
Correlation between Commercial Anti-RBD IgG Titer and Neutralization Titer against SARS-CoV-2 Beta Variant

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Specimens
2.2. Chemiluminescent Microparticle Immunoassay for the Detection of Anti-N and Anti-RBD IgG
2.3. Conventional Live Virus Neutralization Assay
2.4. Statistical Analysis
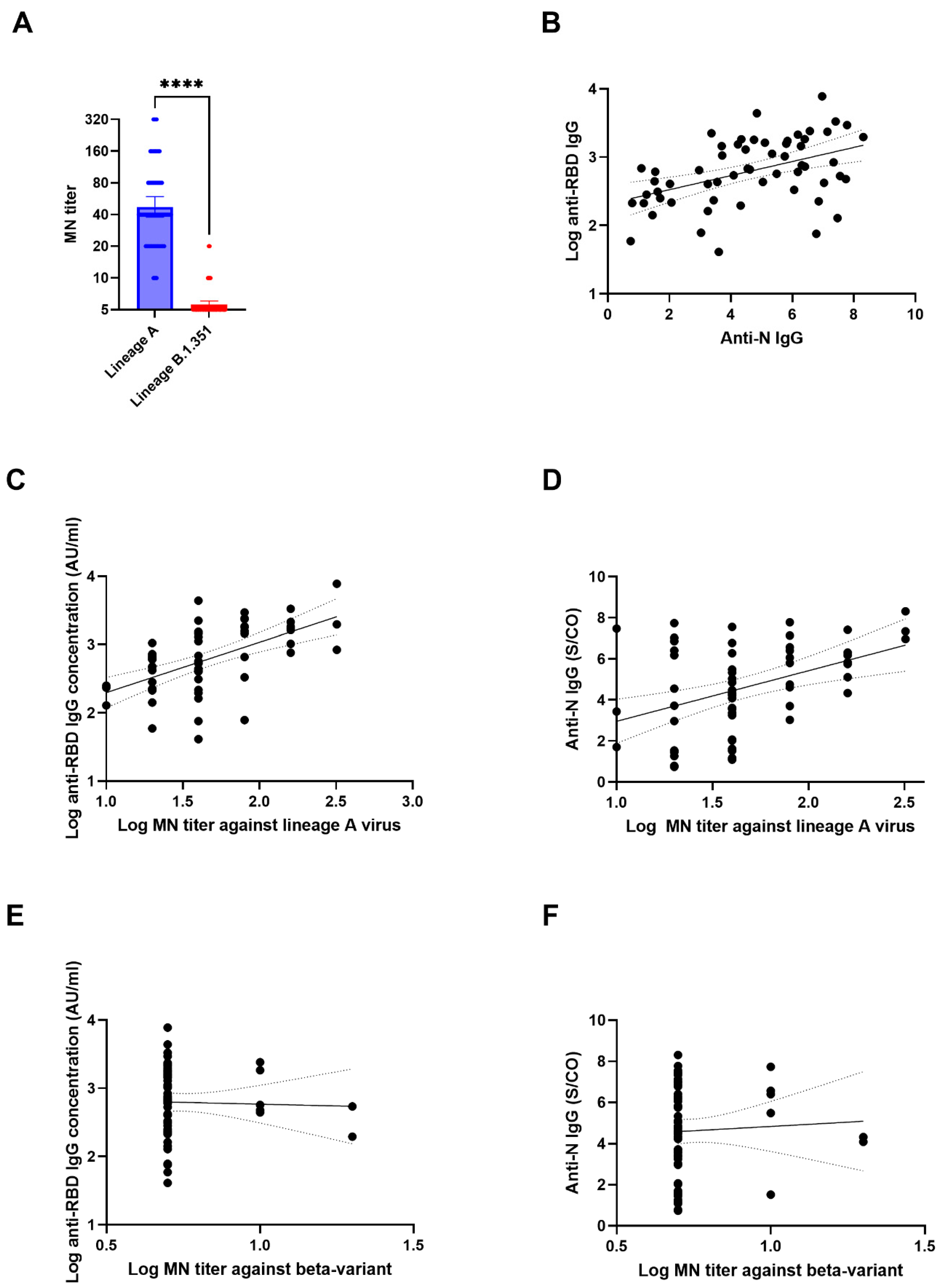
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ong, D.S.; Fragkou, P.C.; Schweitzer, V.A.; Chemaly, R.F.; Moschopoulos, C.D.; Skevaki, C. How to interpret and use COVID-19 serology and immunology tests. Clin. Microbiol. Infect. 2021, 27, 981–986. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.W.; Cheng, V.C.C.; Cai, J.P.; Chan, K.H.; Chen, L.L.; Wong, L.H.; Choi, C.Y.K.; Fong, C.H.Y.; Ng, A.C.K.; Lu, L.; et al. Seroprevalence of SARS-CoV-2 in Hong Kong special administrative region and our returnees evacuated from Hubei province of China: A multi-cohort study. Lancet Microbe 2020, 1, e111–e118. [Google Scholar] [CrossRef]
- To, K.K.W.; Hung, I.F.N.; Chan, K.H.; Yuan, S.; To, W.K.; Tsang, D.N.C.; Cheng, V.C.C.; Chen, Z.; Kok, K.H.; Yuen, K.Y. Serum antibody profile of a patient with Coronavirus disease 2019 reinfection. Clin. Infect. Dis. 2021, 72, e659–e662. [Google Scholar] [CrossRef]
- To, K.K.W.; Tsang, O.T.Y.; Leung, W.S.; Tam, A.R.; Wu, T.C.; Lung, D.C.; Yip, C.C.Y.; Cai, J.P.; Chan, J.M.C.; Chik, T.S.H.; et al. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2, An observational cohort study. Lancet Infect. Dis. 2020, 20, 565–574. [Google Scholar] [CrossRef] [Green Version]
- To, K.K.W.; Sridhar, S.; Chiu, K.H.Y.; Hung, D.L.L.; Li, X.; Hung, I.F.N.; Tam, A.R.; Chung, T.W.H.; Chan, J.F.W.; Zhang, A.J.X.; et al. Lessons learned one year after SARS-CoV-2 emergence leading to COVID-19 pandemic. Emerg. Microbes Infect. 2021, 10, 507–535. [Google Scholar] [CrossRef] [PubMed]
