon, M.A.; Yin, W.; Berrigan, J.; Suder, E.; Zhao, G.; Olejnik, J.; Gummuluru, S.; Muhlberger, E.; Chitalia, V.; et al. CD209L/L-SIGN and CD209/DC-SIGN act as receptors for SARS-CoV-2 and are differentially expressed in lung and kidney epithelial and endothelial cells. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Ke, B.; Feng, Q.; Yang, D.; Lian, Q.; Li, Z.; Lu, L.; Ke, C.; Liu, Z.; Liao, G. Construction and immunogenic studies of a mFc fusion receptor binding domain (RBD) of spike protein as a subunit vaccine against SARS-CoV-2 infection. Chem. Commun. 2020, 56, 8683–8686. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of M(pro) from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Viela, F.; Mathelié-Guinlet, M.; Viljoen, A.; Dufrêne, Y.F. What makes bacterial pathogens so sticky? Mol. Microbiol. 2020, 113, 683–690. [Google Scholar] [CrossRef]
- Kwon, H.; Squire, C.J.; Young, P.G.; Baker, E.N. Autocatalytically generated Thr-Gln ester bond cross-links stabilize the repetitive Ig-domain shaft of a bacterial cell surface adhesin. Proc. Natl. Acad. Sci. USA 2014, 111, 1367–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrobel, A.; Ottoni, C.; Leo, J.C.; Gulla, S.; Linke, D. The repeat structure of two paralogous genes, Yersinia ruckeri invasin (yrInv) and a “Y. ruckeri invasin-like molecule”, (yrIlm) sheds light on the evolution of adhesive capacities of a fish pathogen. J. Struct. Biol. 2018, 201, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linke, D.; Riess, T.; Autenrieth, I.B.; Lupas, A.; Kempf, V.A. Trimeric autotransporter adhesins: Variable structure, common function. Trends Microbiol. 2006, 14, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.F.; Kaiser, P.O.; Linke, D.; Schwarz, H.; Riess, T.; Schäfer, A.; Eble, J.A.; Kempf, V.A. Trimeric autotransporter adhesin-dependent adherence of Bartonella henselae, Bartonella quintana, and Yersinia enterocolitica to matrix components and endothelial cells under static and dynamic flow conditions. Infect. Immun. 2011, 79, 2544–2553. [Google Scholar] [CrossRef] [Green Version]
- Weidensdorfer, M.; Chae, J.I.; Makobe, C.; Stahl, J.; Averhoff, B.; Müller, V.; Schürmann, C.; Brandes, R.P.; Wilharm, G.; Ballhorn, W.; et al. Analysis of Endothelial Adherence of Bartonella henselae and Acinetobacter baumannii Using a Dynamic Human Ex Vivo Infection Model. Infect. Immun. 2015, 84, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Meuskens, I.; Saragliadis, A.; Leo, J.C.; Linke, D. Type V Secretion Systems: An Overview of Passenger Domain Functions. Front. Microbiol. 2019, 10, 1163. [Google Scholar] [CrossRef] [Green Version]
- Koiwai, K.; Hartmann, M.D.; Linke, D.; Lupas, A.N.; Hori, K. Structural Basis for Toughness and Flexibility in the C-terminal Passenger Domain of an Acinetobacter Trimeric Autotransporter Adhesin. J. Biol. Chem. 2016, 291, 3705–3724. [Google Scholar] [CrossRef] [Green Version]
- Bassler, J.; Hernandez Alvarez, B.; Hartmann, M.D.; Lupas, A.N. A domain dictionary of trimeric autotransporter adhesins. Int. J. Med. Microbiol. 2015, 305, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Shin, P.K.; Pawar, P.; Konstantopoulos, K.; Ross, J.M. Characteristics of new Staphylococcus aureus-RBC adhesion mechanism independent of fibrinogen and IgG under hydrodynamic shear conditions. Am. J. Physiol. Cell. Physiol. 2005, 289, C727–C734. [Google Scholar] [CrossRef] [Green Version]
- Reichhardt, C.; Wong, C.; Passos da Silva, D.; Wozniak, D.J.; Parsek, M.R. CdrA Interactions within the Pseudomonas aeruginosa Biofilm Matrix Safeguard It from Proteolysis and Promote Cellular Packing. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J. The remarkably multifunctional fibronectin binding proteins of Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1923–1931. [Google Scholar] [CrossRef] [PubMed]
- Stones, D.H.; Krachler, A.M. Fatal attraction: How bacterial adhesins affect host signaling and what we can learn from them. Int. J. Mol. Sci. 2015, 16, 2626–2640. [Google Scholar] [CrossRef] [Green Version]
- Adrian, J.; Bonsignore, P.; Hammer, S.; Frickey, T.; Hauck, C.R. Adaptation to Host-Specific Bacterial Pathogens Drives Rapid Evolution of a Human Innate Immune Receptor. Curr. Biol. 2019, 29, 616–630.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, T.D.; Annis, D.S.; O’Neill, M.B.; Bohr, L.L.; Smith, T.M.; Poinar, H.N.; Mosher, D.F.; Pepperell, C.S. Adaptation in a Fibronectin Binding Autolysin of Staphylococcus saprophyticus. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, R.; Markus, B.; Carstens Marques de Sousa, K.; Marcos-Hadad, E.; Mugasimangalam, R.C.; Nachum-Biala, Y.; Hawlena, H.; Covo, S.; Harrus, S. Prophage-Driven Genomic Structural Changes Promote Bartonella Vertical Evolution. Genome. Biol. Evol. 2018, 10, 3089–3103. [Google Scholar] [CrossRef] [PubMed]
