COLD-PCR Method for Early Detection of Antiviral Drug-Resistance Mutations in Treatment-Naive Children with Chronic Hepatitis B


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Patients Samples
2.2. Design of Primers for Amplification of Specific Sequences Containing NA Resistance mt
- Forward-HBV: 5′-TCCTGCTCAAGGAACCTCTATG-3′ (nt 532–553)
- Reverse-HBV: 5′-TGTACAATATGTTCCTGTGG-3′ (nt 910–929).
- Forward-HBV-P1: 5′-CACCTGTATTCCCATCCCATC-3′ (nt 595–615)
- Reverse-HBV-P1: 5′-AGGGACTCAAGATGTTGTACA-3′ (nt 768–788).
- Forward-HBV-P2: 5′-CAGTTATATGGATGATGTGGTATTGG-3′ (nt 732–757)
- Reverse-HBV-P2: 5′-TGTACAATATGTTCCTGTGG-3′ (nt 910–929).
2.3. Conventional PCR Combined with Sanger DNA Sequencing
2.4. Cloning and Synthesis of RT-HBV wt and Representative mt Controls
2.5. Optimizing COLD-PCR Conditions and Evaluating Sensitivity of COLD-PCR Assay Combined with Sanger DNA Sequencing
2.6. Other Molecular and Serological Assays
2.7. DNA Sequence and Statistical Analysis
2.8. Real-Time PCR for Confirming NA Drug-Resistance-Associated mt
3. Results
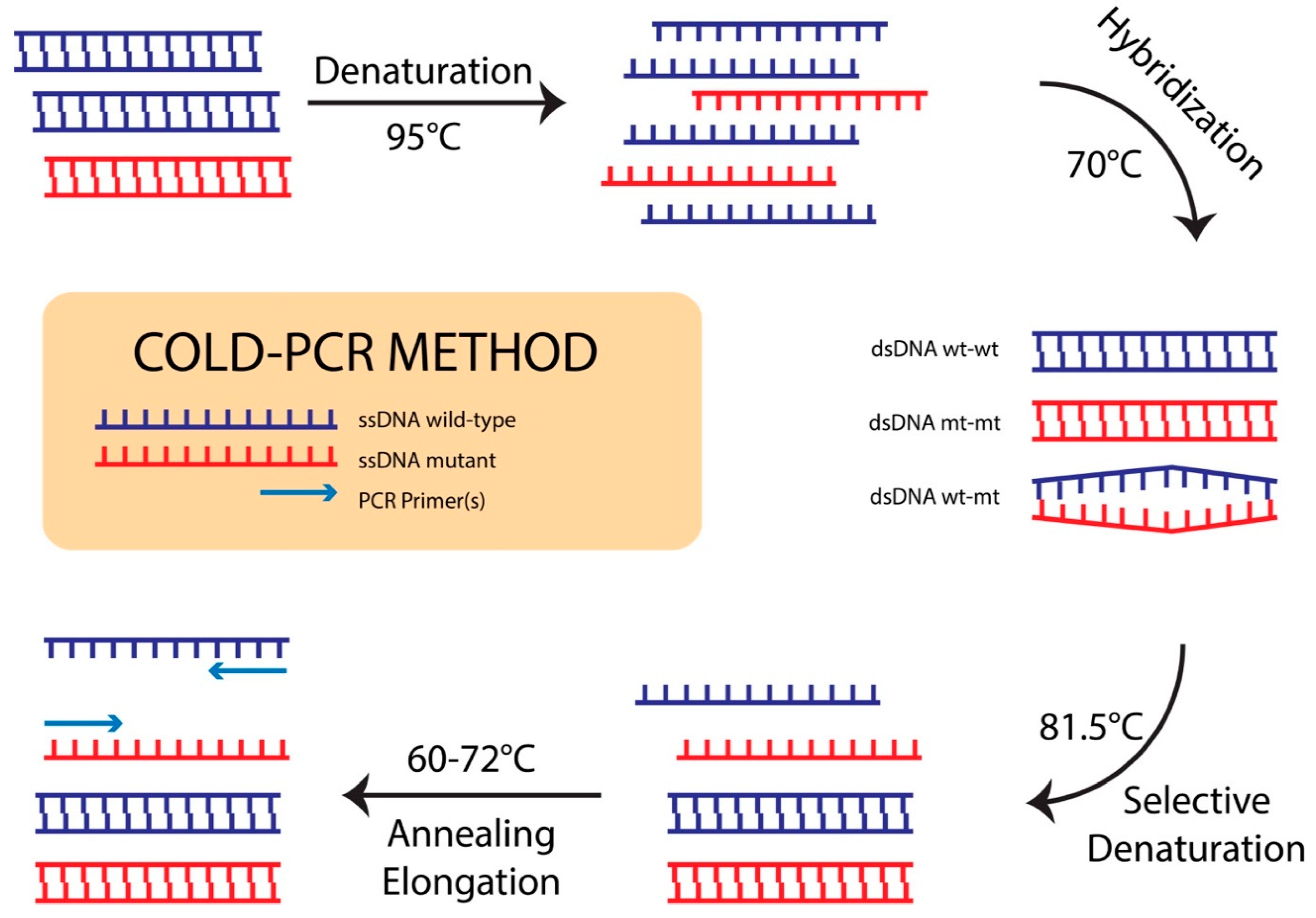
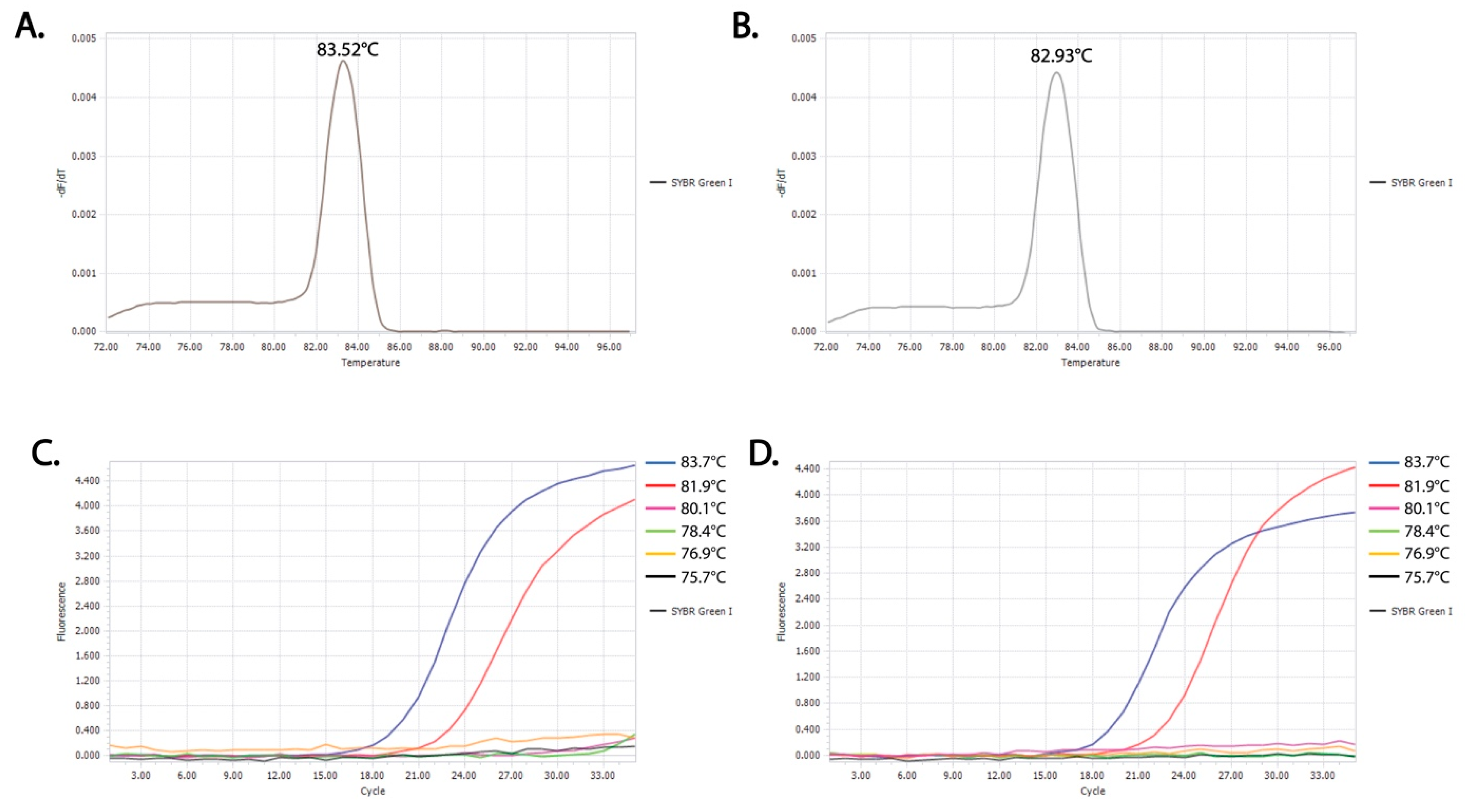
3.1. Identification of Critical Denaturation Temperatures
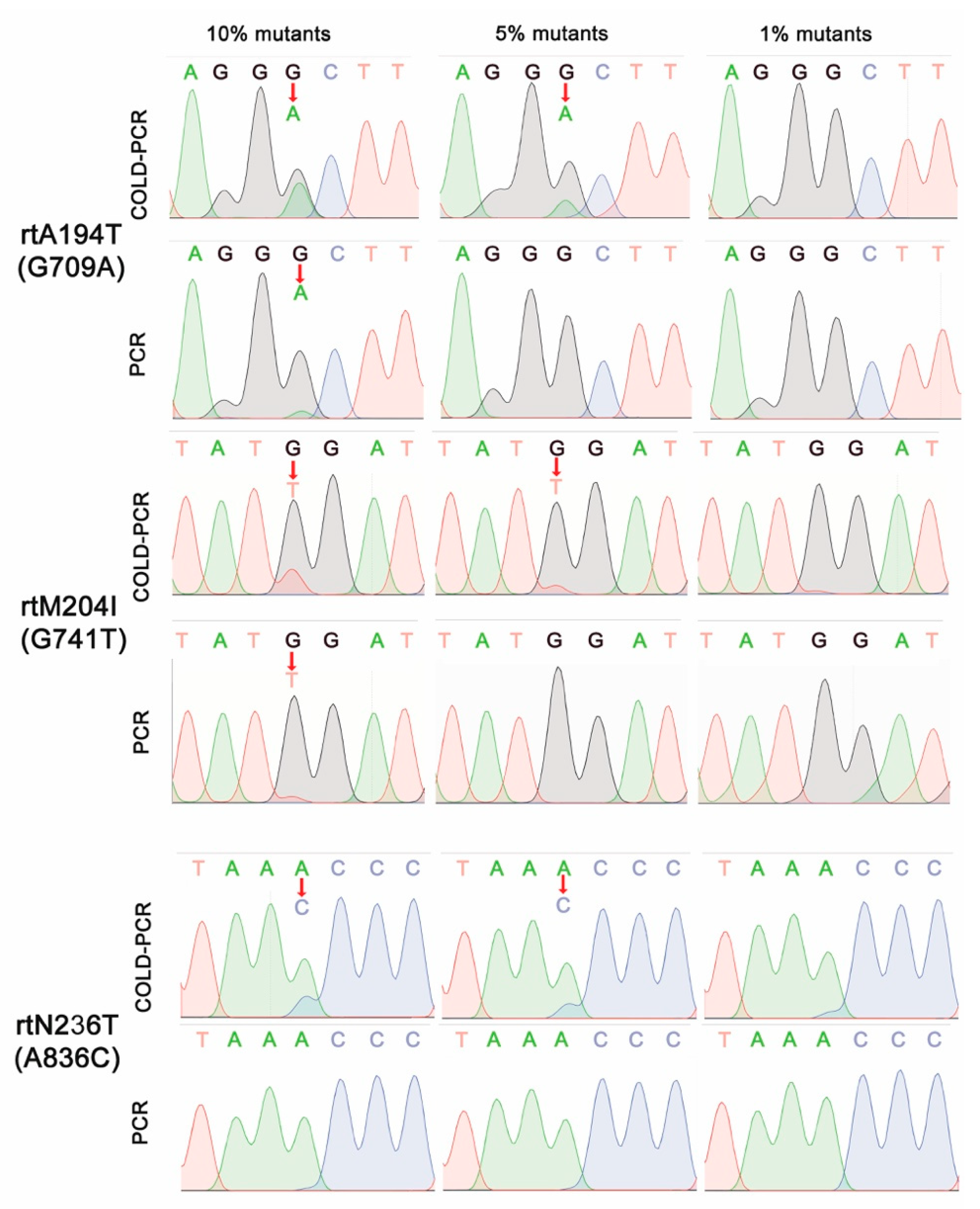
3.2. Sensitive DNA Sequencing Coupled with COLD-PCR Amplification for Detection of NA-Resistance mt
3.3. Characteristics of Subjects
3.4. Rate and Association of NA-Resistance Mutations with Characteristics and Subclinical Index of Treatment-Naive CHB Children
3.5. Profile of Mutations in Treatment-Naive CHB Children
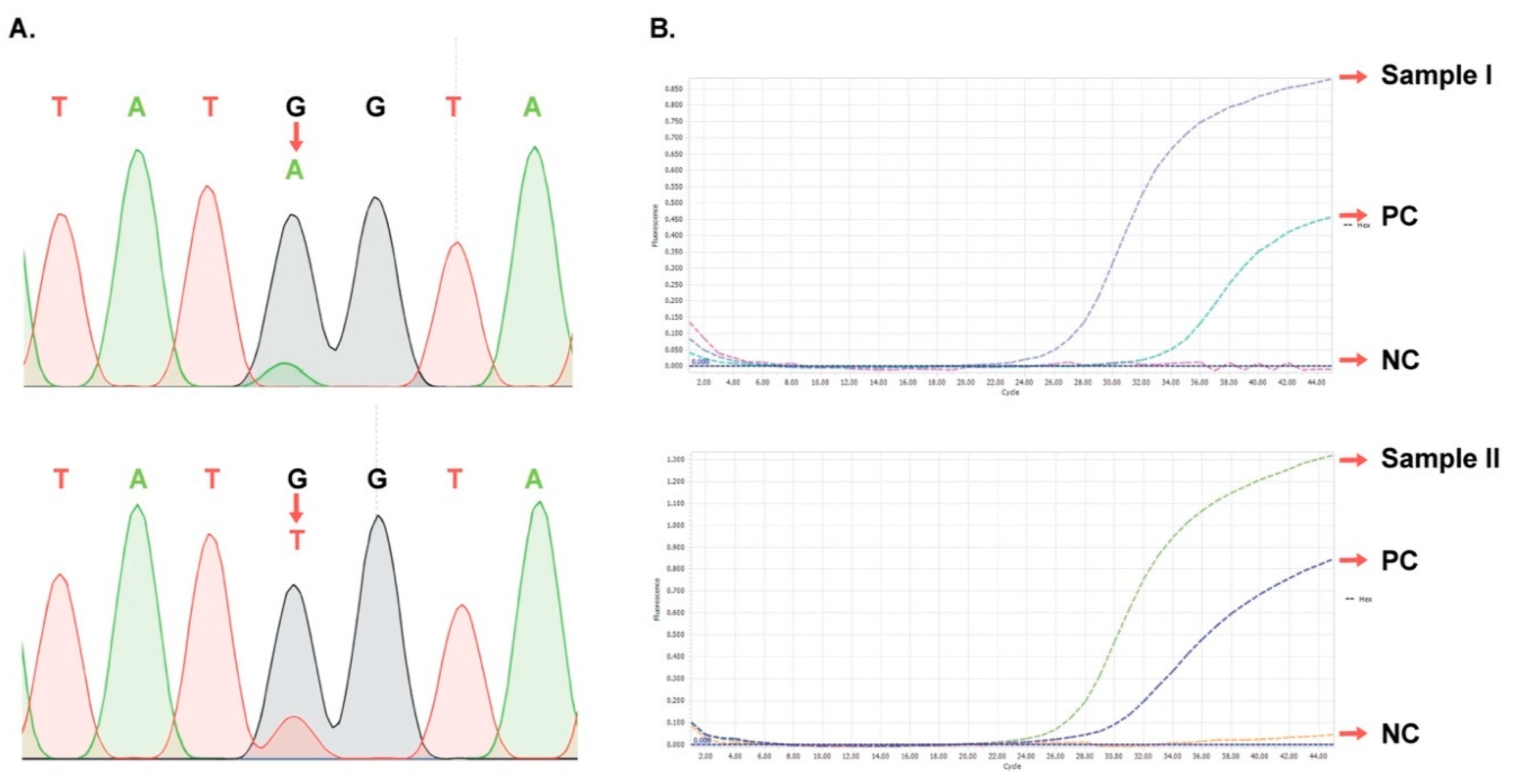
3.6. Verification of Representative Samples Having Minor mt Peaks
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, D.-Z.; Yan, Y.-P.; Choi, B.C.K.; Xu, J.-Q.; Men, K.; Zhang, J.-X.; Liu, Z.-H.; Wang, F.-S. Risk factors and mechanism of transplacental transmission of hepatitis B virus: A case-control study. J. Med. Virol. 2002, 67, 20–26. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Yang, H.; Li, X.; Wen, S.; Guo, Y.; Sun, J.; Hou, J. Quantitative analysis of HBV DNA level and HBeAg titer in hepatitis B surface antigen positive mothers and their babies: HBeAg passage through the placenta and the rate of decay in babies. J. Med. Virol. 2003, 71, 360–366. [Google Scholar] [CrossRef]
