The Oncogenic Roles of PTTG1 and PTTG2 Genes and Pseudogene PTTG3P in Head and Neck Squamous Cell Carcinomas

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. TCGA Data
2.2. Data Analysis
2.3. Gene Analysis
2.4. Functional Enrichment Analysis and Prediction of Gene Function
2.5. Statistical Analysis
2.6. Availability of Data and Materials
3. Results
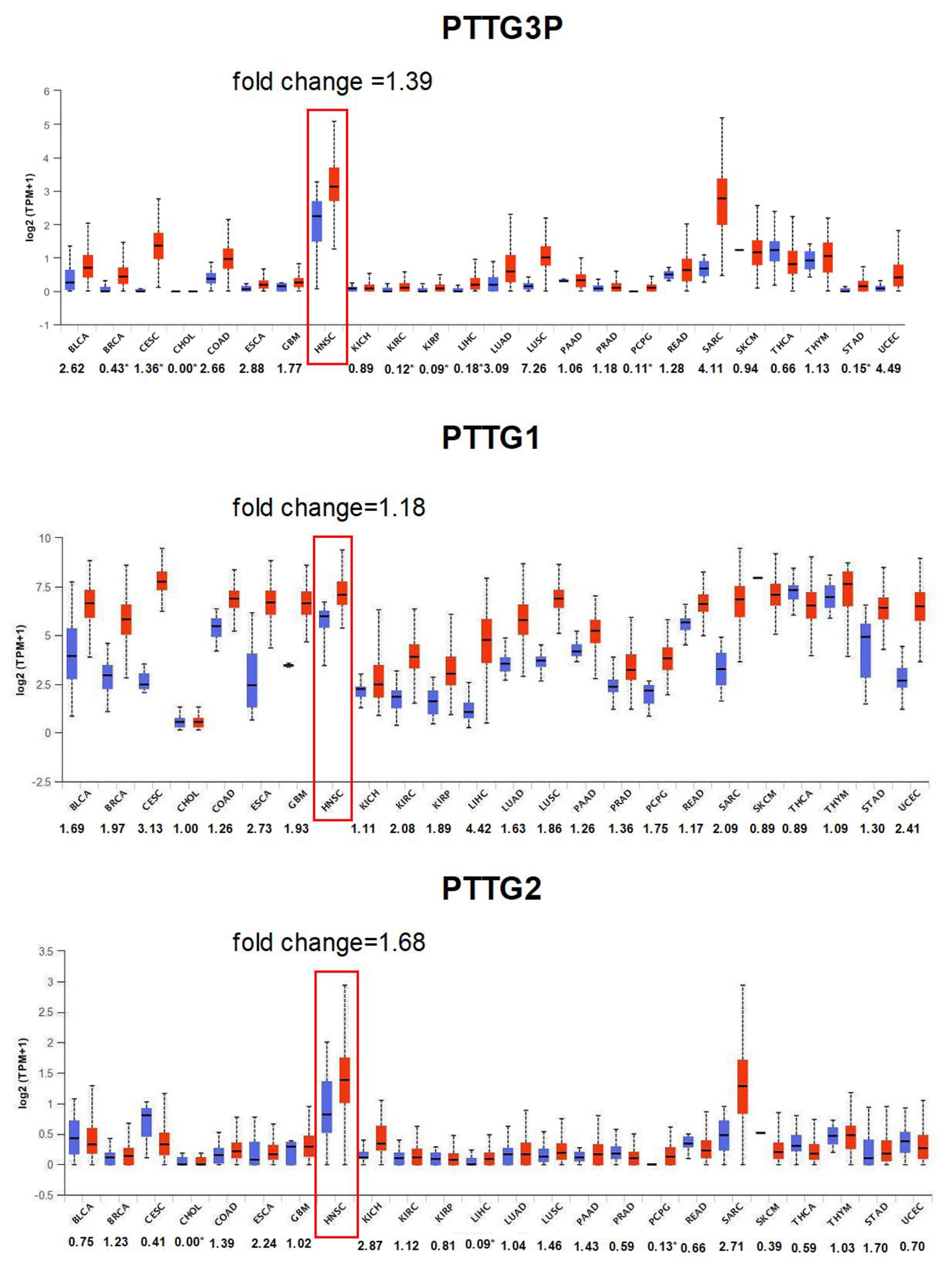
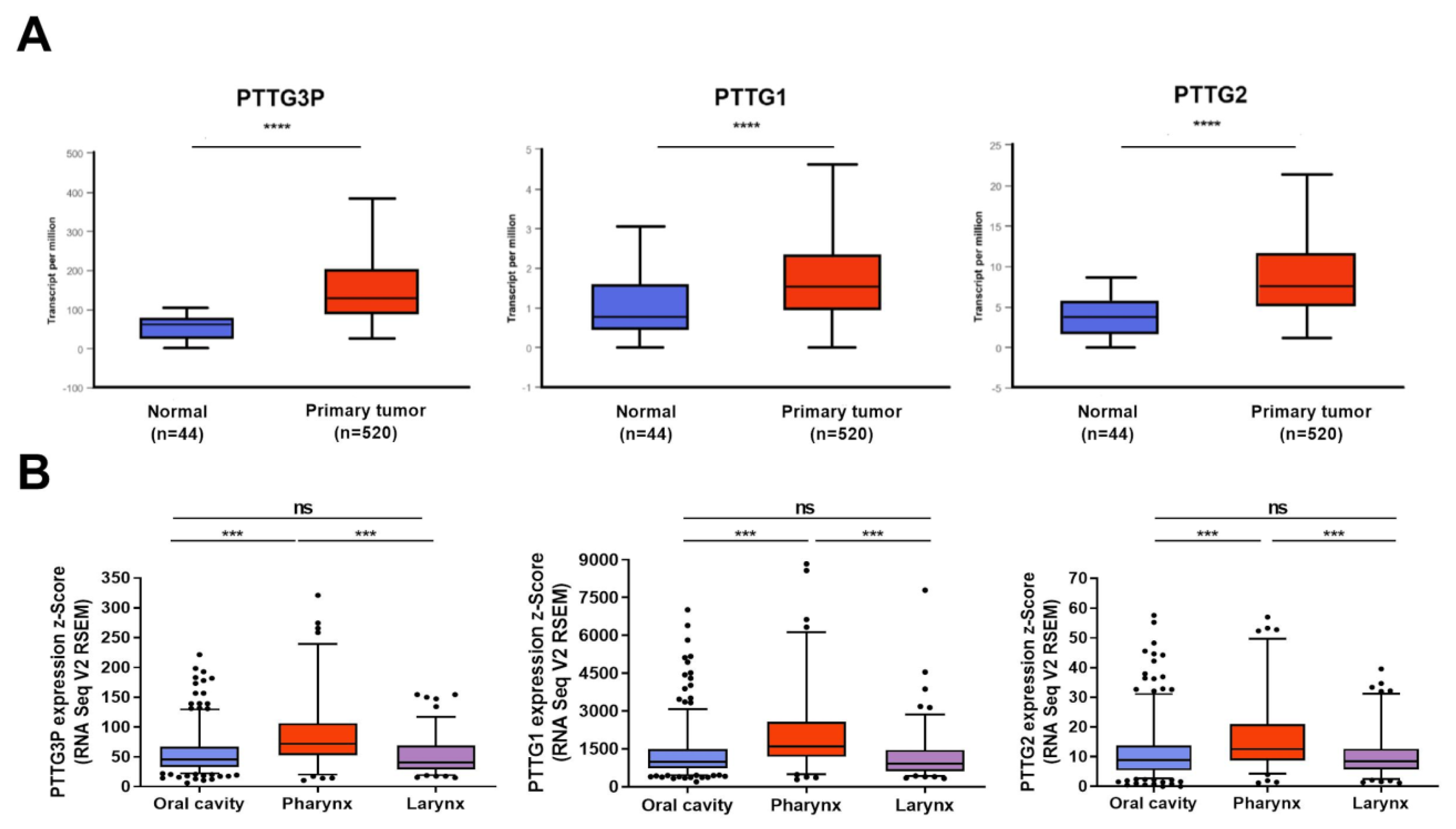
3.1. PTTG3P, PTTG1 and PTTG2 Are Upregulated in Most Cancers, Including HNSCC
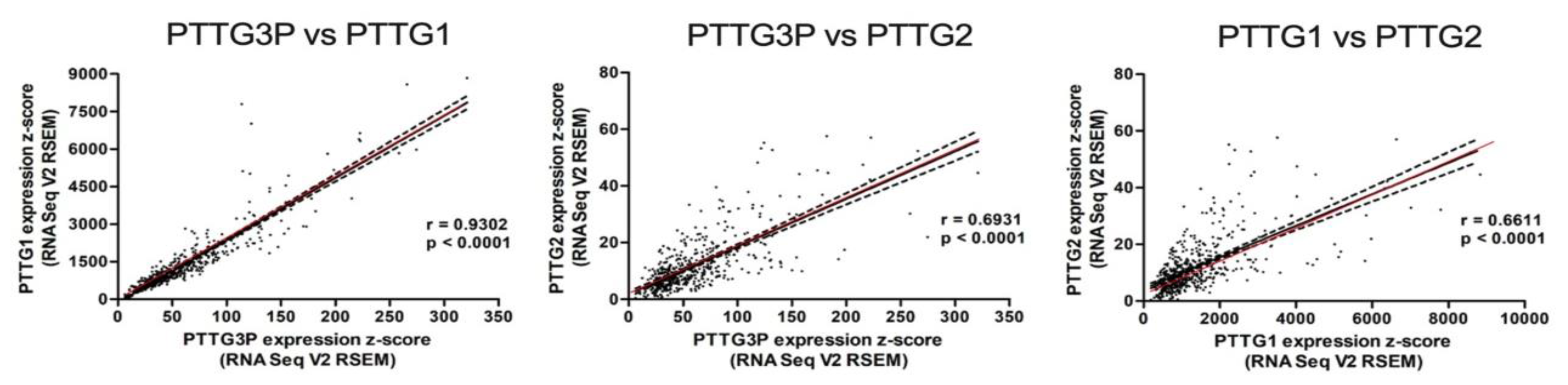
3.2. PTTG3P Positively Correlates with PTTG1 and PTTG2 in HNSCC Patients
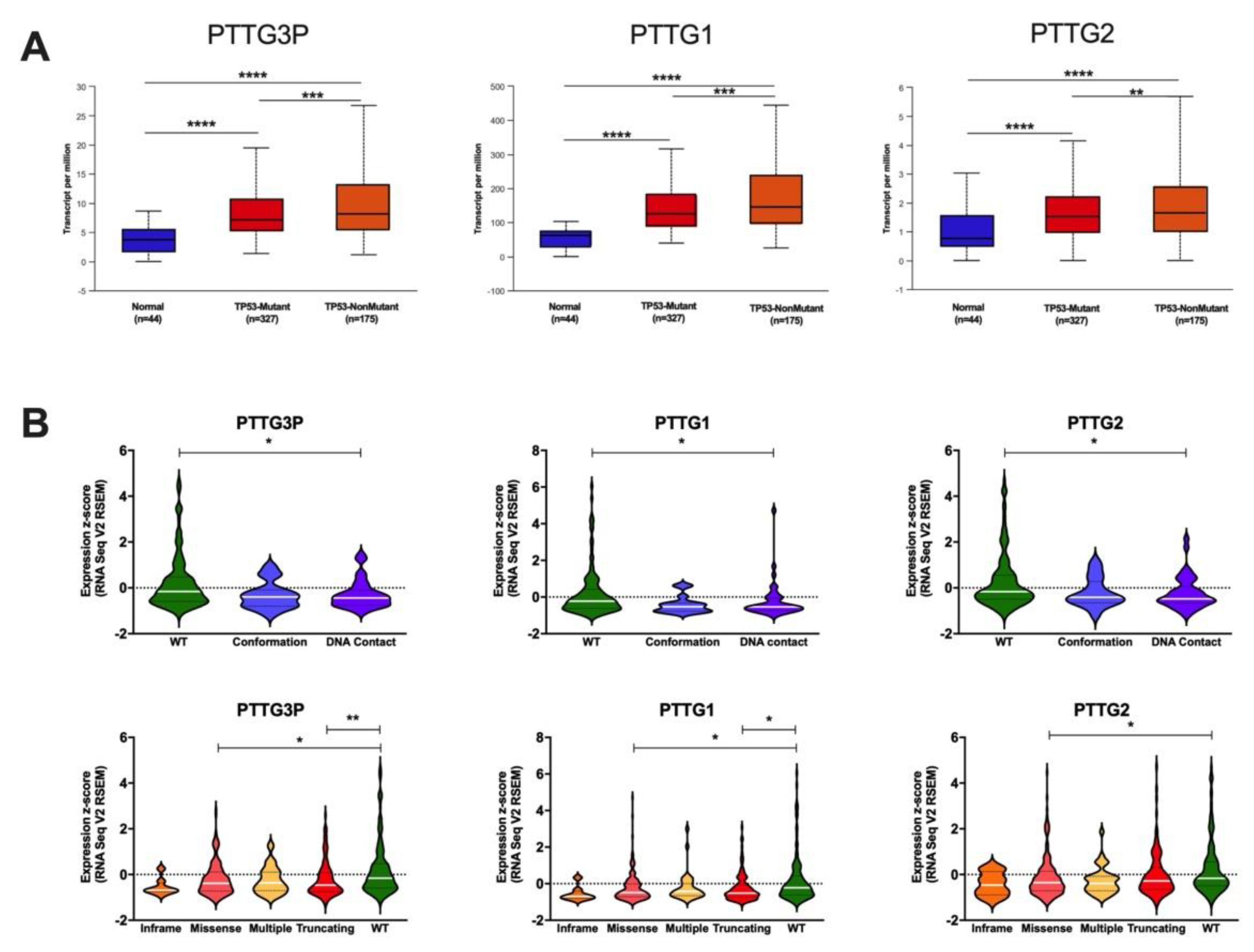
3.3. Expression Levels of PTTG3P, PTTG1 and PTTG2 Depend on TP53 Status and Correlate with Expression of the Genes from the p53 Pathway
