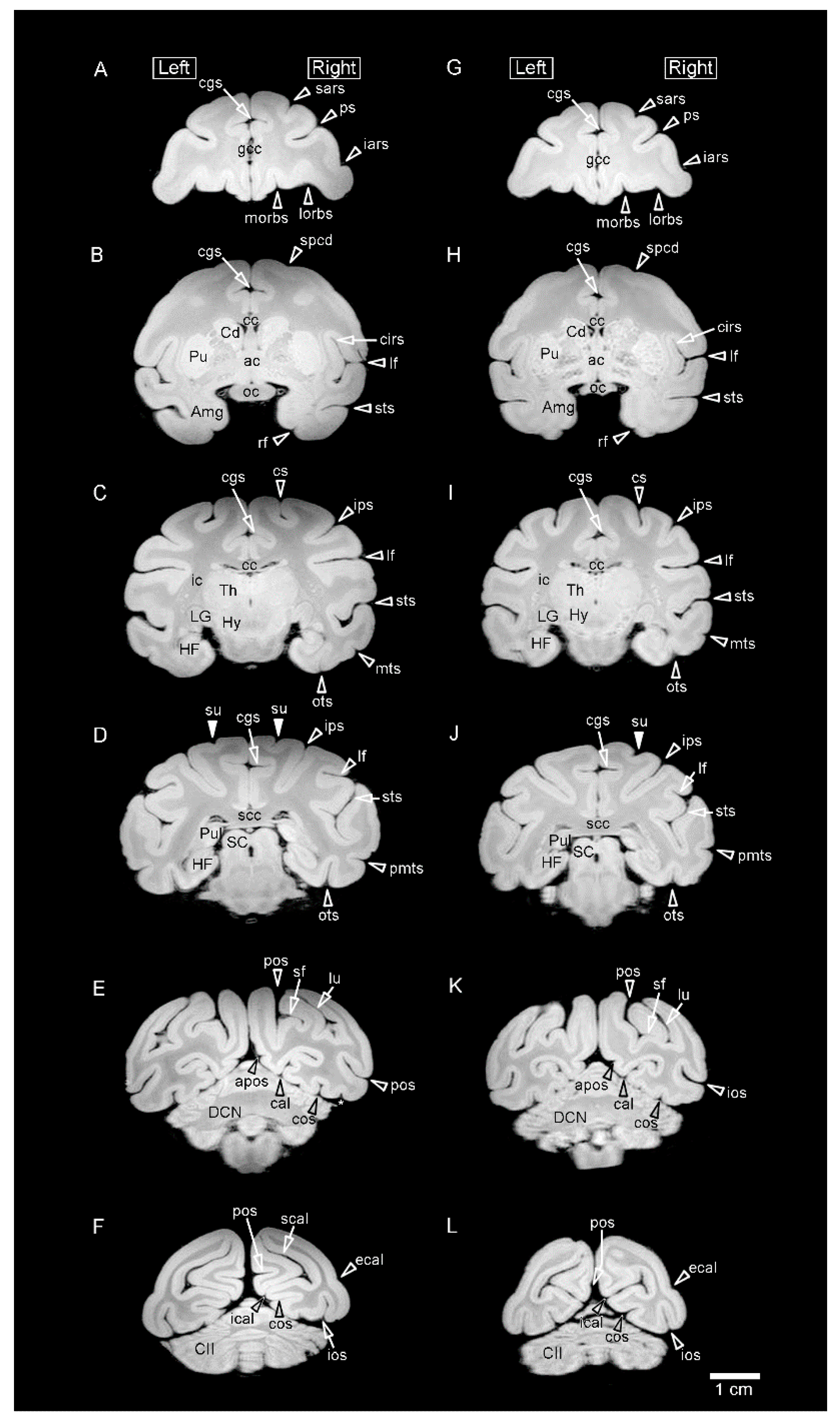
Figure 1.