- Wiseman, E.; Fraser, M.A.; Holden, S.; Glass, A.; Kidson, B.L.; Heron, L.G.; Maley, M.W.; Ayres, A.; Locarnini, S.A.; Levy, M.T. Perinatal transmission of hepatitis B virus: An Australian experience. Med. J. Aust. 2009, 190, 489–492. [Google Scholar] [CrossRef]
- Dusheiko, G. Interruption of mother-to-infant transmission of hepatitis B: Time to include selective antiviral prophylaxis? Lancet 2012, 379, 2019–2021. [Google Scholar] [CrossRef]
- Lok, A.S.F.; McMahon, B.J. Chronic hepatitis B: Update 2009. Hepatology 2009, 50, 661–662. [Google Scholar] [CrossRef]
- Sokal, E.M.; Paganelli, M.; Wirth, S.; Socha, P.; Vajro, P.; Lacaille, F.; Kelly, D.; Mieli-Vergani, G. Management of chronic hepatitis B in childhood: ESPGHAN clinical practice guidelines. J. Hepatol. 2013, 59, 814–829. [Google Scholar] [CrossRef] [Green Version]
- European Association for the Study of the Liver. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, H.; Inui, A.; Fujisawa, T. Pediatric hepatitis B treatment. J. Thorac. Dis. 2017, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrault, N.A.; Lok, A.S.F.; McMahon, B.J.; Chang, K.; Hwang, J.P.; Jonas, M.M.; Brown, R.S.; Bzowej, N.H.; Wong, J.B. Update on prevention, diagnosis, and treatment of chronic hepatitis B: AASLD 2018 hepatitis B guidance. Hepatology 2018, 67, 1560–1599. [Google Scholar] [CrossRef]
- Ohno, M.; Otsuka, M.; Kishikawa, T.; Yoshikawa, T.; Takata, A.; Koike, K. Novel therapeutic approaches for hepatitis B virus covalently closed circular DNA. World J. Gastroenterol. 2015, 21, 7084–7088. [Google Scholar] [CrossRef] [PubMed]
- Fares, M.A.; Holmes, E.C. A Revised Evolutionary History of Hepatitis B Virus (HBV). J. Mol. Evol. 2002, 54, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Mokaya, J.; McNaughton, A.L.; Hadley, M.J.; Beloukas, A.; Geretti, A.-M.; Goedhals, D.; Matthews, P.C. A systematic review of hepatitis B virus (HBV) drug and vaccine escape mutations in Africa: A call for urgent action. PLOS Negl. Trop. Dis. 2018, 12, e0006629. [Google Scholar] [CrossRef] [Green Version]