3.4. PTTG3P, PTTG1 and PTTG2 Levels Differ Depending on Clinicopathological Parameters
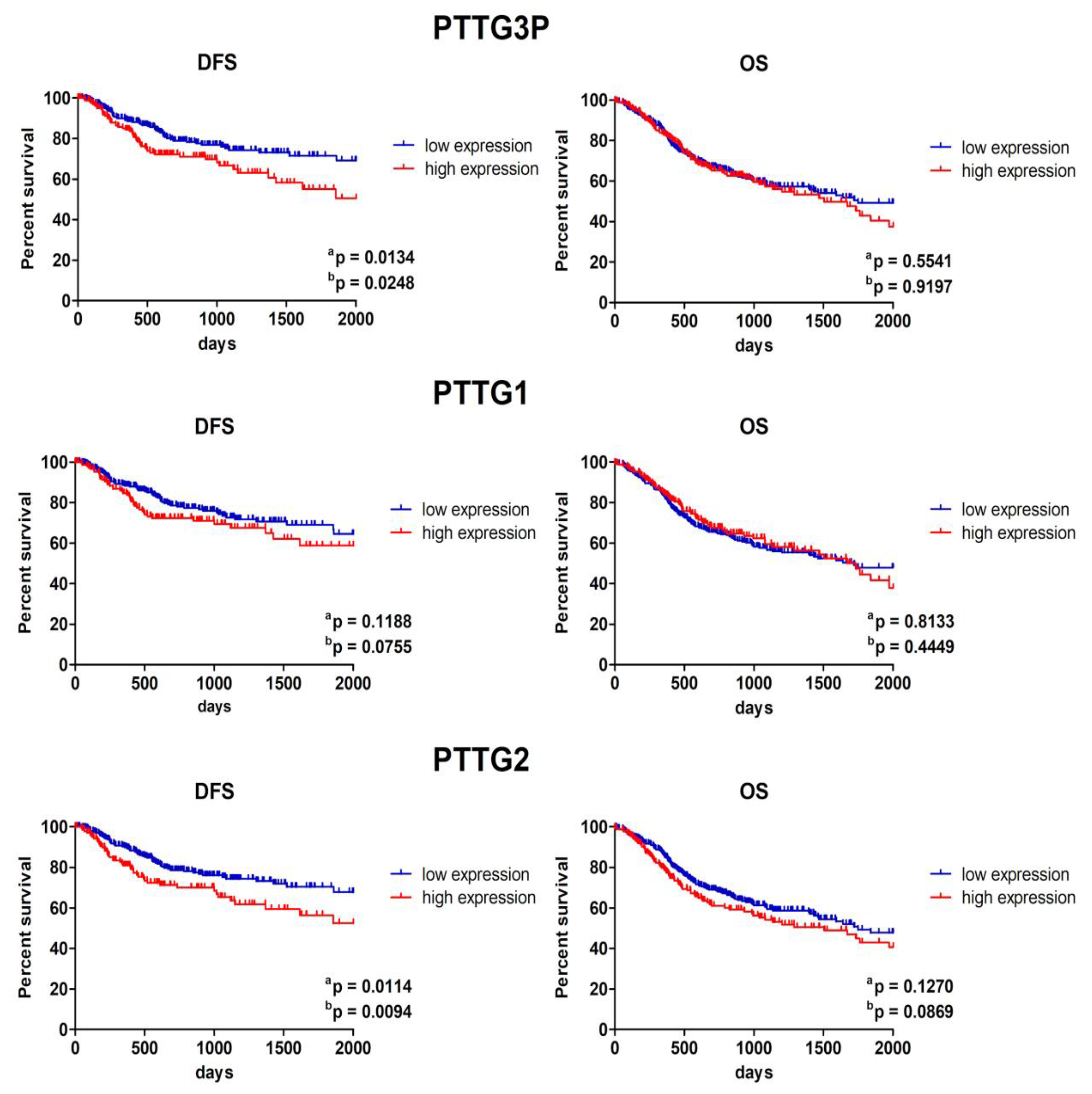
3.5. Patients with Low PTTG3P and PTTG2 Expression Display Longer Disease-Free Survival
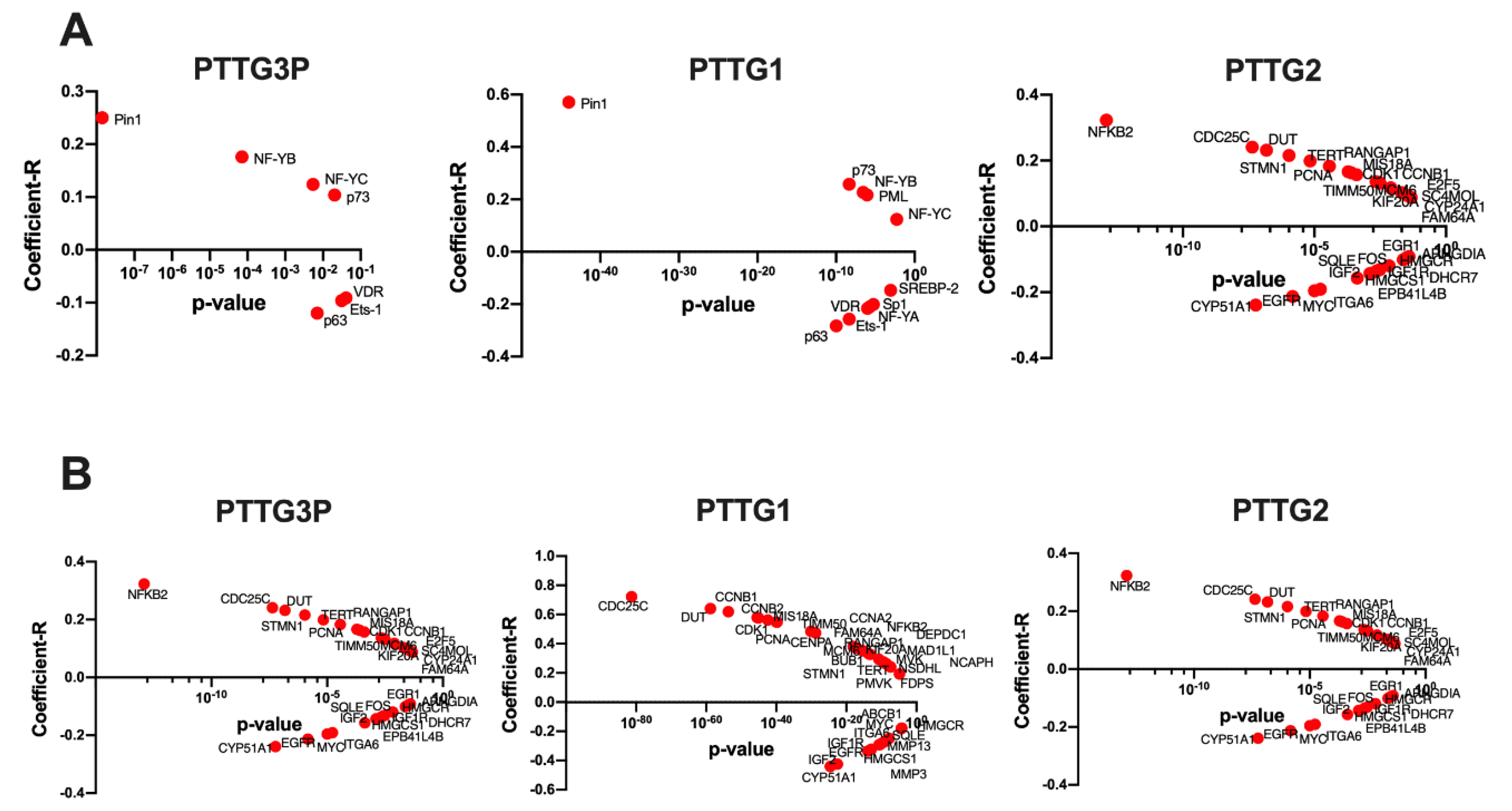
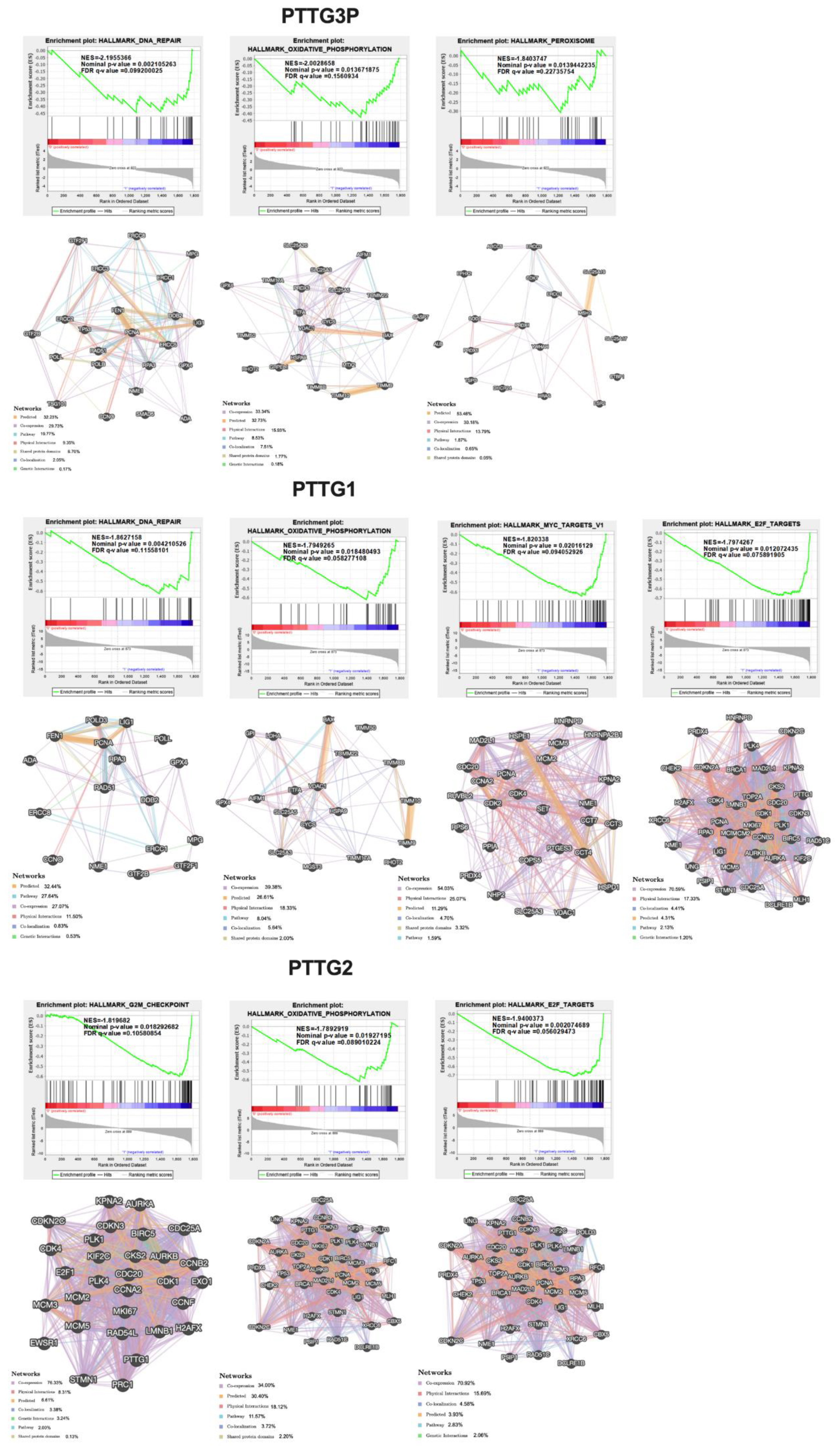
