Disabled Homolog 2 (DAB2) Protein in Tumor Microenvironment Correlates with Aggressive Phenotype in Human Urothelial Carcinoma of the Bladder

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. IHC of Human Samples and Murine Xenograft Tumors
2.2. Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-qPCR)
2.3. Microdissection of Cancerous and Stromal Tissues
2.4. Dual Immunofluorescent Staining of Human Bladder Cancer Tissue
2.5. Cell Lines and Reagents
2.6. Western Blot Analysis
2.7. Transfection of Small Interfering RNA (siRNA)
2.8. Cell Viability Assays
2.9. Matrigel Invasion and Migration Assay
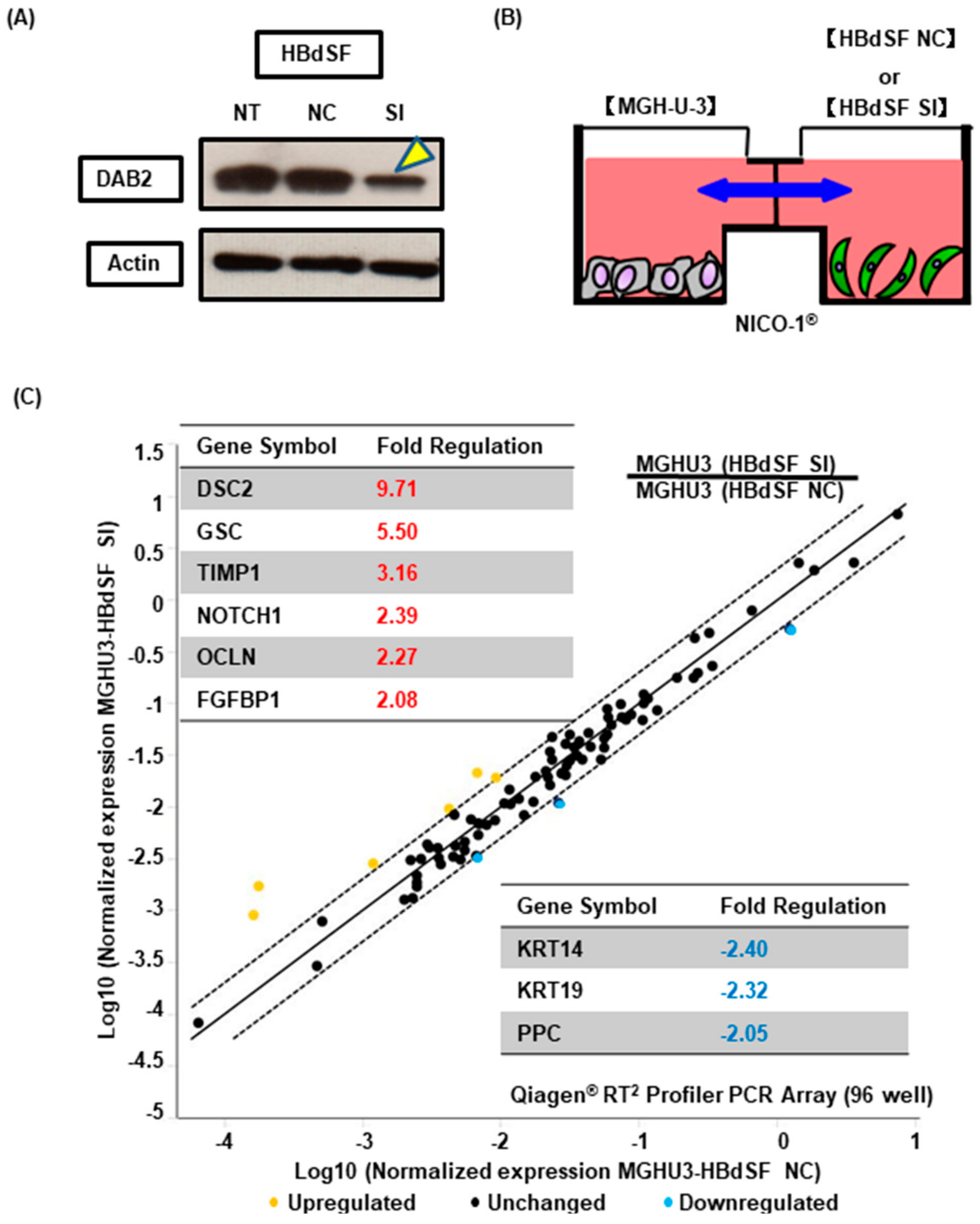
2.10. Interactive Co-Culture of Urothelial Cancer Cells and Bladder Stromal Fibroblasts
2.11. RT² Profiler™ PCR Array
2.12. Animals
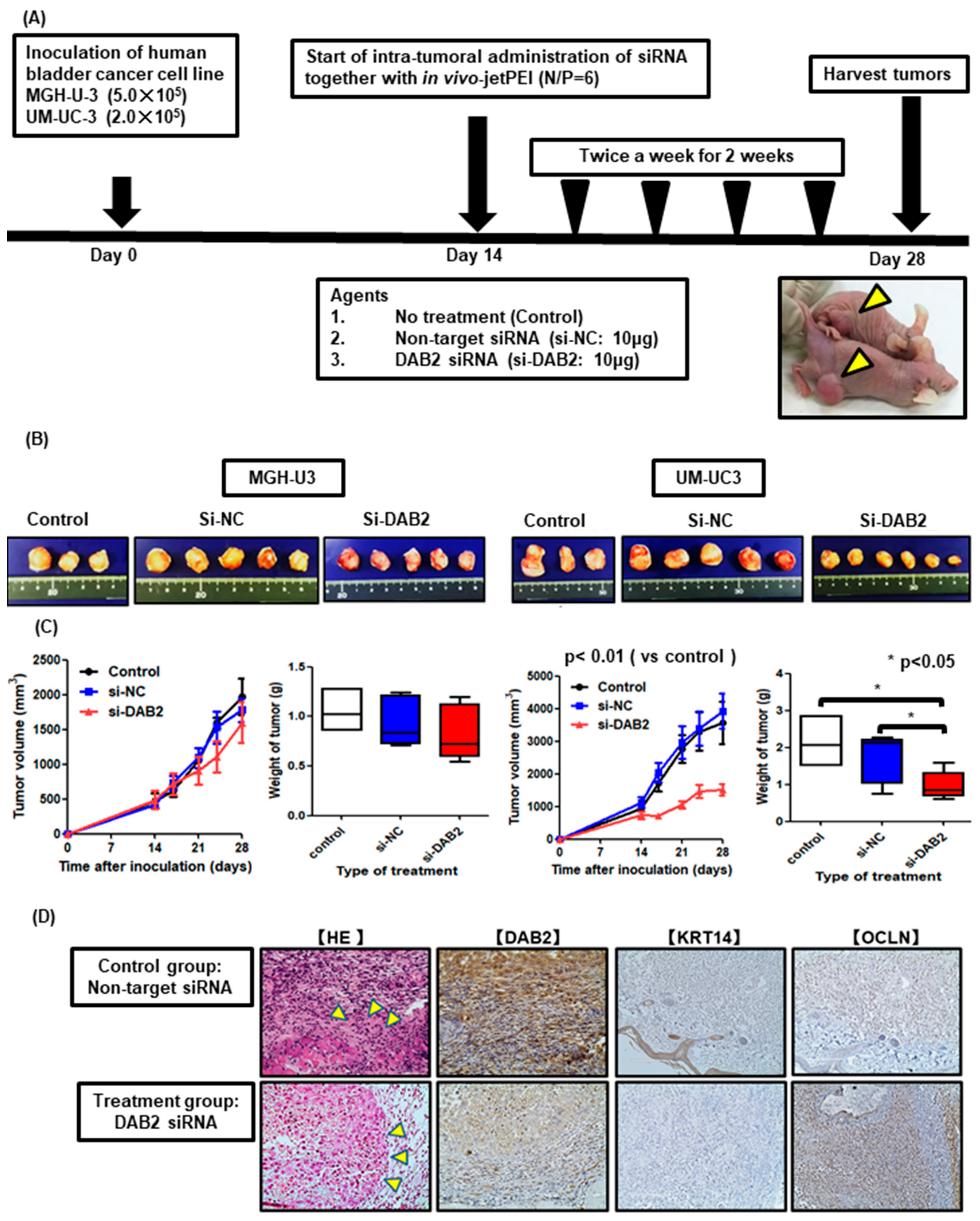
2.13. Xenograft Model and Intratumoral Treatment
2.14. Statistical Analysis
3. Results
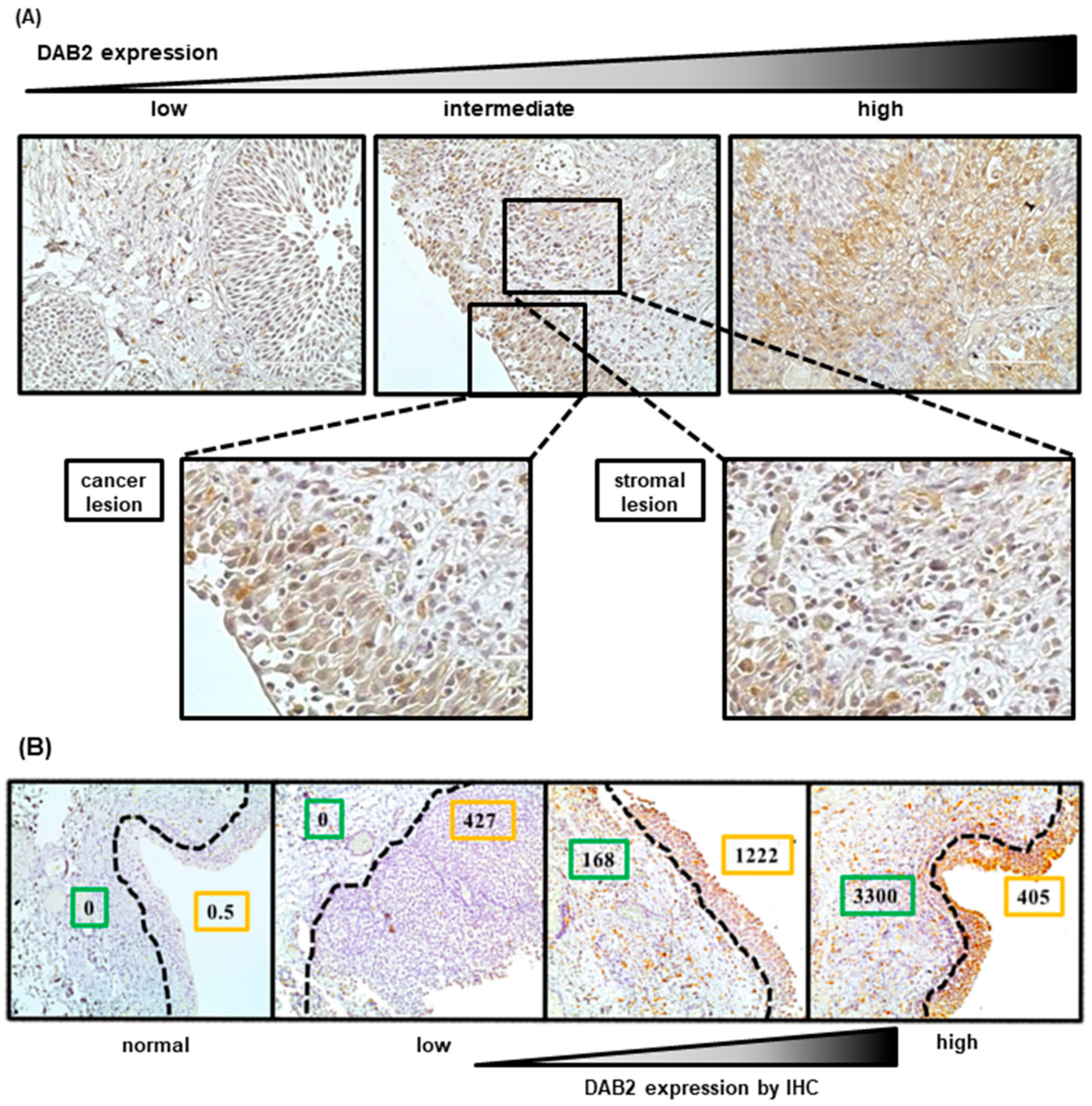
3.1. Association between DAB2 Expression and Clinicopathological Variables in Human Bladder Cancer
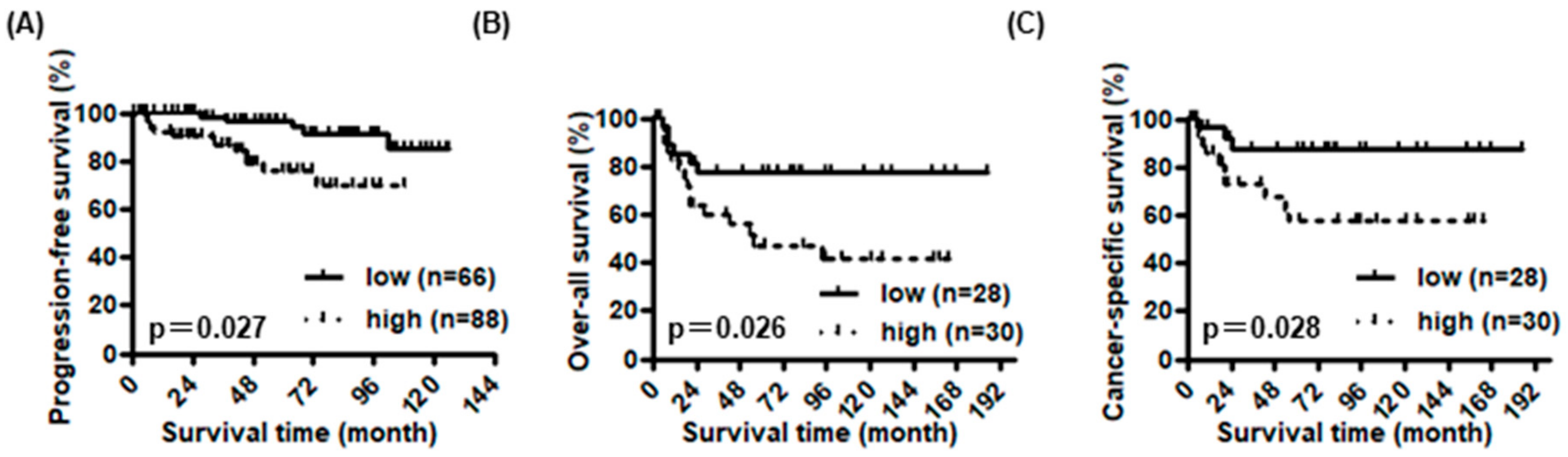
3.2. Prognostic Value of DAB2 Expression in Human Bladder Cancer
3.3. Evaluation of DAB2 Expression in Stromal Areas Adjacent to Cancerous Areas
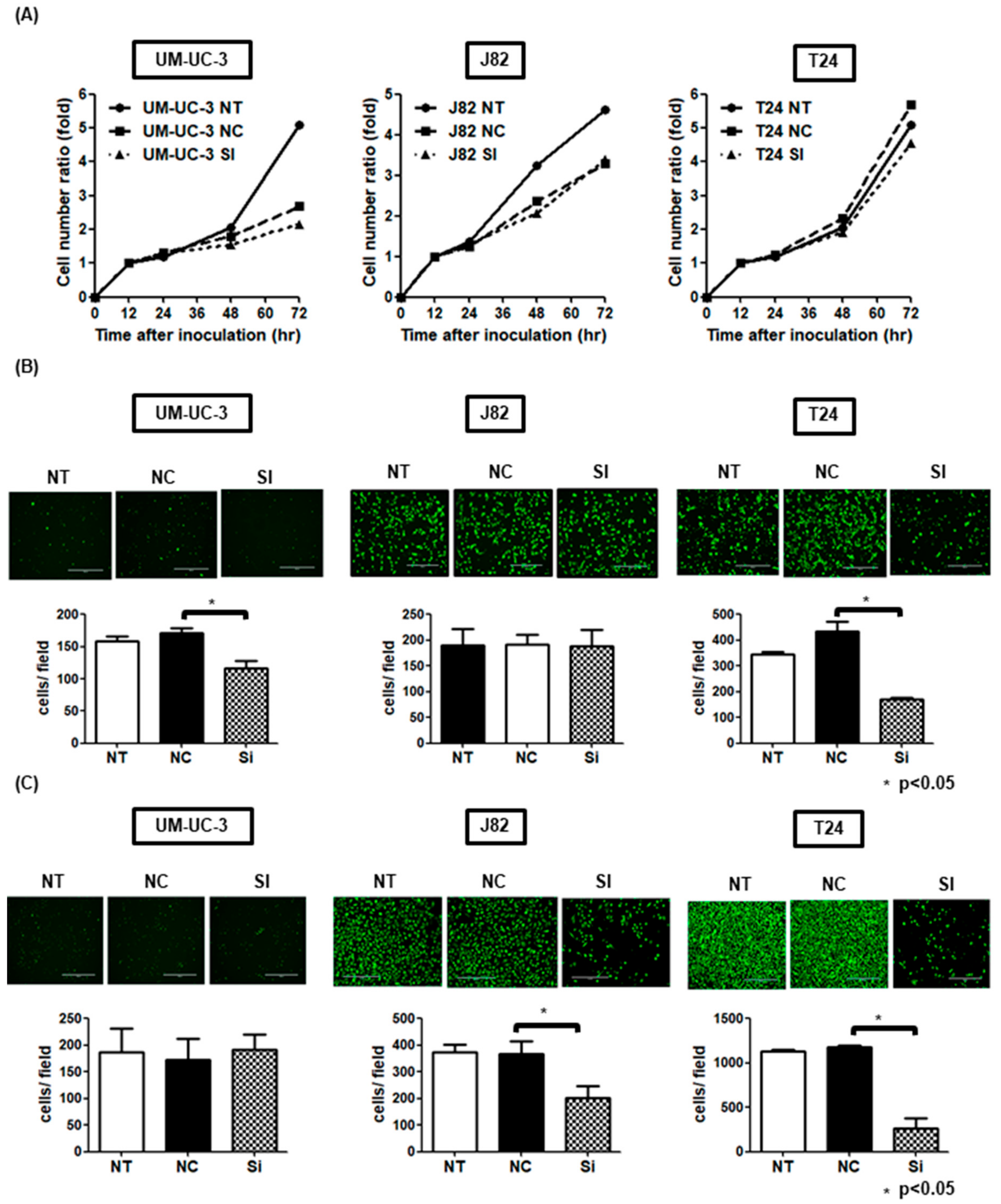
3.4. DAB2 Stimulates Tumor Migration and Invasion of Urothelial Cancer Cells
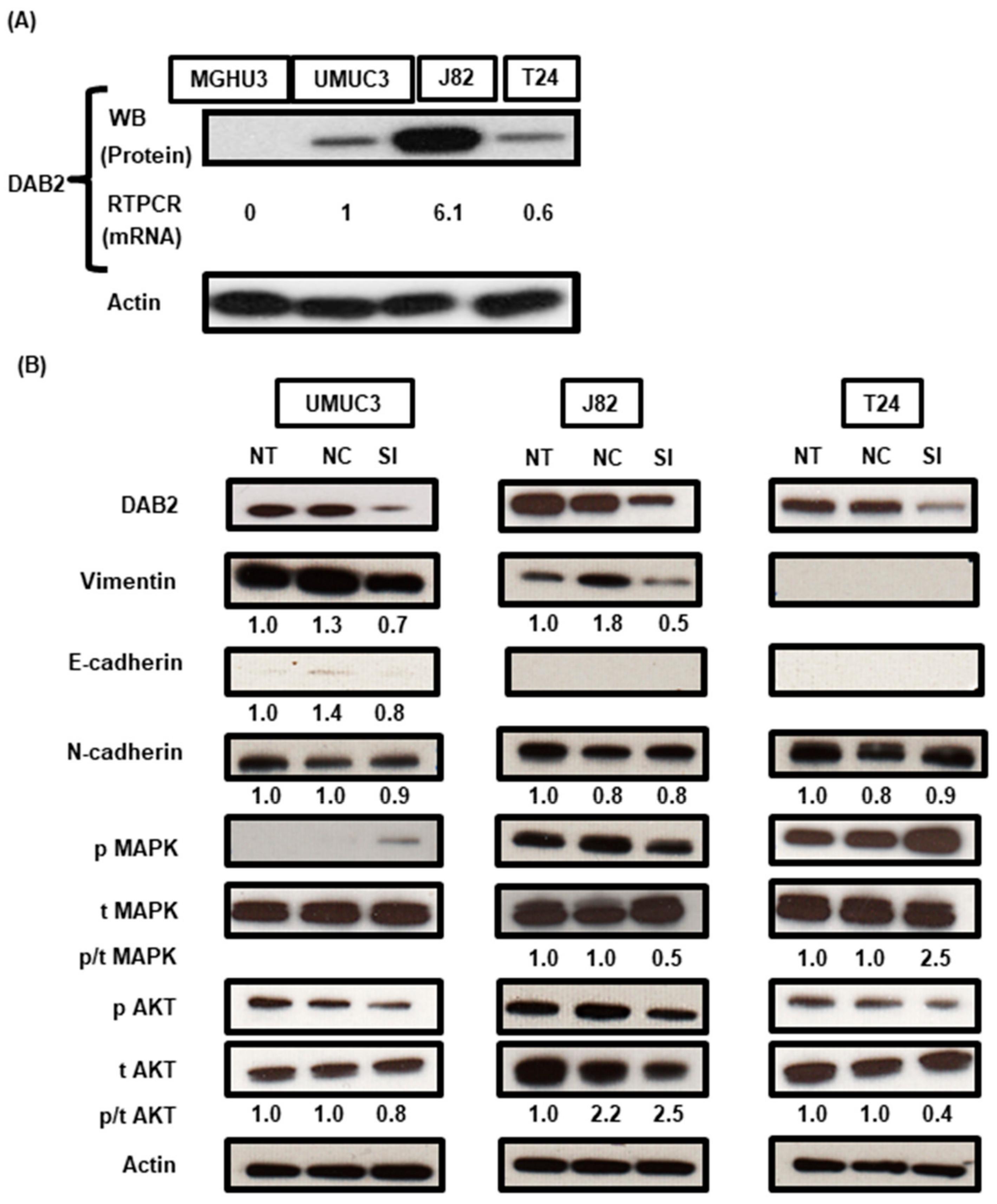
3.5. DAB2 Produced from Bladder Stromal Fibroblasts Correlates with Promotion of EMT in Human UCB
3.6. DAB2 siRNA Treatment Inhibits Tumor Growth and Invasion In Vivo