- Tacke, F.; Kroy, D.C. Treatment for hepatitis B in patients with drug resistance. Ann. Transl. Med. 2016, 4, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.-S.; Lee, A.R.; Kim, D.H.; Lee, J.-H.; Yoo, J.-J.; Ahn, S.H.; Sim, H.; Park, S.; Kang, H.S.; Won, J.; et al. Identification of a quadruple mutation that confers tenofovir resistance in chronic hepatitis B patients. J. Hepatol. 2019, 70, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Yamani, L.N.; Yano, Y.; Utsumi, T.; Wasityastuti, W.; Rinonce, H.T.; Widasari, D.I.; Juniastuti; Lusida, M.I.; Soetjipto; Hayashi, Y. Profile of Mutations in the Reverse Transcriptase and Overlapping Surface Genes of Hepatitis B Virus (HBV) in Treatment-Naïve Indonesian HBV Carriers. Jpn. J. Infect. Dis. 2017, 70, 647–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-M.; Lee, S.-Y.; Kim, B.-J. Naturally occurring hepatitis B virus reverse transcriptase mutations related to potential antiviral drug resistance and liver disease progression. World J. Gastroenterol. 2018, 24, 1708–1724. [Google Scholar] [CrossRef]
- Xu, J.; Wu, B.; Wang, J.H.; Huang, L.; Wang, D.Y.; Zhao, L.; Zhao, G.; Wang, Y. Pre-existing mutations in reverse transcriptase of hepatitis B virus in treatment-naive chinese patients with chronic hepatitis B. PLoS ONE 2015, 10, 1–12. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Wei, M.; Zhang, C.; Xu, T.; Liu, L.; Xu, Z. Potential resistant mutations within HBV reverse transcriptase sequences in nucleos(t)ide analogues-experienced patients with hepatitis B virus infection. Sci. Rep. 2019, 9, 8078. [Google Scholar] [CrossRef] [Green Version]