Representative coronal (axial) MR images (using a RARE sequence with a short TR and the minimum TE settings) obtained from fixed brains of sexually mature cynomolgus monkeys. (A–F) Coronal MR images of the male brain at rostral to caudal levels. (G–L) Coronal MR images of the female brain at rostral to caudal levels. MR images on the same lines are identical levels seen at: (A,G) the rostral end of the genu of corpus callosum (gcc); (B,H) the anterior commissure; (C,I) the lateral geniculate nucleus (LG) of the thalamus; (D,J) the splenium of corpus callosum (scc); (E,K) the deep cerebellar nucleus (DCN); and (F,L) the crus II of ansiform lobule (CII) of cerebellum. Amg, amygdala; apos, anterior parietooccipital sulcus; cal, calcarine sulcus; cgs, cingulate sulcus; cc, corpus callosum; Cd, caudate nucleus; cirs, circular sulcus; cos, collateral sulcus; cs, central sulcus; ecal, external calcarine sulcus; HF, hippocampal formation; Hy, hypothalamus; iars, inferior ram of arcuate sulcus; ic, internal capsule; ical, inferior calcarine sulcus; ios, inferior occipital sulcus; ips, intraparietal sulcus; lf, lateral fissure; lorb, lateral orbital sulcus; lu, lunate sulcus; morb, medial orbital sulcus; mts, middle temporal sulcus; oc, optic chiasma; ots, occipitotemporal sulcus; pmts, posterior middle temporal sulcus; pos, parietooccipital sulcus; ps, principal sulcus; Pu, putamen; Pul, pulvinar of the thalamus; rf, rhinal fissure; sars, superior ram of arcuate sulcus; SC, superior colliculus; scal, superior calcarine sulcus; sf, simian fossa; spcd, superior precentral dimple; sts, superior temporal sulcus; su, superior postcentral dimple; Th, thalamus.
Figure 1.
Representative coronal (axial) MR images (using a RARE sequence with a short TR and the minimum TE settings) obtained from fixed brains of sexually mature cynomolgus monkeys. (A–F) Coronal MR images of the male brain at rostral to caudal levels. (G–L) Coronal MR images of the female brain at rostral to caudal levels. MR images on the same lines are identical levels seen at: (A,G) the rostral end of the genu of corpus callosum (gcc); (B,H) the anterior commissure; (C,I) the lateral geniculate nucleus (LG) of the thalamus; (D,J) the splenium of corpus callosum (scc); (E,K) the deep cerebellar nucleus (DCN); and (F,L) the crus II of ansiform lobule (CII) of cerebellum. Amg, amygdala; apos, anterior parietooccipital sulcus; cal, calcarine sulcus; cgs, cingulate sulcus; cc, corpus callosum; Cd, caudate nucleus; cirs, circular sulcus; cos, collateral sulcus; cs, central sulcus; ecal, external calcarine sulcus; HF, hippocampal formation; Hy, hypothalamus; iars, inferior ram of arcuate sulcus; ic, internal capsule; ical, inferior calcarine sulcus; ios, inferior occipital sulcus; ips, intraparietal sulcus; lf, lateral fissure; lorb, lateral orbital sulcus; lu, lunate sulcus; morb, medial orbital sulcus; mts, middle temporal sulcus; oc, optic chiasma; ots, occipitotemporal sulcus; pmts, posterior middle temporal sulcus; pos, parietooccipital sulcus; ps, principal sulcus; Pu, putamen; Pul, pulvinar of the thalamus; rf, rhinal fissure; sars, superior ram of arcuate sulcus; SC, superior colliculus; scal, superior calcarine sulcus; sf, simian fossa; spcd, superior precentral dimple; sts, superior temporal sulcus; su, superior postcentral dimple; Th, thalamus.