3.6. Expression Levels of PTTG3P, PTTG1 and PTTG2 Correlate with Genes Involved in Important Cellular Processes
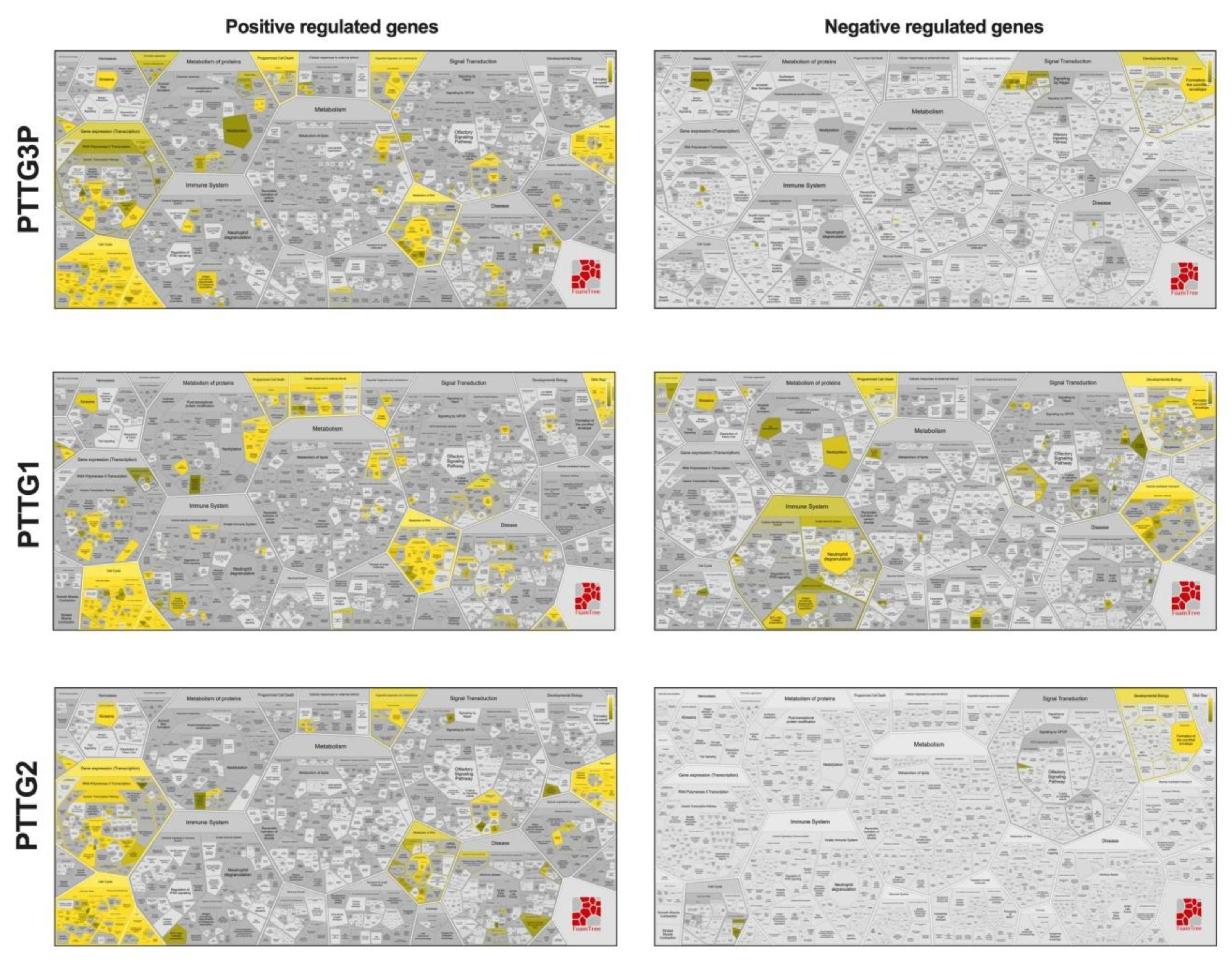
3.7. Patients with High and Low Expressions of PTTG3P, PTTG1 or PTTG2 Have Different Patterns of Genes
4. Discussion
5. Conclusions
- The expression of PTTG3P, PTTG1 and PTTG2 was upregulated in cancer tissue compared to normal in most cancer types including HNSCC and depends on TP53 status.
- Statistical significance was obtained between tumors from the oral cavity and pharynx and between tumors from the pharynx and larynx in all three genes.
- PTTG3P positively correlated with PTTG1 and PTTG2 in HNSCC tissue.
- Patients with low expressions of PTTG3P and PTTG2 displayed significantly longer disease-free survival.