- Lok, A.S.; Zoulim, F.; Locarnini, S.; Bartholomeusz, A.; Ghany, M.G.; Pawlotsky, J.-M.; Liaw, Y.-F.; Mizokami, M.; Kuiken, C. Antiviral drug-resistant HBV: Standardization of nomenclature and assays and recommendations for management. Hepatology 2007, 46, 254–265. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Mamon, H.; Kulke, M.H.; Berbeco, R.; Makrigiorgos, G.M. Replacing PCR with COLD-PCR enriches variant DNA sequences and redefines the sensitivity of genetic testing. Nat. Med. 2008, 14, 579–584. [Google Scholar] [CrossRef]
- Liu, C.; Lin, J.; Chen, H.; Shang, H.; Jiang, L.; Chen, J.; Ye, Y.; Yang, B.; Ou, Q. Detection of Hepatitis B Virus Genotypic Resistance Mutations by Coamplification at Lower Denaturation Temperature-PCR Coupled with Sanger Sequencing. J. Clin. Microbiol. 2014, 52, 2933–2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, D.K.-H.; Tsoi, O.; Huang, F.-Y.; Seto, W.-K.; Fung, J.; Lai, C.-L.; Yuen, M.-F. Application of Coamplification at Lower Denaturation Temperature-PCR Sequencing for Early Detection of Antiviral Drug Resistance Mutations of Hepatitis B Virus. J. Clin. Microbiol. 2014, 52, 3209–3215. [Google Scholar] [CrossRef] [Green Version]
- Zoulim, F.; Locarnini, S. Hepatitis B Virus Resistance to Nucleos(t)ide Analogues. Gastroenterology 2009, 137. [Google Scholar] [CrossRef]
- Mahabadi, M.; Norouzi, M.; Alavian, S.M.; Samimirad, K.; Azad, T.M.; Saberfar, E.; Mahmoodi, M.; Ramezani, F.; Karimzadeh, H.; Malekzadeh, R.; et al. Drug-Related Mutational Patterns in Hepatitis B Virus (HBV) Reverse Transcriptase Proteins From Iranian Treatment-Naïve Chronic HBV Patients. Hepat. Mon. 2013, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Wang, F.; Huang, B.; Chen, P.; Zhong, L. Detection and analysis of resistance mutations of hepatitis B virus. Int. J. Clin. Exp. Med. 2015, 8, 9630–9639. [Google Scholar] [PubMed]
- Liu, B.-M.; Li, T.; Xu, J.; Li, X.-G.; Dong, J.-P.; Yan, P.; Yang, J.-X.; Yan, L.; Gao, Z.-Y.; Li, W.-P.; et al. Characterization of potential antiviral resistance mutations in hepatitis B virus reverse transcriptase sequences in treatment-naïve Chinese patients. Antivir. Res. 2010, 85, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Zöllner, B.; Sterneck, M.; Wursthorn, K.; Petersen, J.; Schröter, M.; Laufs, R.; Feucht, H.H. Prevalence, incidence, and clinical relevance of the reverse transcriptase V207I mutation outside the YMDD motif of the hepatitis B virus polymerase during lamivudine therapy. J. Clin. Microbiol. 2005, 43, 2503–2505. [Google Scholar] [CrossRef] [Green Version]