- Lamelas, A.; Harris, S.R.; Röltgen, K.; Dangy, J.P.; Hauser, J.; Kingsley, R.A.; Connor, T.R.; Sie, A.; Hodgson, A.; Dougan, G.; et al. Emergence of a new epidemic Neisseria meningitidis serogroup A Clone in the African meningitis belt: High-resolution picture of genomic changes that mediate immune evasion. MBio 2014, 5, e01974-14. [Google Scholar] [CrossRef] [Green Version]
- Diderrich, R.; Kock, M.; Maestre-Reyna, M.; Keller, P.; Steuber, H.; Rupp, S.; Essen, L.O.; Mösch, H.U. Structural Hot Spots Determine Functional Diversity of the Candida glabrata Epithelial Adhesin Family. J. Biol. Chem. 2015, 290, 19597–19613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, S.; Tchesnokova, V.; McVeigh, A.; Kisiela, D.I.; Dori, K.; Navarro, A.; Sokurenko, E.V.; Savarino, S.J. Adaptive evolution of class 5 fimbrial genes in enterotoxigenic Escherichia coli and its functional consequences. J. Biol. Chem. 2012, 287, 6150–6158. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, S.C.; Monday, S.R.; Hoffmann, M.; Fischer, M.; Kase, J.A. Plasmids from Shiga Toxin-Producing Escherichia coli Strains with Rare Enterohemolysin Gene (ehxA) Subtypes Reveal Pathogenicity Potential and Display a Novel Evolutionary Path. Appl. Environ. Microbiol. 2016, 82, 6367–6377. [Google Scholar] [CrossRef] [Green Version]
- Sjöholm, K.; Kilsgård, O.; Teleman, J.; Happonen, L.; Malmström, L.; Malmström, J. Targeted Proteomics and Absolute Protein Quantification for the Construction of a Stoichiometric Host-Pathogen Surface Density Model. Mol. Cell. Proteom. 2017, 16, S29–S41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, C.; Malmström, L.; Aebersold, R.; Malmström, J. Proteome-wide selected reaction monitoring assays for the human pathogen Streptococcus pyogenes. Nat. Commun. 2012, 3, 1301. [Google Scholar] [CrossRef]
- Karlsson, C.A.Q.; Järnum, S.; Winstedt, L.; Kjellman, C.; Björck, L.; Linder, A.; Malmström, J.A. Streptococcus pyogenes Infection and the Human Proteome with a Special Focus on the Immunoglobulin G-cleaving Enzyme IdeS. Mol. Cell. Proteom. 2018, 17, 1097–1111. [Google Scholar] [CrossRef] [Green Version]
- Lawson, C.L.; Chiu, W. Comparing cryo-EM structures. J. Struct. Biol. 2018, 204, 523–526. [Google Scholar] [CrossRef]
- Earl, L.A.; Falconieri, V.; Subramaniam, S. Microbiology catches the cryo-EM bug. Curr. Opin. Microbiol. 2018, 43, 199–207. [Google Scholar] [CrossRef]
- Hauri, S.; Khakzad, H.; Happonen, L.; Teleman, J.; Malmström, J.; Malmström, L. Rapid determination of quaternary protein structures in complex biological samples. Nat. Commun. 2019, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Bastos, V.A.; Gomes-Neto, F.; Rocha, S.L.G.; Teixeira-Ferreira, A.; Perales, J.; Neves-Ferreira, A.G.C.; Valente, R.H. The interaction between the natural metalloendopeptidase inhibitor BJ46a and its target toxin jararhagin analyzed by structural mass spectrometry and molecular modeling. J. Proteom. 2020, 221, 103761. [Google Scholar] [CrossRef] [PubMed]
- Slavata, L.; Chmelik, J.; Kavan, D.; Filandrová, R.; Fiala, J.; Rosůlek, M.; Mrázek, H.; Kukačka, Z.; Vališ, K.; Man, P.; et al. MS-Based Approaches Enable the Structural Characterization of Transcription Factor/DNA Response Element Complex. Biomolecules 2019, 9, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seacrist, C.D.; Kuenze, G.; Hoffmann, R.M.; Moeller, B.E.; Burke, J.E.; Meiler, J.; Blind, R.D. Integrated Structural Modeling of Full-Length LRH-1 Reveals Inter-domain Interactions Contribute to Receptor Structure and Function. Structure 2020, 28, 830–846.e9. [Google Scholar] [CrossRef]
- Zhang, M.M.; Beno, B.R.; Huang, R.Y.; Adhikari, J.; Deyanova, E.G.; Li, J.; Chen, G.; Gross, M.L. An Integrated Approach for Determining a Protein-Protein Binding Interface in Solution and an Evaluation of Hydrogen-Deuterium Exchange Kinetics for Adjudicating Candidate Docking Models. Anal. Chem. 2019, 91, 15709–15717. [Google Scholar] [CrossRef] [PubMed]
- López-Castilla, A.; Thomassin, J.L.; Bardiaux, B.; Zheng, W.; Nivaskumar, M.; Yu, X.; Nilges, M.; Egelman, E.H.; Izadi-Pruneyre, N.; Francetic, O. Structure of the calcium-dependent type 2 secretion pseudopilus. Nat. Microbiol. 2017, 2, 1686–1695. [Google Scholar] [CrossRef] [Green Version]
- Bardiaux, B.; de Amorim, G.C.; Luna Rico, A.; Zheng, W.; Guilvout, I.; Jollivet, C.; Nilges, M.; Egelman, E.H.; Izadi-Pruneyre, N.; Francetic, O. Structure and Assembly of the Enterohemorrhagic Escherichia. Coli. Type 4 Pilus. Structure 2019, 27, 1082–1093.e5. [Google Scholar] [CrossRef]