3.7. DAB2 siRNA Treatment Inhibits EMT of Bladder Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, M.; Fujimoto, K.; Anai, S.; Ohnishi, S.; Nakai, Y.; Inoue, T.; Matsumura, Y.; Tomioka, A.; Ikeda, T.; Tanaka, N.; et al. Clinical significance of heme oxygenase-1 expression in non-muscle-invasive bladder cancer. Urol. Int. 2010, 85, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Grossman, H.B.; Natale, R.B.; Tangen, C.M.; Speights, V.O.; Vogelzang, N.J.; Trump, D.L.; de Vere White, R.W.; Sarosdy, M.F.; Wood, D.P., Jr.; Raghavan, D.; et al. Neoadjuvant chemotherapy plus cystectomy compared with cystectomy alone for locally advanced bladder cancer. N. Engl. J. Med. 2003, 349, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Grigore, A.D.; Jolly, M.K.; Jia, D.; Farach-Carson, M.C.; Levine, H. Tumor budding: The Name is EMT. Partial EMT. J. Clin. Med. 2016, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, B.; Schaeffer, D.F.; Riddell, R.H.; Kirsch, R. Tumor budding in colorectal carcinoma: Time to take notice. Mod. Pathol. 2012, 25, 1315–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, R.E.; Gheiler, E.; Oskanian, P.; Tiguert, R.; Sakr, W.; Wood, D.P., Jr.; Pontes, J.E.; Grignon, D.J. Grading the invasive component of urothelial carcinoma of the bladder and its relationship with progression-free survival. Am. J. Surg. Pathol. 2000, 24, 980–987. [Google Scholar] [CrossRef]