- Bartholomeusz, A.; Locarnini, S. Hepatitis B virus mutations associated with antiviral therapy. J. Med. Virol. 2006, 78, S52–S55. [Google Scholar] [CrossRef]
- Schildgen, O.; Sirma, H.; Funk, A.; Olotu, C.; Wend, U.C.; Hartmann, H.; Helm, M.; Rockstroh, J.K.; Willems, W.R.; Will, H.; et al. Variant of Hepatitis B Virus with Primary Resistance to Adefovir. N. Engl. J. Med. 2006, 354, 1807–1812. [Google Scholar] [CrossRef] [Green Version]
- Yim, H.J.; Hussain, M.; Liu, Y.; Wong, S.N.; Fung, S.K.; Lok, A.S.F. Evolution of multi-drug resistant hepatitis B virus during sequential therapy. Hepatology 2006, 44, 703–712. [Google Scholar] [CrossRef]
- Locarnini, S. Primary resistance, multidrug resistance, and cross-resistance pathways in HBV as a consequence of treatment failure. Hepatol. Int. 2008, 2, 147–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlando, R.; Tosone, G.; Portella, G.; Veropalumbo, E.; D’Onofrio, M.; Piazza, M. Prolonged Persistence of Lamivudine-resistant Mutant and Emergence of New Lamivudine-resistant Mutants Two Years After Lamivudine Withdrawal in HBsAg-positive Chronic Hepatitis Patient: A Case Report. Infection 2008, 36, 472–474. [Google Scholar] [CrossRef] [PubMed]
- Pollicino, T.; Isgrò, G.; Di Stefano, R.; Ferraro, D.; Maimone, S.; Brancatelli, S.; Squadrito, G.; Di Marco, V.; Craxì, A.; Raimondo, G. Variability of reverse transcriptase and overlapping S gene in hepatitis B virus isolates from untreated and lamivudine-resistant chronic hepatitis B patients. Antivir 2009, 14, 649–654. [Google Scholar]
- Schildgen, O.; Olotu, C.; Funk, A.; Zollner, B.; Helm, M.; Rockstroh, J.K.; Sirma, H. Selection and Counterselection of the rtI233V Adefovir Resistance Mutation during Antiviral Therapy. J. Clin. Microbiol. 2010, 48, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liao, Y.; Chen, J.; Cai, B.; Su, Z.; Ying, B.; Lu, X.; Tao, C.; Wang, L. Erratum: Corrigendum: Epidemiology study of HBV genotypes and antiviral drug resistance in multi-ethnic regions from Western China. Sci. Rep. 2016, 6, 20451. [Google Scholar] [CrossRef] [Green Version]
