
Maternal Stress Affects Fetal Growth but Not Developmental Instability in Rabbits


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species and Treatment
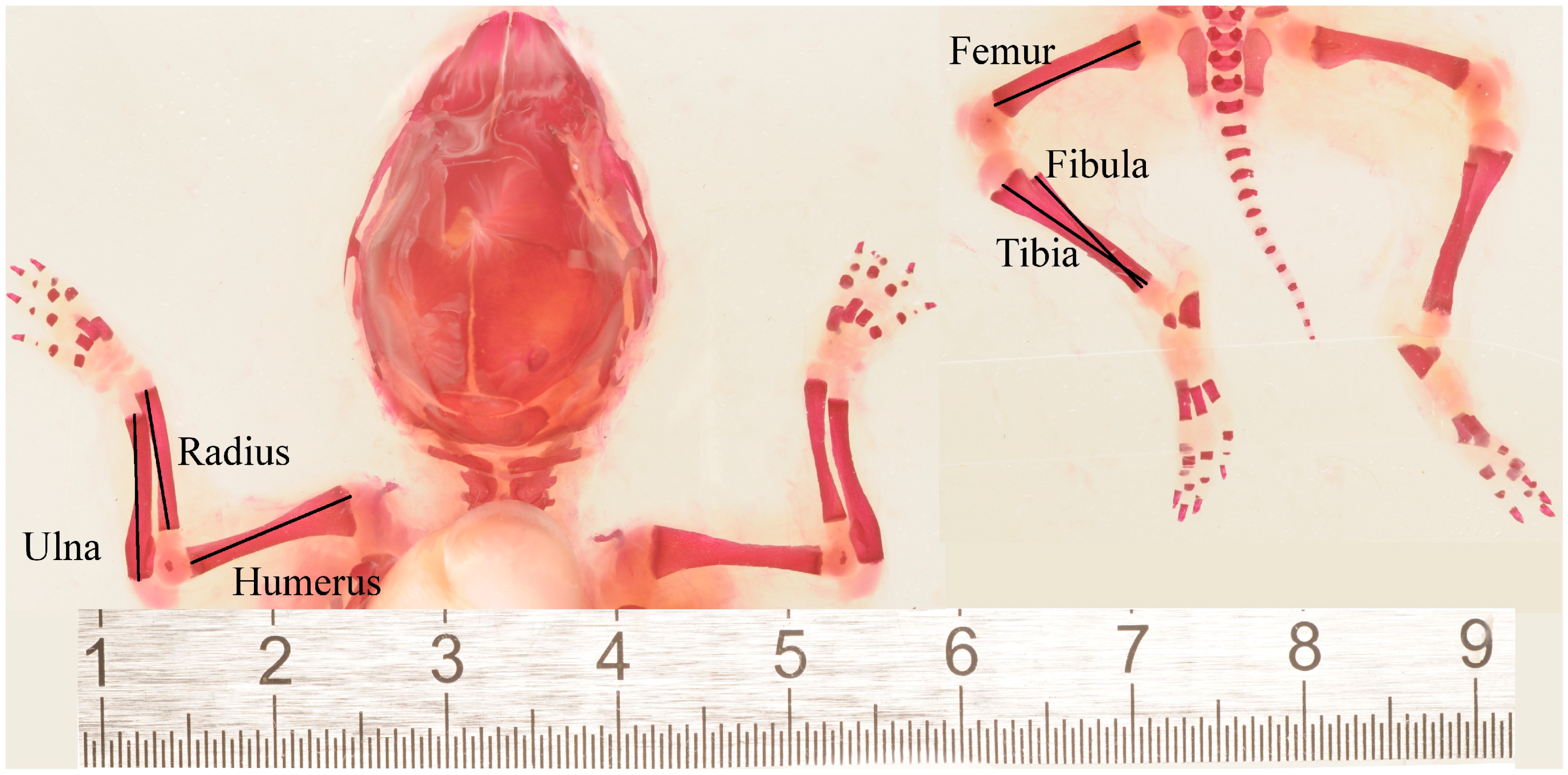
2.2. Limb Asymmetry
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Polak, M. Developmental Instability: Causes and Consequences; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Savriama, Y.; Klingenberg, C.P. Beyond bilateral symmetry: Geometric morphometrics for any type of symmetry. BMC Evol. Biol. 2011, 11, 280. [Google Scholar] [CrossRef] [PubMed]
- Lerner, I.M. Genetic Homeostasis; Edinburgh and London: Oliver & Boyd: New York, NY, USA, 1954. [Google Scholar]
- Rasmussen, M. Frequency of morphological deviants as a criterion of developmental stability. Hereditas 1960, 46, 511–535. [Google Scholar] [CrossRef]
- Lens, L.; van Dongen, S.; Kark, S.; Matthysen, E. Fluctuating asymmetry as an indicator of fitness: Can we bridge the gap between studies? Biol. Rev. 2002, 77, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, S. Fluctuating asymmetry and developmental instability in evolutionary biology: Past, present, future. J. Evol. Biol. 2006, 19, 1727–1743. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Klingenberg, C.P. The genetics and evolution of fluctuating asymmetry. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 1–21. [Google Scholar] [CrossRef]
- Pertoldi, C.; Kristensen, T.N.; Andersen, D.H.; Loeschcke, V. Developmental instability as an estimator of genetic stress. Heredity 2006, 96, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Parasitism and developmental instability of hosts: A review. Oikos 1999, 77, 189–196. [Google Scholar] [CrossRef]
- Lens, L.; van Dongen, S.; Matthysen, E. Fluctuating asymmetry as an early warning system in the critically endangered Taita Thrush. Conserv. Biol. 2002, 16, 479–487. [Google Scholar] [CrossRef]
- Hoffman, A.A.; Woods, R.E. Associating environ- mental stress with developmental stability: Problems and patterns. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 387–401. [Google Scholar]
- Parson, P.A. Maternal age and developmental variability. J. Exp. Biol. 1962, 39, 251–260. [Google Scholar]