- Analysis of gene correlations and GSEA indicated that patients with high expressions of PTTG3P, PTTG1 and PTTG2 have different expression patterns of genes associated with carcinogenesis pathways in comparison to patients with low expressions of these three transcripts.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BLCA | Bladder urothelial carcinoma |
| BRCA | Breast invasive carcinoma |
| CESC | Cervical squamous cell carcinoma |
| CHOL | Cholangiocarcinoma |
| COAD | Colon adenocarcinoma |
| CT | Chemotherapy |
| DFS | Disease-free survival |
| ESCA | Esophageal carcinoma |
| GBM | Glioblastoma multiforme |
| GSEA | Gene Set Enrichment Analysis |
| HNSCC | Head and Neck squamous cell carcinoma |
| HPV | Human papillomavirus |
| KICH | Kidney Chromophobe |
| KIRC | Kidney renal clear cell carcinoma |
| KIRP | Kidney renal papillary cell carcinoma |
| LIHC | Liver hepatocellular carcinoma |
| LUAD | Lung adenocarcinoma |
| LUSC | Lung squamous cell carcinoma |
| OS | Overall survival |
| OXPHOS | Oxidative phosphorylation |
| PAAD | Pancreatic adenocarcinoma |
| PRAD | Prostate adenocarcinoma |
| RT | Radiotherapy |
| SARC | Sarcoma |
| SKCM | Skin Cutaneous Melanoma |
| TCGA | The Cancer Genome Atlas |
| THCA | Thyroid carcinoma |
| THYM | Thymoma |
| STAD | Stomach adenocarcinoma |
| UCEC | Uterine Corpus Endometrial Carcinoma |
References
- Jou, A.; Hess, J. Epidemiology and Molecular Biology of Head and Neck Cancer. Oncol. Res. Treat. 2017, 40, 328–332. [Google Scholar] [CrossRef]
- Dok, R.; Nuyts, S. HPV Positive Head and Neck Cancers: Molecular Pathogenesis and Evolving Treatment Strategies. Cancers 2016, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, N.; Fedewa, S.; Chen, A.Y. Epidemiology and Demographics of the Head and Neck Cancer Population. Oral Maxillofac. Surg. Clin. N. Am. 2018, 30, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 540. [Google Scholar] [CrossRef] [Green Version]
- Irani, S. miRNAs Signature in Head and Neck Squamous Cell Carcinoma Metastasis: A Literature Review. J. Dent. 2016, 17, 71–83. [Google Scholar]
- An, Y.; Furber, K.L.; Ji, S. Pseudogenes regulate parental gene expression via ceRNA network. J. Cell. Mol. Med. 2017, 21, 185–192. [Google Scholar] [CrossRef]
- Hu, X.; Yang, L.; Mo, Y.-Y. Role of Pseudogenes in Tumorigenesis. Cancers 2018, 10, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalenko, T.F.; Patrushev, L.I. Pseudogenes as Functionally Significant Elements of the Genome. Biochemistry 2018, 83, 1332–1349. [Google Scholar] [CrossRef]
- Gao, L.; Ren, W.; Zhang, L.; Li, S.; Kong, X.; Zhang, H.; Dong, J.; Cai, G.; Jin, C.; Zheng, D.; et al. PTENp1, a natural sponge of miR-21, mediates PTEN expression to inhibit the proliferation of oral squamous cell carcinoma. Mol. Carcinog. 2016, 56, 1322–1334. [Google Scholar] [CrossRef]
- Liu, J.; Xing, Y.; Xu, L.; Chen, W.; Cao, W.; Zhang, C. Decreased expression of pseudogene PTENP1 promotes malignant behaviours and is associated with the poor survival of patients with HNSCC. Sci. Rep. 2017, 7, 41179. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yin, C.; Zhang, B.; Sun, Y.; Shi, L.; Liu, N.; Liang, S.; Lu, S.; Liu, Y.; Zhang, J.; et al. PTTG1 promotes migration and invasion of human non-small cell lung cancer cells and is modulated by miR-186. Carcinogenesis 2013, 34, 2145–2155. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Puri, R.; Lefkowitz, E.; Kakar, S.S. Identification of the human pituitary tumor transforming gene (hPTTG) family: Molecular structure, expression, and chromosomal localization. Gene 2000, 248, 41–50. [Google Scholar] [CrossRef]
- Guo, Y.; Shao, Y.; Chen, J.; Xu, S.; Zhang, X.; Liu, H. Expression of pituitary tumor-transforming 2 in human glioblastoma cell lines and its role in glioblastomatumorigenesis. Exp. Ther. Med. 2016, 11, 1847–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Mateos, M.A.; Espina, Á.G.; Torres, B.; del Estal, M.M.G.; Romero-Franco, A.; Ríos, R.M.; Pintor-Toro, J.A. PTTG1/securin modulates microtubule nucleation and cell migration. Mol. Biol. Cell 2011, 22, 4302–4311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.-B.; Li, F.; Li, Y.-Q.; Yang, F. Pituitary tumor transforming gene PTTG2 induces psoriasis by regulating vimentin and E-cadherin expression. Int. J. Clin. Exp. Pathol. 2015, 8, 10887–10893. [Google Scholar]
- Zhang, Z.; Shi, Z. The pseudogene PTTG3P promotes cell migration and invasion in esophageal squamous cell carcinoma. Open Med. 2019, 14, 516–522. [Google Scholar] [CrossRef]
- Guo, X.-C.; Li, L.; Gao, Z.-H.; Zhou, H.-W.; Li, J.; Wang, Q.-Q. The long non-coding RNA PTTG3P promotes growth and metastasis of cervical cancer through PTTG1. Aging 2019, 11, 1333–1341. [Google Scholar] [CrossRef]