- Shaw, T.; Bartholomeusz, A.; Locarnini, S. HBV drug resistance: Mechanisms, detection and interpretation. J. Hepatol. 2006, 44, 593–606. [Google Scholar] [CrossRef]
- Tenney, D.J.; Rose, R.E.; Baldick, C.J.; Pokornowski, K.A.; Eggers, B.J.; Fang, J.; Wichroski, M.J.; Xu, D.; Yang, J.; Wilber, R.B.; et al. Long-term monitoring shows hepatitis B virus resistance to entecavir in nucleoside-naïve patients is rare through 5 years of therapy. Hepatology 2009, 49, 1503–1514. [Google Scholar] [CrossRef]
- Nguyen, M.H.; Garcia, R.T.; Trinh, H.N.; Nguyen, H.A.; Nguyen, K.K.; Nguyen, L.H.; Levitt, B. Prevalence of hepatitis B virus DNA polymerase mutations in treatment-naïve patients with chronic hepatitis B. Aliment. Pharmacol. Ther. 2009, 30, 1150–1158. [Google Scholar] [CrossRef]
- Fan, J.; Wang, Y.; Xiong, H.; Guo, X.; Cheng, Y.-C. Impact of the rtI187V polymerase substitution of hepatitis B virus on viral replication and antiviral drug susceptibility. J. Gen. Virol. 2014, 95, 2523–2530. [Google Scholar] [CrossRef] [Green Version]
- Yamada, N.; Sugiyama, R.; Nitta, S.; Murayama, A.; Kobayashi, M.; Okuse, C.; Suzuki, M.; Yasuda, K.; Yotsuyanagi, H.; Moriya, K.; et al. Resistance mutations of hepatitis B virus in entecavir-refractory patients. Hepatol. Commun. 2017, 1, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-S.; Suh, D.J.; Lim, Y.; Jung, S.W.; Kim, K.M.; Lee, H.C.; Chung, Y.; Lee, Y.S.; Yoo, W.; Kim, S. Increased risk of adefovir resistance in patients with lamivudine-resistant chronic hepatitis B after 48 weeks of adefovir dipivoxil monotherapy. Hepatology 2006, 43, 1385–1391. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Total n (%) | Min-Max | Average | NA-Drug Resistance-Associated mt n | Other mt and wt n | Odds Ratio (95% CI) | p Value |
|---|---|---|---|---|---|---|---|