![Symmetry 15 01164 g001]()
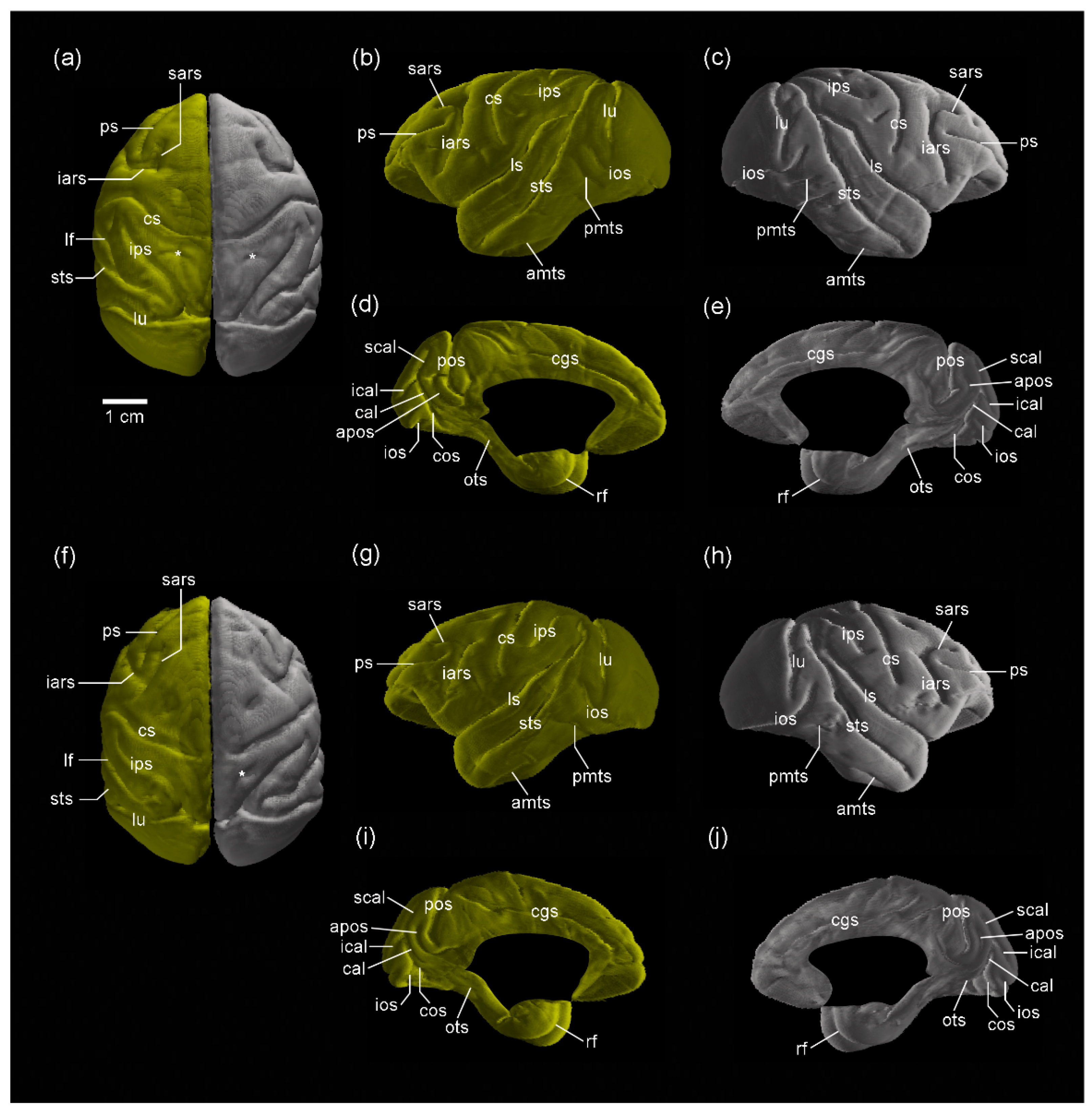
Figure 2.
Three-dimensional volume-rendered images of the cerebral cortex of sexually mature cynomolgus monkeys. (a–e) Dorsal view of the left and right hemispheres, lateral view of the left hemisphere, lateral view of the right hemisphere, medial view of the left hemisphere, and medial view of the right hemisphere of males. (f–j) Dorsal view of the left and right hemispheres, lateral view of the left hemisphere, lateral view of the right hemisphere, medial view of the left hemisphere, and medial view of the right hemisphere of females. The superior postcentral dimple (*) is seen in both the left and right hemispheres in males, amts, anterior middle temporal sulcus; apos, anterior parietooccipital sulcus; cal, calcarine sulcus; cgs, cingulate sulcus; cos, collateral sulcus; cs, central sulcus; iars, inferior ram of arcuate sulcus; ical, inferior calcarine sulcus; ios, inferior occipital sulcus; ips, intraparietal sulcus; lf, lateral fissure; lu, lunate sulcus; ots, occipitotemporal sulcus; pmts, posterior middle temporal sulcus; pos, parietooccipital sulcus; ps, principal sulcus; rf, rhinal fissure; sars, superior ram of arcuate sulcus; scal, superior calcarine sulcus; sts, superior temporal sulcus; *, superior postcentral dimple.
Figure 2.
Three-dimensional volume-rendered images of the cerebral cortex of sexually mature cynomolgus monkeys. (a–e) Dorsal view of the left and right hemispheres, lateral view of the left hemisphere, lateral view of the right hemisphere, medial view of the left hemisphere, and medial view of the right hemisphere of males. (f–j) Dorsal view of the left and right hemispheres, lateral view of the left hemisphere, lateral view of the right hemisphere, medial view of the left hemisphere, and medial view of the right hemisphere of females. The superior postcentral dimple (*) is seen in both the left and right hemispheres in males, amts, anterior middle temporal sulcus; apos, anterior parietooccipital sulcus; cal, calcarine sulcus; cgs, cingulate sulcus; cos, collateral sulcus; cs, central sulcus; iars, inferior ram of arcuate sulcus; ical, inferior calcarine sulcus; ios, inferior occipital sulcus; ips, intraparietal sulcus; lf, lateral fissure; lu, lunate sulcus; ots, occipitotemporal sulcus; pmts, posterior middle temporal sulcus; pos, parietooccipital sulcus; ps, principal sulcus; rf, rhinal fissure; sars, superior ram of arcuate sulcus; scal, superior calcarine sulcus; sts, superior temporal sulcus; *, superior postcentral dimple.