- Huang, J.-L.; Cao, S.-W.; Ou, Q.-S.; Yang, B.; Zheng, S.-H.; Tang, J.; Chen, J.; Hu, Y.-W.; Zheng, L.; Wang, Q. The long non-coding RNA PTTG3P promotes cell growth and metastasis via up-regulating PTTG1 and activating PI3K/AKT signaling in hepatocellular carcinoma. Mol. Cancer 2018, 17, 93. [Google Scholar] [CrossRef]
- Bernal, J.A.; Hernández, A. p53 stabilization can be uncoupled from its role in transcriptional activation by loss of PTTG1/securin. J. Biochem. 2007, 141, 737–745. [Google Scholar] [CrossRef]
- Méndez-Vidal, C.; Estal, M.D.M.G.-D.; Moreno-Mateos, M.A.; Espina-Zambrano, Á.G.; Torres, B.; Pintor-Toro, J.A. PTTG2 silencing results in induction of epithelial-to-mesenchymal transition and apoptosis. Cell Death Dis. 2013, 4, e530. [Google Scholar] [CrossRef] [Green Version]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in Context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Liu, Z.; Myers, J.N. TP53 Mutations in Head and Neck Squamous Cell Carcinoma and Their Impact on Disease Progression and Treatment Response. J. Cell. Biochem. 2016, 117, 2682–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freed-Pastor, W.A.; Prives, C. Mutant p53: One name, many proteins. Genes Dev. 2012, 26, 1268–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caponio, V.C.A.; Troiano, G.; Adipietro, I.; Zhurakivska, K.; Arena, C.; Mangieri, D.; Mascitti, M.; Cirillo, N.; Muzio, L.L. Computational analysis of TP53 mutational landscape unveils key prognostic signatures and distinct pathobiological pathways in head and neck squamous cell cancer. Br. J. Cancer 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, l1. [Google Scholar]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Li, J.-H.; Liu, S.; Zhou, H.; Qu, L.-H.; Yang, J.-H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [Green Version]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Klimczak, M.; Biecek, P.; Zylicz, A.; Zylicz, M. Heat shock proteins create a signature to predict the clinical outcome in breast cancer. Sci. Rep. 2019, 9, 7507. [Google Scholar] [CrossRef] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38 (Suppl. 2), W214–W220. [Google Scholar] [CrossRef]
- Lin, X.; Yang, Y.; Guo, Y.; Liu, H.; Jiang, J.; Zheng, F.; Wu, B. PTTG1 is involved in TNF-α-related hepatocellular carcinoma via the induction of c-myc. Cancer Med. 2019, 8, 5702–5715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Q.; Jin, B. The clinical value and biological function of PTTG1 in colorectal cancer. Biomed. Pharmacother. 2017, 89, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Lou, W.; Ding, B.; Fan, W. High Expression of Pseudogene PTTG3P Indicates a Poor Prognosis in Human Breast Cancer. Mol. Ther. Oncol. 2019, 14, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, M.L.; Modasia, B.; Fletcher, A.; Thompson, R.J.; Brookes, K.; Rae, P.C.; Nieto, H.R.; Poole, V.L.; Roberts, S.; Campbell, M.J.; et al. PTTG and PBF functionally interact with p53 and predict overall survival in head and neck cancer. Cancer Res. 2018, 78. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanachetty, S.; Balaiya, V.; High, W.A.; Koster, M.I. Loss of TP63 Promotes the Metastasis of Head and Neck Squamous Cell Carcinoma by Activating MAPK and STAT3 Signaling. Mol. Cancer Res. 2019, 17, 1279–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roychowdhury, A.; Samadder, S.; Das, P.; Mazumder, D.I.; Chatterjee, A.; Addya, S.; Mondal, R.; Roy, A.; Roychoudhury, S.; Panda, C.K. Deregulation of H19 is associated with cervical carcinoma. Genomics 2020, 112, 961–970. [Google Scholar] [CrossRef]
- Solbach, C.; Roller, M.; Eckerdt, F.; Peters, J.M.; Knecht, R. Pituitary tumor-transforming gene expression is a prognostic marker for tumor recurrence in squamous cell carcinoma of the head and neck. BMC Cancer 2006, 6, 242. [Google Scholar] [CrossRef] [Green Version]
- Shibata, Y.; Haruki, N.; Kuwabara, Y.; Nishiwaki, T.; Kato, J.; Shinoda, N.; Sato, A.; Kimura, M.; Koyama, H.; Toyama, T.; et al. Expression of PTTG (pituitary tumor transforming gene) in esophageal cancer. Jpn. J. Clin. Oncol. 2002, 32, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Guo, J.; Zhou, J.J.; Lu, J.-J.; Chen, Q.; Zhang, C.; Qing, C.; Koeffler, H.P.; Tong, Y. FoxM1 transactivates PTTG1 and promotes colorectal cancer cell migration and invasion. BMC Med. Genom. 2015, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Weng, W.; Ni, S.; Wang, Y.; Xu, M.; Zhang, Q.; Yang, Y.; Wu, Y.; Xu, Q.; Qi, P.; Tan, C.; et al. PTTG3P promotes gastric tumour cell proliferation and invasion and is an indicator of poor prognosis. J. Cell. Mol. Med. 2017, 21, 3360–3371. [Google Scholar] [CrossRef]
- Yang, S.; Wang, X.; Liu, J.; Ding, B.; Shi, K.; Chen, J.; Lou, W. Distinct expression pattern and prognostic values of pituitary tumor transforming gene family genes in non-small cell lung cancer. Oncol. Lett. 2019, 18, 4481–4494. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Cantalupo, P.G.; Pipas, J.M. Identification of Head and Neck Cancer Subtypes Based on Human Papillomavirus Presence and E2F-Regulated Gene Expression. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaubatz, S.; Lindeman, G.J.; Ishida, S.; Jakoi, L.; Nevins, J.R.; Livingston, D.M.; Rempel, R.E. E2F4 and E2F5 Play an Essential Role in Pocket Protein–Mediated G1 Control. Mol. Cell 2000, 6, 729–735. [Google Scholar] [CrossRef]