- Fong, C.H.Y.; Cai, J.P.; Dissanayake, T.K.; Chen, L.L.; Choi, C.Y.K.; Wong, L.H.; Ng, A.C.K.; Pang, P.K.; Ho, D.T.Y.; Poon, R.W.S.; et al. Improved detection of antibodies against SARS-CoV-2 by microsphere-based antibody assay. Int. J. Mol. Sci. 2020, 21, 6595. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.C.; Tiu, C.; Hu, Z.; Chen, V.C.W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ace2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; To, K.K.W.; Chan, K.H.; Wong, Y.C.; Zhou, R.; Kwan, K.Y.; Fong, C.H.Y.; Chen, L.L.; Choi, C.Y.K.; Lu, L.; et al. High neutralizing antibody titer in intensive care unit patients with COVID-19. Emerg. Microbes Infect. 2020, 9, 1664–1670. [Google Scholar] [CrossRef] [PubMed]
- Boehm, E.; Kronig, I.; Neher, R.A.; Eckerle, I.; Vetter, P.; Kaiser, L. Novel SARS-CoV-2 variants: The pandemics within the pandemic. Clin. Microbiol. Infect. 2021, 27, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Lu, L.; Choi, C.Y.K.; Cai, J.P.; Tsoi, H.W.; Chu, A.W.H.; Ip, J.D.; Chan, W.M.; Zhang, R.R.; Zhang, X.; et al. Impact of SARS-CoV-2 variant-associated RBD mutations on the susceptibility to serum antibodies elicited by COVID-19 infection or vaccination. Clin. Infect. Dis. 2021, 26, ciab656. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Chu, A.W.H.; Zhang, R.R.; Chan, W.M.; Ip, J.D.; Tsoi, H.W.; Chen, L.L.; Cai, J.P.; Lung, D.C.; Tam, A.R.; et al. The impact of spike n501y mutation on neutralizing activity and rbd binding of SARS-CoV-2 convalescent serum. EBioMedicine 2021, 71, 103544. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Chan, J.F.W.; Yuen, T.T.T.; Shuai, H.; Yuan, S.; Wang, Y.; Hu, B.; Yip, C.C.Y.; Tsang, J.O.L.; Huang, X.; et al. Comparative tropism, replication kinetics, and cell damage profiling of SARS-CoV-2 and SARS-CoV with implications for clinical manifestations, transmissibility, and laboratory studies of COVID-19, An observational study. Lancet Microbe 2020, 1, e14–e23. [Google Scholar] [CrossRef]
- GeurtsvanKessel, C.H.; Okba, N.M.; Igloi, Z.; Bogers, S.; Embregts, C.W.; Laksono, B.M.; Leijten, L.; Rokx, C.; Rijnders, B.; Rahamat-Langendoen, J.; et al. An evaluation of COVID-19 serological assays informs future diagnostics and exposure assessment. Nat. Commun. 2020, 11, 3436. [Google Scholar] [CrossRef] [PubMed]