- Miyake, M.; Gotoh, D.; Shimada, K.; Tatsumi, Y.; Nakai, Y.; Anai, S.; Torimoto, K.; Aoki, K.; Tanaka, N.; Konishi, N.; et al. Exploration of risk factors predicting outcomes for primary T1 high-grade bladder cancer and validation of the Spanish Urological Club for Oncological Treatment scoring model: Long-term follow-up experience at a single institute. Int. J. Urol. 2015, 22, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Krüger, S.; Noack, F.; Böhle, A.; Feller, A.C. Histologic tumor growth pattern is significantly associated with disease-related survival in muscle-invasive transitional cell carcinoma of the urinary bladder. Oncol. Rep. 2004, 12, 609–613. [Google Scholar] [CrossRef]
- Bircan, S.; Candir, O.; Kapucuoglu, N. The effect of tumor invasion patterns on pathologic stage of bladder urothelial carcinomas. Pathol. Oncol. Res. 2005, 11, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Miyake, M.; Hirao, S.; Mibu, H.; Tanaka, M.; Takashima, K.; Shimada, K.; Hirao, K. Clinical significance of subepithelial growth patterns in non-muscle invasive bladder cancer. BMC Urol. 2011, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Hori, S.; Miyake, M.; Ohnishi, S.; Tatsumi, Y.; Morizawa, Y.; Nakai, Y.; Anai, S.; Tanaka, N.; Fujimoto, K. Clinical significance of α- and β-Klotho in urothelial carcinoma of the bladder. Oncol. Rep. 2016, 36, 2117–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, M.; Hori, S.; Morizawa, Y.; Toritsuka, M.; Tatsumi, Y.; Ohnishi, S.; Shimada, K.; Furuya, H.; Khadka, V.S.; Deng, Y.; et al. Collagen type IV alpha 1 (COL4A1) and collagen type XIII alpha 1 (COL13A1) produced in