- Livshits, G.; Davidi, L.; Kobyliansky, E.; Ben-Amital, D.; Levi, Y.; Merlob, P. Decreased developmental stability as assessed by fluctuating asymmetry of morphometric traits in preterm infants. Am. J. Med. Genet. 1988, 29, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Rosen, V.C. Effects of maternal body morphology, morning sickness, gestational diabetes and hypertension on fluctuating asymmetry in young woman. Evol. Hum. Behav. 2001, 22, 373–384. [Google Scholar] [CrossRef]
- Van Dongen, S.; Gangestad, S.W. Human fluctuating asymmetry in relation to health and quality: A meta-analysis. Evol. Hum. Behav. 2011, 32, 380–398. [Google Scholar] [CrossRef]
- Thornhill, R.; Møller, A.P. Developmental stability, disease and medicine. Biol. Rev. 1997, 72, 497–548. [Google Scholar] [CrossRef] [PubMed]
- Wilber, E.; Newell-Morris, L.; Streissguth, A.P. Dermatoglyphic asymmetry in fetal alcohol syndrome. Biol. Neonate 1993, 64, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kieser, J.A.; Groeneveld, H.T.; Da Silva, P.C. Dental asymmetry, maternal obesity and smoking. Am. J. Phys. Anthropol. 1997, 102, 133–139. [Google Scholar] [CrossRef]
- Van Dongen, S.; Wijnaendts, L.C.D.; Ten Broek, C.M.A.; Galis, F. Fluctuating asymmetry does not consistently reflect severe developmental disorders in human foetuses. Evolution 2009, 63, 1832–1844. [Google Scholar] [CrossRef] [PubMed]
- King, S.; Mancini-Marïe, A.; Brunet, A.; Walker, E.; Meaney, M.J.; Laplante, D.P. Prenatal maternal stress from a natural disaster predicts dermatoglyphic asymmetry in humans. Dev. Psychopathol. 2009, 21, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Benderlioglu, Z. Fluctuating asymmetry and steroid hormones: A review. Symmetry 2010, 2, 541–553. [Google Scholar] [CrossRef]
- Eriksen, M.S.; Espmark, Å.M.; Poppe, T.; Braastad, B.O.; Salte, R.; Bakken, M. Fluctuating asymmetry in farmed Atlantic salmon (Salmo salar) juveniles: Also a maternal matter? Environ. Biol. Fishes 2006, 81, 87–99. [Google Scholar] [CrossRef]
- Eriksen, M.S.; Haug, A.; Torjesen, P.A.; Bakken, M. Prenatal exposure to corticosterone impairs embryonic development and increases fluctuating asymmetry in chickens (Gallus gallus domesticus). Br. Poult. Sci. 2003, 44, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Satterlee, D.G.; Hester, A.; Leray, K.; Schmidt, J.B. Influences of maternal corticosterone and selection for contrasting adrenocortical responsiveness in Japanese quail on developmental instability of female progeny. Poult. Sci. 2008, 87, 1504–1509. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, M.; McCormick, M.I. Hormonally mediated maternal effects shape offspring survival potential in stressful environments. Oecologia 2009, 160, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Helle, S.; Suorsa, P.; Huhta, E.; Hakkarainen, H. Fluctuating feather asymmetry in relation to corticosterone levels is seks-dependent in Eurasian treecreeper (Certhia familiaris) nestlings. Biol. Lett. 2010, 6, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.M. Relationships between developmental stability and fitness—Application for conservation biology. Conserv. Biol. 1995, 9, 18–24. [Google Scholar] [CrossRef]
- Breno, M.; Bots, J.; de Schaepdrijver, L.; van Dongen, S. Fluctuating asymmetry as risk marker for stress and structural defects in a toxicological experiment. Birth Defects Res. B Dev. Reprod. Toxicol. 2013, 98, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Dawson, A.B. A note on the staining of skeleton of cleared specimens with alizarin red S. Stain Technol. 1926, 1, 123–124. [Google Scholar] [CrossRef]
- Breno, M.; Bots, J.; van Dongen, S. Between-family variation and quantitative genetics of developmental instability of long bones in rabbit fetuses. Biol. J. Linn. Soc. 2013, 109, 33–42. [Google Scholar] [CrossRef]
- Breno, M.; Bots, J.; van Dongen, S. Heritabilities of directional asymmetry in the fore- and hindlimbs of rabbit fetuses. PLoS ONE 2013, 8, e76358. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Van Dongen, S.; Molenberghs, G.; Matthysen, E. The statistical analysis of fluctuating asymmetry: REML estimation of a mixed regression model. J. Evol. Biol. 1999, 12, 94–102. [Google Scholar] [CrossRef]
- Carney, E.W.; Kimmel, C.A. Interpretation of skeletal variations for human risk assessment: Delayed ossification and wavy ribs. Birth Defect Res. B Dev. Reprod. Toxicol. 2007, 80, 473–496. [Google Scholar] [CrossRef] [PubMed]
- Baayen, R.H.; Davidson, D.J.; Bates, D.M. Mixed-effects modelling with crossed random effects for subjects and items. J. Mem. Lang. 2008, 59, 390–412. [Google Scholar] [CrossRef]
- The R project for statistical computing. Available online: http://www-r-project.org (accessed on 17 August 2016).