![Symmetry 15 01164 g002]()
Table 1.
Left and right sides of the cortical volume, cortical surface area, fronto-occipital (FO) length of the cerebral hemispheres, global-gyrification index (global-GI), and asymmetry quotient (AQ) values of these four measurements in adult cynomolgus monkeys.
Table 1.
Left and right sides of the cortical volume, cortical surface area, fronto-occipital (FO) length of the cerebral hemispheres, global-gyrification index (global-GI), and asymmetry quotient (AQ) values of these four measurements in adult cynomolgus monkeys.
| | Male (n = 5) | Female (n = 5) |
|---|
| | Left | Right | AQ Values | Left | Right | AQ Values |
|---|
| Volume (mm3) | 12,847 ± 1347 | 12,740 ± 1178 | −0.007 ± 0.014 | 11,626 ± 902 | 11,688 ± 878 | 0.005 ± 0.021 |
| Surface area (mm2) | 6661 ± 587 | 6613 ± 542 | −0.007 ± 0.008 | 7287 ± 457 | 7252 ± 381 | −0.004 ± 0.023 |
| FO length (mm) | 60.8 ± 2.2 a | 60.7 ± 2.3 b | −0.002 ± 0.006 | 49.6 ± 1.6 | 49.7 ± 1.2 | 0.002 ± 0.010 |
| Global-GI | 1.683 ± 0.054 | 1.688 ± 0.814 | 0.003 ± 0.008 | 1.684 ± 0.013 | 1.683 ± 0.02 | 0.000 ± 0.005 |
Table 2.
Sulcal surface areas of the left and right sides of the cerebral cortex in adult cynomolgus monkeys.
Table 2.
Sulcal surface areas of the left and right sides of the cerebral cortex in adult cynomolgus monkeys.
| | Male (n = 5) | Female (n = 5) |
|---|
| | Left | Right | Left | Right |
|---|
| Major cerebral sulci | | | | |
| Lateral fissure (lf) | 353.1 ± 56.9 | 352.3 ± 54.0 | 386.2 ± 50.9 | 386.6 ± 50.4 |
| Central sulcus (cs) | 166.9 ± 34.6 | 162.6 ± 26.5 | 168.0 ± 26.2 | 168.3 ± 13.0 |
| Parietooccipital sulcus (pos) | 73.7 ± 22.9 | 64.5 ± 34.0 | 100.2 ± 26.7 | 101.0 ± 13.3 |
| Calcarine sulcus (cal) | 323.7 ± 41.0 | 307.5 ± 31.2 | 319.4 ± 33.4 | 309.4 ± 31.4 |
| Cingulate sulcus (cgs) | 264.0 ± 30.8 | 257.1 ± 28.6 | 301.9 ± 35.7 | 303.6 ± 42.2 |
| Circular sulcus (cirs) | 229.5 ± 38.5 | 239.9 ± 35.9 | 238.1 ± 33.8 | 240.6 ± 35.8 |
| Frontal lobe | | | | |
| Arcuate sulcus, superior ram (sars) | 65.3 ± 16.9 | 65.2 ± 14.3 | 67.8 ± 14.5 | 71.9 ± 12.6 |
| Arcuate sulcus, inferior ram (iars) | 73.8 ± 18.7 | 74.8 ± 17.2 | 86.8 ± 20.6 | 85.4 ± 14.7 |
| Principal sulcus (ps) | 166.1 ± 17.8 | 167.4 ± 31.8 | 188.3 ± 28.2 | 185.5 ± 25.0 |
| Medial orbital sulcus (morb) | 99.2 ± 19.9 | 94.7 ± 20.9 | 132.8 ± 21.6 | 129.8 ± 18.7 |
| Lateral orbital sulcus (lorb) | 42.4 ± 21.7 | 43.1 ± 13.6 | 72.2 ± 10.8 | 63.6 ± 23.4 |
| Olfactory sulcus (olf) | 5.4 ± 0.8 | 6.2 ± 1.3 | 10.1 ± 8.9 | 10.4 ± 9.5 |
| Parietal lobe | | | | |
| Intraparietal sulcus (ips) | 335.6 ± 59.5 | 317.9 ± 51.3 | 335.1 ± 24.5 | 327.6 ± 20.3 |
| Temporal lobe | | | | |