- Bardiaux, B.; Cordier, F.; Brier, S.; López-Castilla, A.; Izadi-Pruneyre, N.; Nilges, M. Dynamics of a type 2 secretion system pseudopilus unraveled by complementary approaches. J. Biomol. NMR 2019, 73, 293–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patwardhan, A.; Ashton, A.; Brandt, R.; Butcher, S.; Carzaniga, R.; Chiu, W.; Collinson, L.; Doux, P.; Duke, E.; Ellisman, M.H.; et al. A 3D cellular context for the macromolecular world. Nat. Struct. Mol. Biol. 2014, 21, 841–845. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Bardiaux, B.; Ahmed, M.; Spehr, J.; König, R.; Lünsdorf, H.; Rand, U.; Lührs, T.; Ritter, C. Structure determination of helical filaments by solid-state NMR spectroscopy. Proc. Natl. Acad. Sci. USA 2016, 113, E272–E281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferber, M.; Kosinski, J.; Ori, A.; Rashid, U.J.; Moreno-Morcillo, M.; Simon, B.; Bouvier, G.; Batista, P.R.; Müller, C.W.; Beck, M.; et al. Automated structure modeling of large protein assemblies using crosslinks as distance restraints. Nat. Methods 2016, 13, 515–520. [Google Scholar] [CrossRef]
- Ramboarina, S.; Garnett, J.A.; Zhou, M.; Li, Y.; Peng, Z.; Taylor, J.D.; Lee, W.C.; Bodey, A.; Murray, J.W.; Alguel, Y.; et al. Structural insights into serine-rich fimbriae from Gram-positive bacteria. J. Biol. Chem. 2010, 285, 32446–32457. [Google Scholar] [CrossRef] [Green Version]
- Olano, J.P.; Walker, D.H. Diagnosing emerging and reemerging infectious diseases: The pivotal role of the pathologist. Arch. Pathol. Lab. Med. 2011, 135, 83–91. [Google Scholar] [CrossRef]
- Ramanan, P.; Bryson, A.L.; Binnicker, M.J.; Pritt, B.S.; Patel, R. Syndromic Panel-Based Testing in Clinical Microbiology. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özenci, V.; Patel, R.; Ullberg, M.; Strålin, K. Demise of Polymerase Chain Reaction/Electrospray Ionization-Mass Spectrometry as an Infectious Diseases Diagnostic Tool. Clin. Infect. Dis. 2018, 66, 452–455. [Google Scholar] [CrossRef]
- Campos, M.; Francetic, O.; Nilges, M. Modeling pilus structures from sparse data. J. Struct. Biol. 2011, 173, 436–444. [Google Scholar] [CrossRef]
- Schmid, Y.; Grassl, G.A.; Buhler, O.T.; Skurnik, M.; Autenrieth, I.B.; Bohn, E. Yersinia enterocolitica adhesin A induces production of interleukin-8 in epithelial cells. Infect. Immun. 2004, 72, 6780–6789. [Google Scholar] [CrossRef] [Green Version]
- Leo, J.C.; Goldman, A. The immunoglobulin-binding Eib proteins from Escherichia coli are receptors for IgG Fc. Mol. Immunol. 2009, 46, 1860–1866. [Google Scholar] [CrossRef]
- Leo, J.C.; Łyskowski, A.; Hattula, K.; Hartmann, M.D.; Schwarz, H.; Butcher, S.J.; Linke, D.; Lupas, A.N.; Goldman, A. The structure of E. coli IgG-binding protein D suggests a general model for bending and binding in trimeric autotransporter adhesins. Structure 2011, 19, 1021–1030. [Google Scholar] [CrossRef] [Green Version]
- Kearney, K.J.; Pechlivani, N.; King, R.; Tiede, C.; Phoenix, F.; Cheah, R.; Macrae, F.L.; Simmons, K.J.; Manfield, I.W.; Smith, K.A.; et al. Affimer proteins as a tool to modulate fibrinolysis, stabilize the blood clot, and reduce bleeding complications. Blood 2019, 133, 1233–1244. [Google Scholar] [CrossRef] [Green Version]
- Tiede, C.; Tang, A.A.; Deacon, S.E.; Mandal, U.; Nettleship, J.E.; Owen, R.L.; George, S.E.; Harrison, D.J.; Owens, R.J.; Tomlinson, D.C.; et al. Adhiron: A stable and versatile peptide display scaffold for molecular recognition applications. Protein Eng. Des. Sel. 2014, 27, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Peña-Bahamonde, J.; Nguyen, H.N.; Fanourakis, S.K.; Rodrigues, D.F. Recent advances in graphene-based biosensor technology with applications in life sciences. J. Nanobiotechnol. 2018, 16, 75. [Google Scholar] [CrossRef] [Green Version]
- Elahi, N.; Kamali, M.; Baghersad, M.H.; Amini, B. A fluorescence Nano-biosensors immobilization on Iron (MNPs) and gold (AuNPs) nanoparticles for detection of Shigella spp. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 105, 110113. [Google Scholar] [CrossRef]
- Govender, V.S.; Ramsugit, S.; Pillay, M. Mycobacterium tuberculosis adhesins: Potential biomarkers as anti-tuberculosis therapeutic and diagnostic targets. Microbiology 2014, 160, 1821–1831. [Google Scholar] [CrossRef] [Green Version]
- Peeling, R.W.; Mabey, D. Point-of-care tests for diagnosing infections in the developing world. Clin. Microbiol. Infect. 2010, 16, 1062–1069. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Zhou, L.; He, D.; Wang, K.; Cao, J. Rapid and ultrasensitive E. coli O157:H7 quantitation by combination of ligandmagnetic nanoparticles enrichment with fluorescent nanoparticles based two-color flow cytometry. Analyst 2011, 136, 4183–4191. [Google Scholar] [CrossRef]