- Cappon, G.D.; Fleeman, T.L.; Chapin, R.E.; Hurt, M.E. Effects of feed restriction during organogenesis on embryo-fetal development in rabbit. Birth Defect Res. B Dev. Reprod. Toxicol. 2005, 74, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A. Spontaneous malformations of the New Zealand White rabbit: The background to safety evaluation tests. Lab. Anim. 1968, 2, 195–206. [Google Scholar] [CrossRef]
- Khera, K.S. Maternal toxicity in humans and animals: Effects on fetal development and criteria for detection. Teratig. Carcinog. Mutagen. 1987, 7, 287–295. [Google Scholar] [CrossRef]
- Wingfield, J.C.; Maney, D.L.; Breuner, C.W.; Jacobs, J.D.; Lynn, S.; Ramenofsky, M.; Richardson, R.D. Ecological bases of hormone-behavior interactions: The ‘emergency life history stage’. Am. Zool. 1998, 38, 191–206. [Google Scholar] [CrossRef]
- Painter, R.C.; Roseboom, T.J.; de Rooij, S.R. Long-term effects of prenatal stress and glucocorticoid exposure. Birth Defects Res. C Embryo Today 2012, 96, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Pravosudov, V.V.; Kitaysky, A.S. Effects of nutritional restrictions during post-hatching development on adrenocortical function in western scrub-jays (Aphelocoma californica). Gen. Comp. Endocrinol. 2006, 145, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Shine, R. A New Hypothesis for the evolution of viviparity in reptiles. Am. Nat. 1995, 145, 809–823. [Google Scholar] [CrossRef]
- Shine, R. Does viviparity evolve in cold climate reptiles because pregnant females maintain stable (not high) body temperatures? Evolution 2004, 58, 1809–1808. [Google Scholar] [CrossRef] [PubMed]
- Dimasuay, K.G.; Boeuf, P.; Powell, T.L.; Jansson, T. Placental responses to changes in the maternal environment determine fetal growth. Front. Physiol. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Newell-Morris, L.L.; Fahrenbruch, C.E.; Sackett, G.P. Prenatal psychological stress, dermatoglyphic asymmetry and pregnancy outcome in the pigtailed macaque (Macaca nemestrina). Biol. Neonate 1989, 56, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Hallgrímsson, B. Fluctuating asymmetry in the mammalian skeleton—Evolutionary and develop-mental implications. Evol. Biol. 1998, 30, 187–251. [Google Scholar]
- Gonzalez, P.N.; Lotto, F.P.; Hallgrímsson, B. Canalization and developmental instability of the fetal skull in a mouse model of maternal nutritional stress. Am. J Phys. Anthropol. 2014, 154, 544–553. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Trait | σ2FA | σ2ME-photo. | σ2ME-meas. | %ME | DA |
|---|---|---|---|---|---|
| Humerus | 0.0239 | 0.0105 | 0.0098 | 46 | 0.0020 |
| Ulna | 0.0648 | 0.0299 | 0.0134 | 40 | −0.0013 |
| Radius | 0.0144 | 0.0037 | 0.0103 | 49 | 0.0018 |
| Femur | 0.0166 | 0.0019 | 0.0100 | 41 | −0.0001 |
| Tibia | 0.0323 | 0.0136 | 0.0098 | 42 | −0.0009 |
| Fibula | 0.0158 | 0.0003 | 0.0209 | 57 | −0.0008 |
| Trait | Control | Low | Medium | High | F |
|---|---|---|---|---|---|
| Maternal food consumption (g) | 5.65 ± 0.87 | 5.38 ± 1.62 | 4.75 ± 1.05 | 4.28 ± 1.69 | 3.96 |