| Superior temporal sulcus (sts) | 472.3 ± 109.2 | 474.7 ± 97.1 | 542.3 ± 36.9 | 540.2 ± 43.3 |
| Occipitotemporal sulcus (ots) | 130.2 ± 73.3 | 134.6 ± 43.0 | 146.2 ± 26.8 | 140.7 ± 15.8 |
| Anterior middle temporal sulcus (amts) | 59.7 ± 23.1 | 55.2 ± 11.0 | 58.8 ± 8.0 | 52.5 ± 13.0 |
| Posterior middle temporal sulcus (pmts) | 25.2 ± 9.7 | 37.4 ± 20.96 | 45.5 ± 12.7 | 38.1 ± 17.2 |
| Rhinal fissure (rf) | 22.3 ± 7.4 | 24.1 ± 7.9 | 16.0 ± 6.9 | 17.2 ± 8.9 |
| Collateral sulcus (cos) | 77.7 ± 26.2 | 71.0 ± 18.5 | 92.1 ± 31.4 | 80.0 ± 14.9 |
| Occipital lobe | | | | |
| Lunate sulcus (lu) | 101.8 ± 19.0 | 102.7 ± 5.2 | 113.7 ± 23.3 | 109.0 ± 15.5 |
| Inferior occipital sulcus (ios) | 175.9 ± 45.8 | 183.0 ± 25.2 | 220.9 ± 28.8 | 225.1 ± 10.1 |
| External calcarine sulcus (ecal) | 33.1 ± 6.7 | 27.6 ± 8.7 | 47.9 ± 13.3 | 50.6 ± 17.0 |
| Superior calcarine sulcus (scal) | 75.6 ± 10.3 | 64.2 ± 15.2 | 91.1 ± 39.3 | 84.8 ± 32.4 |
| Inferior calcarine sulcus (ical) | 75.7 ± 27.2 | 78.7 ± 26.2 | 79.7 ± 32.5 | 84.6 ± 31.5 |
| Simian fossa (sf) | 136.2 ± 18.0 | 121.4 ± 21.9 | 121.4 ± 21.9 | 117.2 ± 27.6 |
| Limbic cortex/Others | | | | |
| Anterior parietooccipital sulcus (apos) | 44.7 ± 21.4 | 44.7 ± 13.6 | 24.5 ± 20.2 | 26.6 ± 17.2 |
| Secondary sulci | 195.5 ± 51.1 | 188.2 ± 67.4 | 139.5 ± 41.3 | 170.5 ± 49.2 |
Table 3.
Asymmetry quotient (AQ) of the sulcal surface areas of the cerebral cortex in adult cynomolgus monkeys.
Table 3.
Asymmetry quotient (AQ) of the sulcal surface areas of the cerebral cortex in adult cynomolgus monkeys.
| | AQ Values |
|---|
| | Male (n = 5) | Female (n = 5) |
|---|
| Major cerebral sulci | | |
| Lateral fissure (lf) | −0.001 ± 0.039 | 0.001 ± 0.050 |
| Central sulcus (cs) | −0.019 ± 0.072 | 0.010 ± 0.099 |
| Parietooccipital sulcus (pos) | −0.202 ± 0.219 | 0.029 ± 0.141 |
| Calcarine sulcus (cal) | −0.049 ± 0.037 | −0.032 ± 0.085 |
| Cingulate sulcus (cgs) | −0.026 ± 0.030 | 0.003 ± 0.041 |
| Circular sulcus (cirs) | 0.047 ± 0.034 | 0.010 ± 0.018 |
| Frontal lobe | | |
| Arcuate sulcus, superior ram (sars) | 0.006 ± 0.156 | 0.065 ± 0.084 |
| Arcuate sulcus, inferior ram (iars) | 0.018 ± 0.029 | −0.005 ± 0.160 |
| Principal sulcus (ps) | −0.002 ± 0.110 | −0.012 ± 0.042 |
| Medial orbital sulcus (morb) | −0.050 ± 0.029 | −0.019 ± 0.085 |
| Lateral orbital sulcus (lorb) | 0.067 ± 0.297 | −0.179 ± 0.467 |
| Olfactory sulcus (olf) | 0.126 ± 0.208 | 0.006 ± 0.120 |
| Parietal lobe | | |
| Intraparietal sulcus (ips) | −0.052 ± 0.045 | −0.022 ± 0.045 |
| Temporal lobe | | |
| Superior temporal sulcus (sts) | 0.011 ± 0.048 | −0.005 ± 0.031 |
| Occipitotemporal sulcus (ots) | 0.095 ± 0.196 | −0.029 ± 0.163 |
| Anterior middle temporal sulcus (amts) | −0.007 ± 0.320 | −0.130 ± 0.296 |