- Kretzer, J.W.; Lehmann, R.; Schmelcher, M.; Banz, M.; Kim, K.P.; Korn, C.; Loessner, M.J. Use of high-affinity cell wall-binding domains of bacteriophage endolysins for immobilization and separation of bacterial cells. Appl. Environ. Microbiol. 2007, 73, 1992–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilecky, M.; Schildberger, A.; Orth-Höller, D.; Weber, V. Pathogen enrichment from human whole blood for the diagnosis of bloodstream infection: Prospects and limitations. Diagn. Microbiol. Infect. Dis. 2019, 94, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Auriti, C.; Prencipe, G.; Moriondo, M.; Bersani, I.; Bertaina, C.; Mondì, V.; Inglese, R. Mannose-Binding Lectin: Biologic Characteristics and Role in the Susceptibility to Infections and Ischemia-Reperfusion Related Injury in Critically Ill Neonates. J. Immunol. Res. 2017, 2017, 7045630. [Google Scholar] [CrossRef] [PubMed]
- Unione, L.; Gimeno, A.; Valverde, P.; Calloni, I.; Coelho, H.; Mirabella, S.; Poveda, A.; Arda, A.; Jimenez-Barbero, J. Glycans in Infectious Diseases. A Molecular Recognition Perspective. Curr. Med. Chem. 2017, 24, 4057–4080. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.W. The role of mannose-binding lectin in health and disease. Mol. Immunol. 2003, 40, 423–429. [Google Scholar] [CrossRef]
- Kangwa, M.; Yelemane, V.; Ponnurangam, A.; Fernández-Lahore, M. An engineered Staphylococcal Protein A based ligand: Production, characterization and potential application for the capture of Immunoglobulin and Fc-fusion proteins. Protein Expr. Purif. 2019, 155, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Templier, V.; Roux, A.; Roupioz, Y.; Livache, T. Ligands for label-free detection of whole bacteria on biosensors: A review. Trac-Trend. Anal. Chem. 2016, 79, 71–79. [Google Scholar] [CrossRef]
- Dunne, M.; Loessner, M.J. Modified Bacteriophage Tail Fiber Proteins for Labeling, Immobilization, Capture, and Detection of Bacteria. Methods Mol. Biol. 2019, 1918, 67–86. [Google Scholar] [CrossRef]
- Bicart-See, A.; Rottman, M.; Cartwright, M.; Seiler, B.; Gamini, N.; Rodas, M.; Penary, M.; Giordano, G.; Oswald, E.; Super, M.; et al. Rapid Isolation of Staphylococcus aureus Pathogens from Infected Clinical Samples Using Magnetic Beads Coated with Fc-Mannose Binding Lectin. PLoS ONE 2016, 11, e0156287. [Google Scholar] [CrossRef]
- Wang, J.; Wu, H.; Yang, Y.; Yan, R.; Zhao, Y.; Wang, Y.; Chen, A.; Shao, S.; Jiang, P.; Li, Y.Q. Bacterial species-identifiable magnetic nanosystems for early sepsis diagnosis and extracorporeal photodynamic blood disinfection. Nanoscale 2017, 10, 132–141. [Google Scholar] [CrossRef]
- Jung, S.H.; Hahn, Y.K.; Oh, S.; Kwon, S.; Um, E.; Choi, S.; Kang, J.H. Advection Flows-Enhanced Magnetic Separation for High-Throughput Bacteria Separation from Undiluted Whole Blood. Small 2018, 14, e1801731. [Google Scholar] [CrossRef]
- Zheng, L.; Wan, Y.; Qi, P.; Sun, Y.; Zhang, D.; Yu, L. Lectin functionalized ZnO nanoarrays as a 3D nano-biointerface for bacterial detection. Talanta 2017, 167, 600–606. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhu, B.; Li, Y.; Leow, W.R.; Goh, R.; Ma, B.; Fong, E.; Tang, M.; Chen, X. A synergistic capture strategy for enhanced detection and elimination of bacteria. Angew. Chem. Int. Ed. Engl. 2014, 53, 5837–5841. [Google Scholar] [CrossRef]
- Liu, L.; Chen, S.; Xue, Z.; Zhang, Z.; Qiao, X.; Nie, Z.; Han, D.; Wang, J.; Wang, T. Bacterial capture efficiency in fluid bloodstream improved by bendable nanowires. Nat. Commun. 2018, 9, 444. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Ma, Y.; Pan, J.; Lee, S.H.; Liu, J.; Zhu, H.; Gu, R.; Shea, K.J.; Pan, G. Efficient capture, rapid killing and ultrasensitive detection of bacteria by a nano-decorated multi-functional electrode sensor. Biosens. Bioelectron. 2018, 101, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Sheng, E.Z.; Lu, Y.X.; Xiao, Y.; Li, Z.X.; Wang, H.S.; Dai, Z.H. Simultaneous and ultrasensitive detection of three pesticides using a surface-enhanced Raman scattering-based lateral flow assay test strip. Biosens. Bioelectron. 2021, 181. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Wang, Y.; Zhang, D. An integrated multifunctional photoelectrochemical platform for simultaneous capture, detection, and inactivation of pathogenic bacteria. Sensor. Actuat. B-Chem. 2018, 274, 228–234. [Google Scholar] [CrossRef]
- Dao, T.N.T.; Lee, E.Y.; Koo, B.; Jin, C.E.; Lee, T.Y.; Shin, Y. A microfluidic enrichment platform with a recombinase polymerase amplification sensor for pathogen diagnosis. Anal. Biochem. 2018, 544, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, P.; Petersson, K.; Augustsson, P.; Laurell, T. Acoustic impedance matched buffers enable separation of bacteria from blood cells at high cell concentrations. Sci. Rep. 2018, 8, 9156. [Google Scholar] [CrossRef]
- Dow, P.; Kotz, K.; Gruszka, S.; Holder, J.; Fiering, J. Acoustic separation in plastic microfluidics for rapid detection of bacteria in blood using engineered bacteriophage. Lab. Chip. 2018, 18, 923–932. [Google Scholar] [CrossRef]
- Olanrewaju, A.O.; Ng, A.; DeCorwin-Martin, P.; Robillard, A.; Juncker, D. Microfluidic Capillaric Circuit for Rapid and Facile Bacteria Detection. Anal. Chem. 2017, 89, 6846–6853. [Google Scholar] [CrossRef] [Green Version]