| Maternal weight gain (g) | −0.03 ± 0.04 | −0.05 ± 0.03 | −0.07 ± 0.04 | −0.11 ± 0.06 | 10.445 |
| Litter size (n) | 9 ± 2.55 | 9.1 ± 2.13 | 9.05 ± 1.51 | 9.19 ± 1.83 | 0.0263 |
| Pre-implantation loss (%) | 0.17 ± 0.21 | 0.16 ± 0.16 | 0.17 ± 0.13 | 0.09 ± 0.1 | 1.1372 |
| Post-implantation loss (%) | 0.06 ± 0.09 | 0.08 ± 0.1 | 0.05 ± 0.07 | 0.11 ± 0.11 | 1.5164 |
| Average pup weight (g) | 31.4 ± 2.8 | 30.11 ± 3.45 | 27.73 ± 3.53 | 26.22 ± 4.4 | 11.275 |
| Treatment | pno | mro | 13rib | 1_13rib | R_13rib | rb | sz | sbf | sba | Sbr |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0.02 | 0.12 | 0.56 | 0.13 | 0.09 | 0.13 | 0.01 | 0.03 | 0.07 | 0.11 |
| Low | 0.02 | 0.21 | 0.57 | 0.09 | 0.09 | 0.25 | 0.02 | 0.02 | 0.05 | 0.04 |
| Medium | 0.08 | 0.28 | 0.59 | 0.1 | 0.09 | 0.35 | 0.06 | 0.12 | 0.05 | 0.09 |
| High | 0.15 | 0.49 | 0.78 | 0.09 | 0.03 | 0.35 | 0.14 | 0.25 | 0.06 | 0.07 |
| χ23 | 30.84 | 63.67 | 23.99 | 1.71 | 7.35 | 30.32 | 33.75 | 64.85 | 0.78 | 5.26 |
| Trait | Control | Low | Medium | High | χ23 |
|---|---|---|---|---|---|
| Humerus | 0.0032 | 0.0032 | 0.0031 | 0.0038 | 6.38 |
| Ulna | 0.0059 | 0.0054 | 0.0051 | 0.0056 | 1.86 |
| Radius | 0.0021 | 0.0027 | 0.0026 | 0.0027 | 8.80 |
| Femur | 0.0028 | 0.0027 | 0.0031 | 0.0028 | 2.09 |
| Tibia | 0.0039 | 0.0036 | 0.0039 | 0.0036 | 0.61 |
| Fibula | 0.0026 | 0.0024 | 0.0027 | 0.0031 | 1.63 |
| Fore limb | 0.0038 | 0.0037 | 0.0035 | 0.0040 | 0.51 |
| Hind Limb | 0.0031 | 0.0029 | 0.0033 | 0.0032 | 1.63 |
| Overall | 0.0034 | 0.0033 | 0.0034 | 0.0036 | 2.46 |
| Trait | minor | pno | mro | 13rib | rb | sz | sbf | sbar |
|---|---|---|---|---|---|---|---|---|
| Humerus | −0.0002 | −0.0003 | 0.0002 | 0.0005 | −0.0005 | −0.0007 | 0.0003 | −0.0003 |
| Ulna | 0.0002 | −0.0011 | 0.0000 | 0.0000 | 0.0001 | 0.0004 | −0.0007 | 0.0000 |
| Radius | 0.0003 | 0.0002 | 0.0004 | 0.0000 | 0.0000 | −0.0004 | 0.0008 | 0.0001 |
| Femur | 0.0001 | 0.0013 | 0.0003 | −0.0003 | 0.0001 | −0.0005 | 0.0000 | −0.0003 |
| Tibia | −0.0002 | −0.0005 | −0.0002 | −0.0006 | −0.0001 | 0.0004 | 0.0010 | 0.0000 |
| Fibula | −0.0001 | −0.0002 | 0.0001 | 0.0001 | −0.0003 | 0.0003 | −0.0003 | 0.0001 |
| Fore limb | 0.0000 | −0.0004 | 0.0002 | 0.0002 | −0.0002 | −0.0004 | 0.0001 | 0.0000 |
| Hind Limb | 0.0001 | 0.0002 | 0.0000 | −0.0002 | −0.0001 | 0.0002 | 0.0003 | −0.0001 |
| Overall | 0.0000 | −0.0001 | 0.0001 | 0.0000 | −0.0001 | −0.0001 | 0.0002 | −0.0001 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bots, J.; Breno, M.; De Schaepdrijver, L.; Van Dongen, S. Maternal Stress Affects Fetal Growth but Not Developmental Instability in Rabbits. Symmetry 2016, 8, 101. https://doi.org/10.3390/sym8100101
Bots J, Breno M, De Schaepdrijver L, Van Dongen S. Maternal Stress Affects Fetal Growth but Not Developmental Instability in Rabbits. Symmetry. 2016; 8(10):101. https://doi.org/10.3390/sym8100101
Chicago/Turabian StyleBots, Jessica, Matteo Breno, Luc De Schaepdrijver, and Stefan Van Dongen. 2016. "Maternal Stress Affects Fetal Growth but Not Developmental Instability in Rabbits" Symmetry 8, no. 10: 101. https://doi.org/10.3390/sym8100101