| Age (months) | 2–183 | 62.8 ± 3.8 | |||||
| ≤60 | 75 (52.8) | 48 | 27 | 0.812 (0.403–1.633) | 0.558 | ||
| >60 | 67 (47.2) | 46 | 21 | ||||
| Gender | |||||||
| Male | 80 (56.3) | 53 | 27 | 1.005 (0.499–2.027) | 0.988 | ||
| Female | 62 (43.7) | 41 | 21 | ||||
| AST (IU/L) | 15.5–2161.7 | 153.4 ± 25.3 | |||||
| ≤40 | 40 (28.2) | 28 | 12 | 1.273 (0.578–2.801) | 0.549 | ||
| >40 | 102 (71.8) | 66 | 36 | ||||
| ALT (IU/L) | 11.0–1849.7 | 160.1 ± 21.2 | |||||
| ≤40 | 53 (37.3) | 39 | 14 | 1.722 (0.817–3.629) | 0.151 | ||
| >40 | 89 (62.7) | 55 | 34 | ||||
| HBeAg (+/−) | |||||||
| Positive | 126 (88.7) | 82 | 44 | 0.621 (0.189–2.041) | 0.429 | ||
| Negative | 16 (11.3) | 12 | 4 | ||||
| HBV DNA level (IU/mL) | 6.105–1.109 | 6.108 | |||||
| <108 | 32 (22.5) | 21 | 11 | 0.968 (0.422–2.219) | 0.938 | ||
| ≥108 | 110 (77.5) | 73 | 37 | ||||
| Genotype HBV | |||||||
| B | 109 (76.8) | 68 | 41 | 0.447 (0.178–1.121) | 0.081 | ||
| C | 33 (23.2) | 26 | 7 |
| Characteristics | Total n | LMV Resistance-Associated mt n | ADF Resistance-Associated mt n | Odds Ratio (95% CI) | p Value |
|---|---|---|---|---|---|
| Age (months) | |||||
| ≤60 | 47 | 36 | 11 | 1.418 (0.555–3.623) | 0.464 |
| >60 | 43 | 30 | 13 | ||
| Gender | |||||
| Male | 51 | 37 | 14 | 0.911 (0.354–2.347) | 0.847 |
| Female | 39 | 29 | 10 | ||
| AST (IU/L) | |||||
| ≤40 | 26 | 20 | 06 | 1.304 (0.451–3.775) | 0.624 |
| >40 | 64 | 46 | 18 | ||
| ALT (IU/L) | |||||
| ≤40 | 36 | 26 | 10 | 0.910 (0.352–2.353) | 0.846 |
| >40 | 54 | 40 | 14 | ||
| HBeAg (+/−) | |||||
| Positive | 78 | 57 | 21 | 0.905 (0.223–3.666) | 1.000 |
| Negative | 12 | 09 | 03 | ||
| HBV DNA level (IU/mL) | |||||
| <108 | 19 | 14 | 05 | 1.023 (0.324–3.226) | 0.969 |
| ≥108 | 71 | 52 | 19 | ||
| HBV genotype | |||||
| B | 66 | 65 | 1 | 1495.000 (89.800–24889.032) | < 0.001 |
| C | 24 | 1 | 23 |
| Patients | Mutation Types | Resistance/Susceptibility to NA Drug | References | ||||
|---|---|---|---|---|---|---|---|
| n | % | LMV | ADF | TDF | ETV | ||
| Single-Point Mutation | |||||||
| 54 | 38.0 | rtV207M | R | S | S | S | [25] |