- AbdElFatah, T.; Jalali, M.; Mahshid, S. A nanosurface microfluidic device for capture and detection of bacteria. In Proceedings of the IEEE Life Sciences Conference (LSC), Montreal, QC, Canada, 28–30 October 2018; pp. 167–170. [Google Scholar]
- Qu, Y.; Wei, T.; Zhan, W.; Hu, C.; Cao, L.; Yu, Q.; Chen, H. A reusable supramolecular platform for the specific capture and release of proteins and bacteria. J. Mater. Chem B 2017, 5, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.; Lee, S.H.; Song, J.; Bhattacharjee, S.; Feng, J.; Hong, S.; Song, M.; Kim, W.; Lee, J.; Bang, D.; et al. Nanophotonic Cell Lysis and Polymerase Chain Reaction with Gravity-Driven Cell Enrichment for Rapid Detection of Pathogens. ACS. Nano 2019, 13, 13866–13874. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.B.; Qi, P.; Zhang, D. DNA-templated fluorescent silver nanoclusters for sensitive detection of pathogenic bacteria based on MNP-DNAzyme-AChE complex. Sensor. Actuat. B-Chem. 2018, 276, 42–47. [Google Scholar] [CrossRef]
- Zeltzer, P.M.; Seeger, R.C. Microassay using radioiodinated protein A from Staphylococcus aureus for antibodies bound to cell surface antigens of adherent tumor cells. J. Immunol. Methods 1977, 17, 163–175. [Google Scholar] [CrossRef]
- Schuler, G.; Auböck, J.; Linert, J. Enrichment of epidermal Langerhans cells by immunoadsorption to Staphylococcus aureus cells. J. Immunol. 1983, 130, 2008–2010. [Google Scholar]
- Fishman, J.B.; Berg, E.A. Protein A and Protein G Purification of Antibodies. Cold. Spring. Harb. Protoc. 2019, 2019. [Google Scholar] [CrossRef]
- Rigi, G.; Ghaedmohammadi, S.; Ahmadian, G. A comprehensive review on staphylococcal protein A (SpA): Its production and applications. Biotechnol. Appl. Biochem. 2019, 66, 454–464. [Google Scholar] [CrossRef]
- Cronin, U.P.; Girardeaux, L.; O’Meara, E.; Wilkinson, M.G. Protein A-Mediated Binding of Staphylococcus spp. to Antibodies in Flow Cytometric Assays and Reduction of This Binding by Using Fc Receptor Blocking Reagent. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]
- Sandin, C.; Linse, S.; Areschoug, T.; Woof, J.M.; Reinholdt, J.; Lindahl, G. Isolation and detection of human IgA using a streptococcal IgA-binding peptide. J. Immunol. 2002, 169, 1357–1364. [Google Scholar] [CrossRef] [Green Version]
- Gullsby, K.; Olsen, B.; Bondeson, K. Molecular Typing of Mycoplasma pneumoniae Strains in Sweden from 1996 to 2017 and the Emergence of a New P1 Cytadhesin Gene, Variant 2e. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Röst, H.L.; Rosenberger, G.; Navarro, P.; Gillet, L.; Miladinović, S.M.; Schubert, O.T.; Wolski, W.; Collins, B.C.; Malmström, J.; Malmström, L.; et al. OpenSWATH enables automated, targeted analysis of data-independent acquisition MS data. Nat. Biotechnol. 2014, 32, 219–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, S.; Wang, K.; Li, Z.; Wang, C.; Hao, Z.; Liu, W.; Wan, J. An electrochemical biosensor based on methylene blue-loaded nanocomposites as signal-amplifying tags to detect pathogenic bacteria. Analyst 2020, 145, 4328–4334. [Google Scholar] [CrossRef] [PubMed]
- Cesewski, E.; Johnson, B.N. Electrochemical biosensors for pathogen detection. Biosens. Bioelectron. 2020, 159, 112214. [Google Scholar] [CrossRef]
- Zhang, Y.; Das, G.K.; Vijayaragavan, V.; Xu, Q.C.; Padmanabhan, P.; Bhakoo, K.K.; Selvan, S.T.; Tan, T.T. “Smart” theranostic lanthanide nanoprobes with simultaneous up-conversion fluorescence and tunable T1-T2 magnetic resonance imaging contrast and near-infrared activated photodynamic therapy. Nanoscale 2014, 6, 12609–12617. [Google Scholar] [CrossRef] [Green Version]
- Erickson, D.; Mandal, S.; Yang, A.H.; Cordovez, B. Nanobiosensors: Optofluidic, electrical and mechanical approaches to biomolecular detection at the nanoscale. Microfluid. Nanofluid. 2008, 4, 33–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okshevsky, M.; Meyer, R.L. The role of extracellular DNA in the establishment, maintenance and perpetuation of bacterial biofilms. Crit. Rev. Microbiol. 2015, 41, 341–352. [Google Scholar] [CrossRef]
- Metkar, S.K.; Girigoswami, K. Diagnostic biosensors in medicine—A review. Biocatal. Agr. Biotech. 2019, 17, 271–283. [Google Scholar] [CrossRef]
- Taleat, Z.; Khoshroo, A.; Mazloum-Ardakani, M. Screen-printed electrodes for biosensing: A review (2008–2013). Microchim. Acta 2014, 181, 865–891. [Google Scholar] [CrossRef]
- Mairhofer, J.; Roppert, K.; Ertl, P. Microfluidic systems for pathogen sensing: A review. Sensors 2009, 9, 4804–4823. [Google Scholar] [CrossRef]
- Birch, J.R.; Racher, A.J. Antibody production. Adv. Drug. Deliv. Rev. 2006, 58, 671–685. [Google Scholar] [CrossRef]