| Posterior middle temporal sulcus (pmts) | 0.295 ± 0.522 | −0.238 ± 0.235 |
| Rhinal fissure (rf) | 0.086 ± 0.109 | 0.041 ± 0.170 |
| Collateral sulcus (cos) | −0.071 ± 0.093 | −0.115 ± 0.235 |
| Occipital lobe | | |
| Lunate sulcus (lu) | 0.020 ± 0.168 | −0.031 ± 0.160 |
| Inferior occipital sulcus (ios) | 0.060 ± 0.164 | 0.024 ± 0.107 |
| External calcarine sulcus (ecal) | −0.196 ± 0.330 | 0.046 ± 0.223 |
| Superior calcarine sulcus (scal) | −0.177 ± 0.232 | −0.061 ± 0.152 |
| Inferior calcarine sulcus (ical) | 0.051 ± 0.113 | 0.087 ± 0.176 |
| Simian fossa (sf) | −0.069 ± 0.091 | −0.048 ± 0.133 |
| Limbic cortex/Others | | |
| Anterior parietooccipital sulcus (apos) | 0.064 ± 0.258 | 0.184 ± 0.643 |
| Secondary sulci | −0.072 ± 0.311 | 0.197 ± 0.132 |
Table 4.
Sulcal-gyrification index (sulcal-GI) of the left and right sides of the cerebral cortex in adult cynomolgus monkeys.
Table 4.
Sulcal-gyrification index (sulcal-GI) of the left and right sides of the cerebral cortex in adult cynomolgus monkeys.
| | Male (n = 5) | Female (n = 5) |
|---|
| | Left | Right | Left | Right |
|---|
| Major cerebral sulci | | | | |
| Lateral fissure (lf) | 1.0577± 0.0084 | 1.0580 ± 0.0086 | 1.0619 ± 0.0074 | 1.0612 ± 0.0064 |
| Central sulcus (cs) | 1.0264 ± 0.0046 | 1.0260 ± 0.0045 | 1.0243 ± 0.0048 | 1.0245 ± 0.0028 |
| Parietooccipital sulcus (pos) | 1.0147 ± 0.0056 | 1.0130 ± 0.0077 | 1.0183 ± 0.0054 | 1.0186 ± 0.0031 |
| Calcarine sulcus (cal) | 1.0707 ± 0.0053 | 1.0688 ± 0.0032 | 1.0654 ± 0.0039 | 1.0635 ± 0.0039 |
| Cingulate sulcus (cgs) | 1.0522 ± 0.0046 | 1.0513 ± 0.0056 | 1.0535 ± 0.0055 | 1.0538 ± 0.0071 |
| Circular sulcus (cirs) | 1.0520 ± 0.0069 | 1.0546 ± 0.0054 | 1.0501 ± 0.0055 | 1.0506 ± 0.0056 a |
| Frontal lobe | | | | |
| Arcuate sulcus, superior ram (sars) | 1.0095 ± 0.0026 | 1.0106 ± 0.0028 | 1.0078 ± 0.0023 | 1.0083 ± 0.0020 |
| Arcuate sulcus, inferior ram (iars) | 1.0102± 0.0017 | 1.0113 ± 0.0017 | 1.0097 ± 0.0017 | 1.0102 ± 0.0013 |
| Principal sulcus (ps) | 1.0429 ± 0.0035 | 1.0449 ± 0.0048 | 1.0420 ± 0.0033 | 1.0421 ± 0.0037 |
| Medial orbital sulcus (morb) | 1.0121 ± 0.0030 | 1.0117 ± 0.0032 | 1.0119 ± 0.0020 | 1.0125 ± 0.0019 |
| Lateral orbital sulcus (lorb) | 1.0022 ± 0.0023 | 1.0018 ± 0.0007 | 1.0040 ± 0.0014 | 1.0042 ± 0.0038 |
| Olfactory sulcus (olf) | 1.0005 ± 0.0001 | 1.0006 ± 0.0002 | 1.0005 ± 0.0004 | 1.0006 ± 0.0005 |
| Parietal lobe | | | | |
| Intraparietal sulcus (ips) | 1.0583 ± 0.0121 | 1.0580 ± 0.0109 | 1.0534 ± 0.0039 | 1.0528 ± 0.0030 |
| Temporal lobe | | | | |
| Superior temporal sulcus (sts) | 1.0831 ± 0.0202 | 1.0837 ± 0.0180 | 1.0904 ± 0.0089 | 1.0897 ± 0.0089 |