- Gobeil, S.; Janowska, K.; McDowell, S.; Mansouri, K.; Parks, R.; Stalls, V.; Kopp, M.F.; Manne, K.; Saunders, K.O.N.; Edwards, R.J.; et al. Effect of natural mutations of SARS-CoV-2 on spike structure, conformation, and antigenicity. Science 2021, 6, eabi6226. [Google Scholar] [CrossRef]
- Weisberg, S.P.; Connors, T.J.; Zhu, Y.; Baldwin, M.R.; Lin, W.H.; Wontakal, S.; Szabo, P.A.; Wells, S.B.; Dogra, P.; Gray, J.; et al. Distinct antibody responses to SARS-CoV-2 in children and adults across the COVID-19 clinical spectrum. Nat. Immunol. 2021, 22, 25–31. [Google Scholar] [CrossRef]


{kind=link}
| MN Titer | Anti-RBD IgG (AU/mL) | Anti-N IgG (S/CO) | ||||
|---|---|---|---|---|---|---|
| <50 (n = 1) | 50–999 (n = 35) | ≥1000 (n = 22) | <1.4 (n = 5) | 1.4–4.9 (n = 26) | ≥5.0 (n = 27) | |
| Against wild type lineage A (HKU-001a) | ||||||
| 10–20 | 0 (0) | 15 (42.9) | 1 (4.5) | 3 (60) | 7 (26.9) | 6 (22.2) |
| ≥40 | 1 (100) | 20 (57.1) | 21 (95.5) | 2 (40) | 19 (73.1) | 21 (77.8) |
| Against beta variant (B.1.351) | ||||||
| <10 | 1 (100) | 30 (85.7) | 20 (90.9) | 5 (100) | 23 (88.5) | 23 (85.2) |
| 10–20 | 0 (0) | 5 (14.3) | 2 (9.1) | 0 (0) | 3 (11.5) | 4 (14.8) |
| ≥40 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poon, R.W.-S.; Lu, L.; Fong, C.H.-Y.; Ip, T.-C.; Chen, L.-L.; Zhang, R.R.-Q.; Yip, C.C.-Y.; Cheng, V.C.-C.; Chan, K.-H.; Yuen, K.-Y.; et al. Correlation between Commercial Anti-RBD IgG Titer and Neutralization Titer against SARS-CoV-2 Beta Variant. Diagnostics 2021, 11, 2216. https://doi.org/10.3390/diagnostics11122216
Poon RW-S, Lu L, Fong CH-Y, Ip T-C, Chen L-L, Zhang RR-Q, Yip CC-Y, Cheng VC-C, Chan K-H, Yuen K-Y, et al. Correlation between Commercial Anti-RBD IgG Titer and Neutralization Titer against SARS-CoV-2 Beta Variant. Diagnostics. 2021; 11(12):2216. https://doi.org/10.3390/diagnostics11122216
Chicago/Turabian StylePoon, Rosana Wing-Shan, Lu Lu, Carol Ho-Yan Fong, Tak-Chuen Ip, Lin-Lei Chen, Ricky Rui-Qi Zhang, Cyril Chik-Yan Yip, Vincent Chi-Chung Cheng, Kwok-Hung Chan, Kwok-Yung Yuen, and et al. 2021. "Correlation between Commercial Anti-RBD IgG Titer and Neutralization Titer against SARS-CoV-2 Beta Variant" Diagnostics 11, no. 12: 2216. https://doi.org/10.3390/diagnostics11122216