- Leva-Bueno, J.; Peyman, S.A.; Millner, P.A. A review on impedimetric immunosensors for pathogen and biomarker detection. Med. Microbiol. Immunol. 2020, 209, 343–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzen, T.R.J.; Cheng, Y.Y.R.; Saeidpourazar, R.; Aphale, S.S.; Jalili, N. Adhesin-Specific Nanomechanical Cantilever Biosensors for Detection of Microorganisms. J. Heat. Trans-T Asme. 2011, 133. [Google Scholar] [CrossRef]
- Medina, M.B. Biosensor studies of the binding of extracellular matrix components with immobilized Escherichia coli O157: H7 and inhibition by polysulfated polysaccharides. Biotechnol. Lett. 2002, 24, 77–84. [Google Scholar] [CrossRef]
- Jantra, J.; Kanatharana, P.; Asawatreratanakul, P.; Hedström, M.; Mattiasson, B.; Thavarungkul, P. Real-time label-free affinity biosensors for enumeration of total bacteria based on immobilized concanavalin A. J. Environ. Sci. Health Part A 2011, 46, 1450–1460. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Li, S.; Terebiznik, M.; Guyard, C.; Kerman, K. Biosensors for the Detection of Interaction between Legionella pneumophila Collagen-Like Protein and Glycosaminoglycans. Sensors 2018, 18, 668. [Google Scholar] [CrossRef] [Green Version]
- Hai, W.; Goda, T.; Takeuchi, H.; Yamaoka, S.; Horiguchi, Y.; Matsumoto, A.; Miyahara, Y. Specific Recognition of Human Influenza Virus with PEDOT Bearing Sialic Acid-Terminated Trisaccharides. ACS Appl. Mater. Interfaces 2017, 9, 14162–14170. [Google Scholar] [CrossRef]
- Shimetani, N. Potential of Next-generation POCT in Infectious Disease Rapid Test. Med. Mycol. J. 2017, 58, J91–J94. [Google Scholar] [CrossRef] [Green Version]
- Kretzer, J.W.; Schmelcher, M.; Loessner, M.J. Ultrasensitive and Fast Diagnostics of Viable Listeria Cells by CBD Magnetic Separation Combined with A511::luxAB Detection. Viruses 2018, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Saab, M.E.; Muckle, C.A.; Stryhn, H.; McClure, J.T. Comparison of culture methodology for the detection of methicillin-resistant Staphylococcus pseudintermedius in clinical specimens collected from dogs. J. Vet. Diagn. Investig. 2018, 30, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burtscher, S.; Krieg, P.; Killinger, A.; Al-Ahmad, A.; Seidenstücker, M.; Latorre, S.H.; Bernstein, A. Thin Degradable Coatings for Optimization of Osteointegration Associated with Simultaneous Infection Prophylaxis. Materials 2019, 12, 3495. [Google Scholar] [CrossRef] [Green Version]
- Boardman, A.K.; Allison, S.; Sharon, A.; Sauer-Budge, A.F. Comparison of anti-fouling surface coatings for applications in bacteremia diagnostics. Anal. Methods 2013, 5, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Andel, E.; de Bus, I.; Tijhaar, E.J.; Smulders, M.M.J.; Savelkoul, H.F.J.; Zuilhof, H. Highly Specific Binding on Antifouling Zwitterionic Polymer-Coated Microbeads as Measured by Flow Cytometry. ACS. Appl. Mater. Interfaces 2017, 9, 38211–38221. [Google Scholar] [CrossRef]
- Swar, S.; Máková, V.; Stibor, I. Effectiveness of Diverse Mesoporous Silica Nanoparticles as Potent Vehicles for the Drug L-DOPA. Materials 2019, 12, 202. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.R.E.; Mendonca, P.V.; Almeida, M.C.; Branco, R.; Serra, A.C.; Morais, P.V.; Coelho, J.F.J. Increasing the Antimicrobial Activity of Amphiphilic Cationic Copolymers by the Facile Synthesis of High Molecular Weight Stars by Supplemental Activator and Reducing Agent Atom Transfer Radical Polymerization. Biomacromolecules 2019, 20, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Braun, B.M.; Skelly, J.D.; Ayers, D.C.; Song, J. Significant Suppression of Staphylococcus aureus Colonization on Intramedullary Ti6Al4V Implants Surface-Grafted with Vancomycin-Bearing Polymer Brushes. ACS. Appl. Mater. Interfaces 2019, 11, 28641–28647. [Google Scholar] [CrossRef]
- Perez-Alvarez, L.; Ruiz-Rubio, L.; Azua, I.; Benito, V.; Bilbao, A.; Vilas-Vilela, J.L. Development of multiactive antibacterial multilayers of hyaluronic acid and chitosan onto poly(ethylene terephthalate). Eur. Polym. J. 2019, 112, 31–37. [Google Scholar] [CrossRef]
- Arisoy, F.D.; Kolewe, K.W.; Homyak, B.; Kurtz, I.S.; Schiffman, J.D.; Watkins, J.J. Bioinspired Photocatalytic Shark-Skin Surfaces with Antibacterial and Antifouling Activity via Nanoimprint Lithography. Acs. Appl. Mater. Inter. 2018, 10, 20055–20063. [Google Scholar] [CrossRef] [PubMed]
- Ceresa, C.; Tessarolo, F.; Maniglio, D.; Tambone, E.; Carmagnola, I.; Fedeli, E.; Caola, I.; Nollo, G.; Chiono, V.; Allegrone, G.; et al. Medical-Grade Silicone Coated with Rhamnolipid R89 Is Effective against Staphylococcus spp. Biofilms. Molecules 2019, 24, 3843. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Willing, B.; Bjerketorp, J.; Jansson, J.K.; Hjort, K. Soft inertial microfluidics for high throughput separation of bacteria from human blood cells. Lab. Chip. 2009, 9, 1193–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, E.D.; Huang, C.; Hawkins, B.G.; Gleghorn, J.P.; Kirby, B.J. Rare Cell Capture in Microfluidic Devices. Chem. Eng. Sci. 2011, 66, 1508–1522. [Google Scholar] [CrossRef] [Green Version]