| Occipitotemporal sulcus (ots) | 1.0182 ± 0.0156 | 1.0171 ± 0.0095 | 1.0173 ± 0.0035 | 1.0171 ± 0.0015 |
| Anterior middle temporal sulcus (amts) | 1.0052 ± 0.0022 | 1.0047 ± 0.0011 | 1.0048 ± 0.0012 | 1.0049 ± 0.0014 |
| Posterior middle temporal sulcus (pmts) | 1.0019± 0.0010 | 1.0034 ± 0.0025 | 1.0038 ± 0.0017 | 1.0031 ± 0.0027 |
| Rhinal fissure (rf) | 1.0022 ± 0.0007 | 1.0026 ± 0.0009 | 1.0010 ± 0.0006 | 1.0013 ± 0.0008 |
| Collateral sulcus (cos) | 1.0147 ± 0.0041 | 1.0142 ± 0.0031 | 1.0151 ± 0.0062 | 1.0133 ± 0.0025 |
| Occipital lobe | | | | |
| Lunate sulcus (lu) | 1.0204 ± 0.0022 | 1.0217 ± 0.0007 | 1.0209 ± 0.0055 | 1.0204 ± 0.0033 |
| Inferior occipital sulcus (ios) | 1.0345 ± 0.0087 | 1.0371 ± 0.0049 | 1.0423 ± 0.0068 | 1.0435 ± 0.0058 |
| External calcarine sulcus (ecal) | 1.0007 ± 0.0005 | 1.0008 ± 0.0007 | 1.0019 ± 0.0020 | 1.0039 ± 0.0055 |
| Superior calcarine sulcus (scal) | 1.0213 ± 0.0062 | 1.0174 ± 0.0067 | 1.0223 ± 0.0092 | 1.0211 ± 0.0087 |
| Inferior calcarine sulcus (ical) | 1.0206 ± 0.0052 | 1.0214 ± 0.0051 | 1.0206 ± 0.0089 | 1.0221 ± 0.0074 |
| Simian fossa (sf) | 1.0327± 0.0023 | 1.0266 ± 0.0040 | 1.0266 ± 0.0040 | 1.0256 ± 0.0053 |
| Limbic cortex/Others | | | | |
| Anterior parietooccipital sulcus (apos) | 1.0064 ± 0.0033 | 1.0069 ± 0.0024 | 1.0033 ± 0.0032 | 1.0037 ± 0.0029 |
| Secondary sulci | 1.0110 ± 0.0017 | 1.0127 ± 0.0017 | 1.0092 ± 0.0030 | 1.0084 ± 0.0026 |
Table 5.
Asymmetry quotient (AQ) of the sulcal-gyrification index (sulcal-GI) of the cerebral cortex in adult cynomolgus monkeys.
Table 5.
Asymmetry quotient (AQ) of the sulcal-gyrification index (sulcal-GI) of the cerebral cortex in adult cynomolgus monkeys.
| | AQ Values |
|---|
| | Male (n = 5) | Female (n = 5) |
|---|
| Major cerebral sulci | | |
| Lateral fissure (lf) | 0.000 ± 0.002 | −0.001 ± 0.003 |
| Central sulcus (cs) | 0.000 ± 0.002 | 0.000 ± 0.002 |
| Parietooccipital sulcus (pos) | −0.002 ± 0.002 | 0.000 ± 0.003 |
| Calcarine sulcus (cal) | −0.002 ± 0.003 | −0.002 ± 0.002 |
| Cingulate sulcus (cgs) | −0.001 ± 0.002 | 0.000 ± 0.002 |
| Circular sulcus (cirs) | 0.003 ± 0.002 | 0.001 ± 0.002 |
| Frontal lobe | | |
| Arcuate sulcus, superior ram (sars) | 0.001 ± 0.002 | 0.001 ± 0.001 |
| Arcuate sulcus, inferior ram (iars) | 0.001 ± 0.000 | 0.000 ± 0.001 |
| Principal sulcus (ps) | 0.002 ± 0.003 | 0.000 ± 0.003 |
| Medial orbital sulcus (morb) | 0.000 ± 0.001 | 0.001 ± 0.001 |
| Lateral orbital sulcus (lorb) | 0.000 ± 0.002 | 0.000 ± 0.003 |
| Olfactory sulcus (olf) | 0.000 ± 0.000 | 0.000 ± 0.002 |
| Parietal lobe | | |
| Intraparietal sulcus (ips) | 0.000 ± 0.002 | −0.001 ± 0.003 |
| Temporal lobe | | |
| Superior temporal sulcus (sts) | 0.001 ± 0.004 | −0.001 ± 0.003 |