| 2 | 1.4 | rtV207L | R | S | S | S | [32] |
| 2 | 1.4 | rtS213T | R | S | S | S | [25] |
| 1 | 0.7 | rtL229V | R | S | S | S | [31] |
| 1 | 0.7 | rtP237T | S | R | S | S | [28,33] |
| 8 | 5.6 | rtN238A | S | R | S | S | [25] |
| 1 | 0.7 | rtN238K | S | R | S | S | [24] |
| 14 | 9.9 | rtN238T | S | R | S | S | [28,30] |
| 2 | 1.4 | rtS256G | R | S | S | S | [15,16,26] |
| Multi-Point Mutation | |||||||
| 2 | 1.4 | rtV207M + rtV207I | R | S | S | S | [25,27] |
| 1 | 0.7 | rtV207M + rtS213T | R | S | S | S | [25] |
| 1 | 0.7 | rtV207M + rtL229V | R | S | S | S | [25,31] |
| 1 | 0.7 | rtV207M + rtI233V | R | R | S | S | [25,29,34] |
| 1 | 0.7 | rtN238A + rtS256G | R | R | S | S | [15,16,25,26] |
| 1 | 0.7 | rtL180M + rtM204I + rtN238T | R | R | S | I | [19,23,28] |
| 1 | 0.7 | rtV207M + rtS213T + rtS256G | R | S | S | S | [15,16,25,26] |
| 1 | 0.7 | rtL180M + rtM204V + rtV207I + rtV207M | R | S | S | I | [19,23,25,27] |
| Total | |||||||
| 94 (66.2%) | 70 (49.3%) | 27 (19.0%) | 0 (0%) | 0 (0%) | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phung, T.T.B.; Chu, S.V.; Vu, S.T.; Pham, H.T.; Nguyen, H.M.; Nguyen, H.D.; Le, N.T.; Nguyen, D.V.; Truong, P.T.; Vu, V.T.T.; et al. COLD-PCR Method for Early Detection of Antiviral Drug-Resistance Mutations in Treatment-Naive Children with Chronic Hepatitis B. Diagnostics 2020, 10, 491. https://doi.org/10.3390/diagnostics10070491
Phung TTB, Chu SV, Vu ST, Pham HT, Nguyen HM, Nguyen HD, Le NT, Nguyen DV, Truong PT, Vu VTT, et al. COLD-PCR Method for Early Detection of Antiviral Drug-Resistance Mutations in Treatment-Naive Children with Chronic Hepatitis B. Diagnostics. 2020; 10(7):491. https://doi.org/10.3390/diagnostics10070491
Chicago/Turabian StylePhung, Thuy Thi Bich, Son Van Chu, Son Thien Vu, Hanh Thi Pham, Hang Minh Nguyen, Hoan Dang Nguyen, Ngan Thi Le, Dung Van Nguyen, Phuong Thai Truong, Van Thi Tuong Vu, and et al. 2020. "COLD-PCR Method for Early Detection of Antiviral Drug-Resistance Mutations in Treatment-Naive Children with Chronic Hepatitis B" Diagnostics 10, no. 7: 491. https://doi.org/10.3390/diagnostics10070491