- Shangguan, J.; Li, Y.; He, D.; He, X.; Wang, K.; Zou, Z.; Shi, H. A combination of positive dielectrophoresis driven on-line enrichment and aptamer-fluorescent silica nanoparticle label for rapid and sensitive detection of Staphylococcus aureus. Analyst 2015, 140, 4489–4497. [Google Scholar] [CrossRef]
- Pinkner, J.S.; Remaut, H.; Buelens, F.; Miller, E.; Åberg, V.; Pemberton, N.; Hedenström, M.; Larsson, A.; Seed, P.; Waksman, G.; et al. Rationally designed small compounds inhibit pilus biogenesis in uropathogenic bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 17897–17902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qvortrup, K.; Hultqvist, L.D.; Nilsson, M.; Jakobsen, T.H.; Jansen, C.U.; Uhd, J.; Andersen, J.B.; Nielsen, T.E.; Givskov, M.; Tolker-Nielsen, T. Small Molecule Anti-biofilm Agents Developed on the Basis of Mechanistic Understanding of Biofilm Formation. Front. Chem. 2019, 7, 742. [Google Scholar] [CrossRef] [Green Version]
- Zalewska-Piątek, B.M.; Piątek, R.J. Alternative treatment approaches of urinary tract infections caused by uropathogenic Escherichia coli strains. Acta Biochim. Pol. 2019, 66, 129–138. [Google Scholar] [CrossRef]
- Chorell, E.; Pinkner, J.S.; Bengtsson, C.; Banchelin, T.S.; Edvinsson, S.; Linusson, A.; Hultgren, S.J.; Almqvist, F. Mapping pilicide anti-virulence effect in Escherichia coli, a comprehensive structure-activity study. Bioorg. Med. Chem. 2012, 20, 3128–3142. [Google Scholar] [CrossRef] [Green Version]
- Drancourt, M.; Michel-Lepage, A.; Boyer, S.; Raoult, D. The Point-of-Care Laboratory in Clinical Microbiology. Clin. Microbiol. Rev. 2016, 29, 429–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Z.; Guo, J.; Xiao, Y.; He, Z.; Xia, X.; Huang, Z.; Guan, H.; Ling, X.; Li, J.; Diao, B.; et al. Comparison of BioFire FilmArray gastrointestinal panel versus Luminex xTAG Gastrointestinal Pathogen Panel (xTAG GPP) for diarrheal pathogen detection in China. Int. J. Infect. Dis. 2020, 99, 414–420. [Google Scholar] [CrossRef]
- Buchan, B.W.; Windham, S.; Balada-Llasat, J.M.; Leber, A.; Harrington, A.; Relich, R.; Murphy, C.; Dien Bard, J.; Naccache, S.; Ronen, S.; et al. Practical Comparison of the BioFire FilmArray Pneumonia Panel to Routine Diagnostic Methods and Potential Impact on Antimicrobial Stewardship in Adult Hospitalized Patients with Lower Respiratory Tract Infections. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Happonen, L.; Khakzad, H.; Malmström, L.; Malmström, J. Structural proteomics, electron cryo-microscopy and structural modeling approaches in bacteria-human protein interactions. Med. Microbiol. Immunol. 2020, 209, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Galvan, D.D.; Parekh, V.; Liu, E.; Liu, E.L.; Yu, Q. Sensitive Bacterial Detection via Dielectrophoretic-Enhanced Mass Transport Using Surface-Plasmon-Resonance Biosensors. Anal. Chem. 2018, 90, 14635–14642. [Google Scholar] [CrossRef] [PubMed]
- Prozeller, D.; Morsbach, S.; Landfester, K. Isothermal titration calorimetry as a complementary method for investigating nanoparticle-protein interactions. Nanoscale 2019, 11, 19265–19273. [Google Scholar] [CrossRef] [Green Version]
- Niether, D.; Wiegand, S. Thermophoresis of biological and biocompatible compounds in aqueous solution. J. Phys. Condens. Matter 2019, 31, 503003. [Google Scholar] [CrossRef] [PubMed]
- Philo, J.S. Is any measurement method optimal for all aggregate sizes and types? AAPS. J. 2006, 8, E564–E571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vankadari, N.; Wilce, J.A. Emerging WuHan (COVID-19) coronavirus: Glycan shield and structure prediction of spike glycoprotein and its interaction with human CD26. Emerg. Microbes. Infect. 2020, 9, 601–604. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Priority 1: Critical | |
| Acinetobacter baumannii | carbapenem-resistant |
| Pseudomonas aeruginosa | carbapenem-resistant |
| Enterobacteriales | carbapenem-resistant, ESBL-producing |
| Priority 2: High | |
| Enterococcus faecium | vancomycin-resistant |
| Staphylococcus aureus | methicillin-resistant, vancomycin-intermediate and resistant |
| Helicobacter pylori | clarithromycin-resistant |
| Campylobacter spp. | fluoroquinolone-resistant |
| Salmonellae | fluoroquinolone-resistant |
| Neisseria gonorrhoeae | cephalosporin-resistant, fluoroquinolone-resistant |
| Priority 3: Medium | |
| Streptococcus pneumoniae | penicillin-non-susceptible |
| Haemophilus influenzae | ampicillin-resistant |
| Shigella spp. | fluoroquinolone-resistant |
| Device | Enrichment Technique | Target | Detection Method | Limit of Detection | Time | Reference |
|---|---|---|---|---|---|---|
| 3D printed microfluidic biosensor | Aptamer coated magnetic beads with magnetic separation | Plasmodium falciparum lactate dehydrogenase (PfLDH) enzyme | Colorimetric | Parasitemia < 0.01% | 180 min | [109] |