- Burke, P. Mitochondria, Bioenergetics and Apoptosis in Cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Zacksenhaus, E.; Shrestha, M.; Liu, J.C.; Vorobieva, I.; Chung, P.E.; Ju, Y.; Nir, U.; Jiang, Z. Mitochondrial OXPHOS Induced by RB1 Deficiency in Breast Cancer: Implications for Anabolic Metabolism, Stemness, and Metastasis. Trends Cancer 2017, 3, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, S.; Klijanienko, J.; Giaginis, C.; Rodriguez, J.; Jouffroy, T.; Girod, A.; Point, D.; Tsourouflis, G.; Satre-Garau, X. Peroxisome proliferator-activated receptor-γ in mobile tongue squamous cell carcinoma: Associations with clinicopathological parameters and patients survival. J. Cancer Res. Clin. Oncol. 2011, 137, 251–259. [Google Scholar] [CrossRef]
- Miller, D.M.; Thomas, S.D.; Islam, A.; Muench, D.; Sedoris, K. c-Myc and Cancer Metabolism. Clin. Cancer Res. 2012, 18, 5546–5553. [Google Scholar] [CrossRef] [Green Version]
- Stark, G.R.; Taylor, W.R. Analyzing the G2/M checkpoint. Methods Mol. Biol. 2004, 280, 51–82. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PTTG3P | PTTG1 | PTTG2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Group | Mean ± SEM | p-Value | n | Mean ± SEM | p-Value | n | Mean ± SEM | p-Value | n |
| Grade | G1 + G2 | 55.57 ± 2.016 | <0.0001 | 368 | 1284 ± 58.95 | <0.0001 | 368 | 11.17 ± 0.4642 | 0.005 | 368 |
| G3 + G4 | 73.03 ± 4.117 | 132 | 1623 ± 101.1 | 132 | 14.09 ± 0.9859 | 132 | ||||
| HPV p16 status | Negative | 46.65 ± 3.023 | <0.0001 | 80 | 958.6 ± 62.19 | <0.0001 | 73 | 8.528 ± 0.6258 | <0.0001 | 73 |
| Positive | 102.6 ± 10.77 | 42 | 2601 ± 314.9 | 39 | 18.47 ± 1.983 | 39 | ||||
| Lymph Node Neck Dissection | Positive | 59.05 ± 2.062 | 0.800 | 422 | 1382 ± 58.68 | 0.033 | 422 | 11.42 ± 0.4430 | 0.003 | 422 |
| Negative | 68.07 ± 3.750 | 97 | 1420 ± 80.86 | 97 | 14.59 ± 1.133 | 97 | ||||
| Angiolymphatic Invasion | Positive | 66.06 ± 4.830 | 0.154 | 125 | 1575 ± 132.8 | 0.098 | 125 | 13.56 ±1.061 | 0.034 | 125 |
| Negative | 55.19 ± 2.428 | 225 | 1273 ± 72.55 | 225 | 10.61 ± 0.5364 | 225 | ||||
| Perineural Invasion | Positive | 53.03 ± 2.570 | 0.557 | 169 | 1230 ± 75.92 | 0.032 | 167 | 10.97 ± 0.6415 | 0.210 | 169 |
| Negative | 64.66 ± 3.580 | 195 | 1536 ± 102.5 | 193 | 12.42 ± 0.7516 | 195 | ||||
| Age | <60 | 62.39 ± 2.842 | 0.863 | 258 | 1454 ± 80.86 | 0.808 | 258 | 12.26 ± 0.6242 | 0.918 | 258 |
| >60 | 58.88 ± 2.276 | 263 | 1316 ± 58.67 | 263 | 11.82 ± 0.5682 | 263 | ||||
| Gender | Female | 56.10 ± 3.600 | 0.077 | 137 | 1251 ± 94.08 | 0.055 | 137 | 10.98 ± 0.7738 | 0.070 | 137 |
| Male | 62.25 ± 2.095 | 385 | 1434 ± 58.47 | 385 | 12.41 ± 0.4985 | 385 | ||||
| Alcohol | Positive | 60.26 ± 2.235 | 0.494 | 348 | 1363 ± 58.02 | 0.606 | 348 | 11.59 ± 0.5001 | 0.151 | 348 |
| Negative | 62.48 ± 3.258 | 163 | 1454 ± 99.58 | 163 | 13.06 ± 0.8033 | 163 | ||||
| Smoking | No/Ex | 58.45 ± 2.628 | 0.911 | 191 | 1349 ± 60.24 | 0.711 | 294 | 11.74 ± 0.5441 | 0.708 | 294 |
| Yes | 61.6 ± 2.487 | 319 | 1406 ± 82.29 | 216 | 12.07 ± 0.6676 | 216 | ||||
| Cancer Stage | I + II | 55.40 ± 2.872 | 0.577 | 101 | 1154 ± 61.18 | 0.686 | 101 | 11.12 ± 0.8306 | 0.823 | 101 |
| III + IV | 59.57 ± 2.371 | 349 | 1393 ± 66.51 | 349 | 11.93 ± 0.5197 | 349 | ||||
| T Stage | T1 + T2 | 63.94 ± 3.409 | 0.037 | 185 | 1434 ± 88.89 | 0.106 | 185 | 12.3 ± 0.7339 | 0.354 | 185 |
| T3 + T4 | 55.54 ± 2.266 | 274 | 1284 ± 65.23 | 274 | 11.48 ± 0.5600 | 274 | ||||
| N Stage | N0 + N1 | 56.15 ± 2.360 | 0.365 | 243 | 1266 ± 63.75 | 0.196 | 243 | 10.65 ± 0.5611 | 0.014 | 243 |
| N2 + N3 | 62.03 ± 3.618 | 179 | 1474 ± 100.9 | 179 | 12.99 ± 0.7680 | 179 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzechowiak, I.; Graś, J.; Szymańska, D.; Biernacka, M.; Guglas, K.; Poter, P.; Mackiewicz, A.; Kolenda, T. The Oncogenic Roles of PTTG1 and PTTG2 Genes and Pseudogene PTTG3P in Head and Neck Squamous Cell Carcinomas. Diagnostics 2020, 10, 606. https://doi.org/10.3390/diagnostics10080606
Grzechowiak I, Graś J, Szymańska D, Biernacka M, Guglas K, Poter P, Mackiewicz A, Kolenda T. The Oncogenic Roles of PTTG1 and PTTG2 Genes and Pseudogene PTTG3P in Head and Neck Squamous Cell Carcinomas. Diagnostics. 2020; 10(8):606. https://doi.org/10.3390/diagnostics10080606
Chicago/Turabian StyleGrzechowiak, Inga, Justyna Graś, Dominika Szymańska, Martyna Biernacka, Kacper Guglas, Paulina Poter, Andrzej Mackiewicz, and Tomasz Kolenda. 2020. "The Oncogenic Roles of PTTG1 and PTTG2 Genes and Pseudogene PTTG3P in Head and Neck Squamous Cell Carcinomas" Diagnostics 10, no. 8: 606. https://doi.org/10.3390/diagnostics10080606