| Occipitotemporal sulcus (ots) | −0.001 ± 0.006 | 0.000 ± 0.003 |
| Anterior middle temporal sulcus (amts) | −0.001 ± 0.001 | 0.000 ± 0.001 |
| Posterior middle temporal sulcus (pmts) | 0.001 ± 0.002 | −0.001 ± 0.001 |
| Rhinal fissure (rf) | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Collateral sulcus (cos) | 0.000 ± 0.001 | −0.002 ± 0.005 |
| Occipital lobe | | |
| Lunate sulcus (lu) | 0.001 ± 0.002 | 0.000 ± 0.004 |
| Inferior occipital sulcus (ios) | 0.003 ± 0.004 | 0.001 ± 0.004 |
| External calcarine sulcus (ecal) | 0.000 ± 0.000 | 0.002 ± 0.004 |
| Superior calcarine sulcus (scal) | −0.004 ± 0.004 | −0.001 ± 0.003 |
| Inferior calcarine sulcus (ical) | 0.001 ± 0.002 | 0.001 ± 0.003 |
| Simian fossa (sf) | −0.001 ± 0.003 | −0.001 ± 0.003 |
| Limbic cortex/Others | | |
| Anterior parietooccipital sulcus (apos) | 0.001 ± 0.001 | 0.000 ± 0.001 |
| Secondary sulci | 0.002 ± 0.003 | −0.001 ± 0.003 |
Table 6.
Incidence of secondary sulci on the left and right sides of the cerebral cortex in adult cynomolgus monkeys.
Table 6.
Incidence of secondary sulci on the left and right sides of the cerebral cortex in adult cynomolgus monkeys.
| | Male (n = 5) | Female (n = 5) |
|---|
| | Left | Right | Asymmetrical | Left | Right | Asymmetrical |
|---|
| Frontal lobe | | | | | | |
| Supr of arcuate sulcus (sas) | 100% (5/5) | 100% (5/5) | 0% (0/5) | 100% (5/5) | 100% (5/5) | 0% (0/5) |
| Anterior supraprincipal dimple (aspd) | 80% (4/5) | 100% (5/5) | 20% (1/5) | 0% (0/5) | 40% (2/5) | 0% (0/5) |
| Posterior supraprincipal dimple(pspd) | 40% (2/5) | 60% (3/5) | 60% (3/5) | 80% (4/5) | 60% (3/5) | 60% (3/5) |
| Superior precentral dimple (spcd) | 100% (5/5) | 100% (5/5) | 0% (0/5) | 100% (5/5) | 80% (4/5) | 20% (1/5) |
| Infraprincipal dimple (ipd) | 80% (4/5) | 60% (3/5) | 40% (2/5) | 80% (4/5) | 40% (2/5) | 0% (0/5) |
| Anterior subcentral dimple (asd) | 100% (5/5) | 100% (5/5) | 0% (0/5) | 100% (5/5) | 80% (4/5) | 20% (1/5) |
| Intermediate orbital sulcus (iorb) | 80% (4/5) | 80% (4/5) | 40% (2/5) | 40% (2/5) | 80% (4/5) | 40% (2/5) |
| Rostral sulcus (ros) | 80% (4/5) | 80% (4/5) | 0% (0/5) a | 60% (3/5) | 40% (2/5) | 60% (3/5) a |
| Parietal lobe | | | | | | |
| Superior postcentral dimple (su) | 80% (4/5) | 80% (4/5) | 0% (0/5) a | 20% (1/5) | 80% (4/5) | 60% (3/5) a |
| Posterior subcentral sulcus (pscs) | 40% (2/5) | 80% (4/5) | 40% (2/5) | 40% (2/5) | 20% (1/5) | 20% (1/5) |
| Temporal Lobe | | | | | | |
| Intermediate middle temporal sulcus (imt) | 80% (4/5) | 60% (3/5) | 20% (1/5) | 80% (4/5) | 80% (4/5) | 40% (2/5) |
| Limbic cortex | | | | | | |
| Subparietal sulcus (sbps) | 60% (3/5) | 60% (3/5) | 0% (0/5) | 40% (2/5) | 60% (3/5) | 20% (1/5) |





{kind=link}
{kind=link}