cancer cells promote tumor budding at the invasion front in human urothelial carcinoma of the bladder. Oncotarget 2017, 8, 36099–36114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, S.C.; Wong, K.K.; Chan, R.K.; Lau, C.C.; Tsao, S.W.; Knapp, R.C.; Berkowitz, R.S. Molecular cloning of differentially expressed genes in human epithelial ovarian cancer. Gynecol. Oncol. 1994, 52, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Bohm, M.; Wieland, I.; Schmidt, C.; Rubben, H.; Allhoff, E.P. Loss of heterozygosity on chromosome 5p13-12 predicts adverse prognosis in advanced bladder cancer independent of tumor stage and grade. J. Urol. 2002, 168, 2655–2658. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Tang, J.; Xie, X. Frequent loss expression of dab2 and promotor hypermethylation in human cancers: A meta-analysis and systematic review. Pak. J. Med. Sci. 2014, 30, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Mok, S.C.; Chan, W.Y.; Wong, K.K.; Cheung, K.K.; Lau, C.C.; Ng, S.W.; Baldini, A.; Colitti, C.V.; Rock, C.O.; Berkowitz, R.S. DOC-2, a candidate tumor suppressor gene in human epithelial ovarian cancer. Oncogene 1998, 16, 2381–2387. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Z.; Sun, W.; Smith, E.; Cohen, C.; Sheng, Z.; Xu, X.X. Restoration of positioning control following disabled-2 expression in ovarian and breast tumor cells. Oncogene 2000, 19, 4847–4854. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Lin, C.Y.; Lee, Y.S.; Tsai, C.L.; Wei, P.C.; Hsueh, S.; Wu, T.I.; Tsai, C.N.; Wang, C.J.; Chao, A.S.; et al. Regulation of ovarian cancer progression by microRNA-187 through targeting disabled homolog-2. Oncogene 2012, 31, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Teckchandani, A.; Toida, N.; Goodchild, J.; Henderson, C.; Watts, J.; Wollscheid, B.; Cooper, J.A. Quantitative proteomics identifies a Dab2/integrin module regulating cell migration. J. Cell Biol. 2009, 186, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Chetrit, D.; Ziv, N.; Ehrlich, M. Dab2 regulates clathrin assembly and cell spreading. Biochem. J. 2009, 418, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijden, A.G.; Mengual, L.; Lozano, J.J.; Ingelmo-Torres, M.; Ribal, M.J.; Fernández, P.L.; Oosterwijk, E.; Schalken, J.A.; Alcaraz, A.; Witjes, J.A. A five-gene expression signature to predict progression in T1G3 bladder cancer. Eur. J. Cancer 2016, 64, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.; Miyake, M.; Tatsumi, Y.; Morizawa, Y.; Nakai, Y.; Ohnishi, S.; Ohnishi, K.; Iida, K.; Gotoh, D.; Tanaka, N.; et al. Gamma-Klotho exhibits multiple roles in tumor growth of human bladder cancer. Oncotarget 2018, 9, 19508–19524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimasaki, T.; Yamamoto, S.; Arisawa, T. Exosome Research and Co-culture Study. Biol. Pharm. Bull. 2018, 41, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Shimada, Y.; Kan, T.; David, S.; Cheng, Y.; Mori, Y.; Agarwal, R.; Paun, B.; Jin, Z.; Olaru, A.; et al. Pituitary tumor-transforming 1 increases cell motility and promotes lymph node metastasis in esophageal squamous cell carcinoma. Cancer Res. 2008, 68, 3214–3224. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.T.; Yang, L.H.; Li, Q.C.; Liu, S.L.; Liu, D.; Xie, X.M.; Wang, E.H. Disabled-2 and Axin are concurrently co-localized and under-expressed in lung cancers. Hum. Pathol. 2011, 42, 1491–1498. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Luo, W.; Howe, P.H. Dab2 stabilizes Axin and attenuates Wnt/β-catenin signaling by preventing protein phosphatase 1 (PP1)-Axin interactions. Oncogene 2009, 28, 2999–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhury, A.; Hussey, G.S.; Ray, P.S.; Jin, G.; Fox, P.L.; Howe, P.H. TGFβ-mediated phosphorylation of hnRNP E1 induces EMT via transcript-selective translational induction of Dab2 and ILEI. Nat. Cell Biol. 2010, 12, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Xu, X.X.; Yi, T.; Tang, B.; Lambeth, J.D. Disabled-2 (Dab2) is an SH3 domain-binding partner of Grb2. Oncogene 1998, 16, 1561–1569. [Google Scholar] [CrossRef] [Green Version]
- Prunier, C.; Howe, P.H. Disabled-2(Dab2) is required for transforming growth factor β-induced epithelial to mesenchymal transition (EMT). J. Biol. Chem. 2005, 280, 17540–17548. [Google Scholar] [CrossRef] [Green Version]
- Miyake, M.; Hori, S.; Morizawa, Y.; Tatsumi, Y.; Nakai, Y.; Anai, S.; Torimoto, K.; Aoki, K.; Tanaka, N.; Shimada, K.; et al. CXCL1-Mediated Interaction of Cancer Cells with Tumor-Associated Macrophages and Cancer-Associated Fibroblasts Promotes Tumor Progression in Human Bladder Cancer. Neoplasia 2016, 18, 636–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Zeisberg, M. Fibroblasts in Cancer. Nat. Rev. Cancer. 