| Enzyme-linked LTF assay (ELLTA) | Long tail fibers (S16 LTF) of bacteriophages immobilized onto paramagnetic beads | Salmonella typhimurium | Colorimetric | 102 cfu/mL | 2 h | [110] |
| Assay | Magnetic beads coated with the engineered chimeric human opsonin protein, Fc-mannose-binding lectin (FcMBL) | Articular fluid samples and synovial tissue samples from patients with S. aureus infections | RT-PCR analysis and MALDI-TOF | 76% ± 5.7% capture efficiency | - | [111] |
| Assay | Iron oxide magnetic nanoparticles functionalized with bacterial species-identifiable aptamers | S. aureus and E. coli | Fluorescence microscopy | 10 CFU | 1.5 h | [112] |
| Microfluidic platform | Induced advectivespiral flows of super-paramagnetic nanoparticles coated with mannose-binding lectin and magnetic separation | E. coli spiked into undiluted rat whole blood | None | 91.68% ± 2.18% capture efficiency | - | [113] |
| 3D Nano-biointerface platform | Zinc oxide nanorod array 3D nano–bio surface functionalized with lectin Concanavalin A | E. coli | Fluorescence microscopy imaging | 0.9 × 102 CFU/mL | - | [114] |
| Nanowire arrays | Functionalized 3D nanowire substrate | S. aureus | Fluorescence microscopy | 10 CFU/mL | 30 min | [115] |
| Nanowire arrays | Bendable polycrystalline nanowires pre-grafted on 3D carbon foam | Human blood spiked with Salmonella spp | Fluorescence microscopy | ~97% capture efficiency | - | [116] |
| Impedanceelectrode sensor | Antibacterial prickly Zn-CuO nanoparticles with burr-like nanostructures | Rat blood spiked with E. coli | Impedance-based electrode sensor | 10 CFU/mL | 20 min | [117] |
| Surface-Enhanced Raman Scattering Multi-Multifunction Chip | 4-mercaptophenylboronic acid | Humanblood spiked with E.coli, S. aureus | Surface-Enhanced Raman Scattering | 1.0 × 102 cells m/L | - | [118] |
| Photoelectrochemical platform | 4-mercaptophenylboronic acid | E. coli | Photoelectrode | 46 CFU/mL | 30 min | [119] |
| Microfluidic platform | Magainin 1 peptide | urine spiked with Salmonella spp; Brucella spp | Recombinase polymerase amplification (RPA) sensor | 5 CFU/mL urine for Salmonella; 10 CFU/mL for Brucella | 60 min | [120] |
| Microfluidic chip | Bulk acoustophoresis | diluted whole blood spiked with Pseudomonas putida | Microscopy | - | 12.5 min | [121] |
| Microfluidic chip | Bulk acoustophoresis | Pseudomonas aeruginosa, S. aureus, E. coli | Luminescent bacterio-phage assay | 45% to 60% capture efficiency | - | [122] |
| Microfluidic capillaric circuit | Antibody-functionalized microbeads | synthetic urine spiked with E. coli | Fluorescence microscopy | 1.2 × 102 CFU/mL | 7 min | [123] |
| Microfluidic chip | Pillar-assisted self-assembly microparticles Nano- filter for | E. coli from samples | Fluorescence microscopy | capture efficiency of 93% | - | [124] |
| Reusable supramolecular platform | Multilayered film and β-cyclodextrin (β-CD) derivatives modified with mannose | Type I fimbriae E. coli and lectin proteins | Fluorescence microscopy | Capture efficiency of 93% | - | [125] |
| Photonic PCR on a chip | Gravity-driven cell enrichment | E. coli | Photonic PCR on a chip | 103 CFU/mL | 10 min | [126] |
| Enzyme-linked lectin sorbent assay (ELLecSA) | Fc-mannose-binding lectin | Bacteria, fungi, virus, parasites. LPS, LTA from Gram-negative and Gram-positive bacteria, as well as lipo-arabino-mannan (LAM) and phosphatidyl-inositol mannoside from M. tuberculosis | Scanning electron microscopy | - | <1 h | [114] |
| Fluorometric assay | Two distinct terminal phosphate-labeled LPS specific aptamers attached onto Zr-MOFs to fabricate the magnetic core-shell for magnetic separation | Acinetobacter baumannii in blood samples | Fluorescent signal amplification by fluorescence probes | 10 cfu/mL | ~2.5 h | [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Belkum, A.; Almeida, C.; Bardiaux, B.; Barrass, S.V.; Butcher, S.J.; Çaykara, T.; Chowdhury, S.; Datar, R.; Eastwood, I.; Goldman, A.; et al. Host-Pathogen Adhesion as the Basis of Innovative Diagnostics for Emerging Pathogens. Diagnostics 2021, 11, 1259. https://doi.org/10.3390/diagnostics11071259
van Belkum A, Almeida C, Bardiaux B, Barrass SV, Butcher SJ, Çaykara T, Chowdhury S, Datar R, Eastwood I, Goldman A, et al. Host-Pathogen Adhesion as the Basis of Innovative Diagnostics for Emerging Pathogens. Diagnostics. 2021; 11(7):1259. https://doi.org/10.3390/diagnostics11071259
Chicago/Turabian Stylevan Belkum, Alex, Carina Almeida, Benjamin Bardiaux, Sarah V. Barrass, Sarah J. Butcher, Tuğçe Çaykara, Sounak Chowdhury, Rucha Datar, Ian Eastwood, Adrian Goldman, and et al. 2021. "Host-Pathogen Adhesion as the Basis of Innovative Diagnostics for Emerging Pathogens" Diagnostics 11, no. 7: 1259. https://doi.org/10.3390/diagnostics11071259