2006, 6, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Spivak-Kroizman, T.R.; Hostetter, G.; Posner, R.; Aziz, M.; Hu, C.; Demeure, M.J.; Von Hoff, D.; Hingorani, S.R.; Palculict, T.B.; Izzo, J.; et al. Hypoxia triggers hedgehog-mediated tumor-stromal interactions in pancreatic cancer. Cancer Res. 2013, 73, 3235–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Donato, M.; Cernera, G.; Migliaccio, A.; Castoria, G. Nerve Growth Factor Induces Proliferation and Aggressiveness in Prostate Cancer Cells. Cancers 2019, 11, 784. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Patients (%) | DAB2 Expression | p-Value † | |||
|---|---|---|---|---|---|
| Low | High | ||||
| Total | 212 | 116 | 96 | ||
| Age, years, median (range) | 71 (34–94) | 70 (34–94) | 71 (41–93) | 0.64 | |
| Gender | Male | 181 (86.8) | 86 | 95 | 0.19 |
| Female | 28 (13.2) | 17 | 11 | ||
| clinical T stage | Ta | 68 (32.1) | 48 | 20 | 0.014 |
| T1 | 81 (38.2) | 39 | 42 | ||
| Tis | 13 (6.1) | 5 | 8 | ||
| ≥T2 | 50 (23.6) | 24 | 26 | ||
| Grade | Low | 71 (33.5) | 55 | 16 | <0.001 |
| High | 141 (66.5) | 61 | 80 | ||
| Carcinoma in situ | Negative | 137 (64.6) | 91 | 46 | <0.001 |
| Positive | 75 (35.4) | 25 | 50 | ||
| Lymphovascular invasion (≥T1) | Negative | 65 (49.6) | 37 | 28 | 0.045 |
| Positive | 66 (50.4) | 26 | 40 | ||
| Histological variants | Yes | 33 (15.6) | 11 | 22 | <0.01 |
| No | 179 (84.4) | 105 | 74 | ||
| Lymph node status | N0 | 207 (97.6) | 114 | 93 | 0.83 |
| N1, N2 | 5 (2.3) | 2 | 3 | ||
| Tumor size | ≤3 cm | 150 (70.8) | 85 | 65 | 0.38 |
| >3 cm | 62 (29.2) | 31 | 31 | ||
| Number of tumor | single | 122 (57.5) | 70 | 52 | 0.36 |
| ≥2 | 90 (42.5) | 46 | 44 | ||
| Infiltration pattern (INF) | a | 86 (40.6) | 57 | 30 | 0.028 |
| b | 83 (39.2) | 40 | 42 | ||
| c | 43 (20.2) | 19 | 24 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itami, Y.; Miyake, M.; Ohnishi, S.; Tatsumi, Y.; Gotoh, D.; Hori, S.; Morizawa, Y.; Iida, K.; Ohnishi, K.; Nakai, Y.; et al. Disabled Homolog 2 (DAB2) Protein in Tumor Microenvironment Correlates with Aggressive Phenotype in Human Urothelial Carcinoma of the Bladder. Diagnostics 2020, 10, 54. https://doi.org/10.3390/diagnostics10010054
Itami Y, Miyake M, Ohnishi S, Tatsumi Y, Gotoh D, Hori S, Morizawa Y, Iida K, Ohnishi K, Nakai Y, et al. Disabled Homolog 2 (DAB2) Protein in Tumor Microenvironment Correlates with Aggressive Phenotype in Human Urothelial Carcinoma of the Bladder. Diagnostics. 2020; 10(1):54. https://doi.org/10.3390/diagnostics10010054
Chicago/Turabian StyleItami, Yoshitaka, Makito Miyake, Sayuri Ohnishi, Yoshihiro Tatsumi, Daisuke Gotoh, Shunta Hori, Yousuke Morizawa, Kota Iida, Kenta Ohnishi, Yasushi Nakai, and et al. 2020. "Disabled Homolog 2 (DAB2) Protein in Tumor Microenvironment Correlates with Aggressive Phenotype in Human Urothelial Carcinoma of the Bladder" Diagnostics 10, no. 1: 54. https://doi.org/10.3390/diagnostics10010054