


Influence of Spatial Dispersal among Species in a Prey–Predator Model with Miniature Predator Groups
 , , , and
, , , and
Abstract
:1. Introduction
2. The Mathematical Model
3. Linear Stability Analysis
- ,
- , and
- .
- ,
- .
- 1.
- ,
- 2.
- .
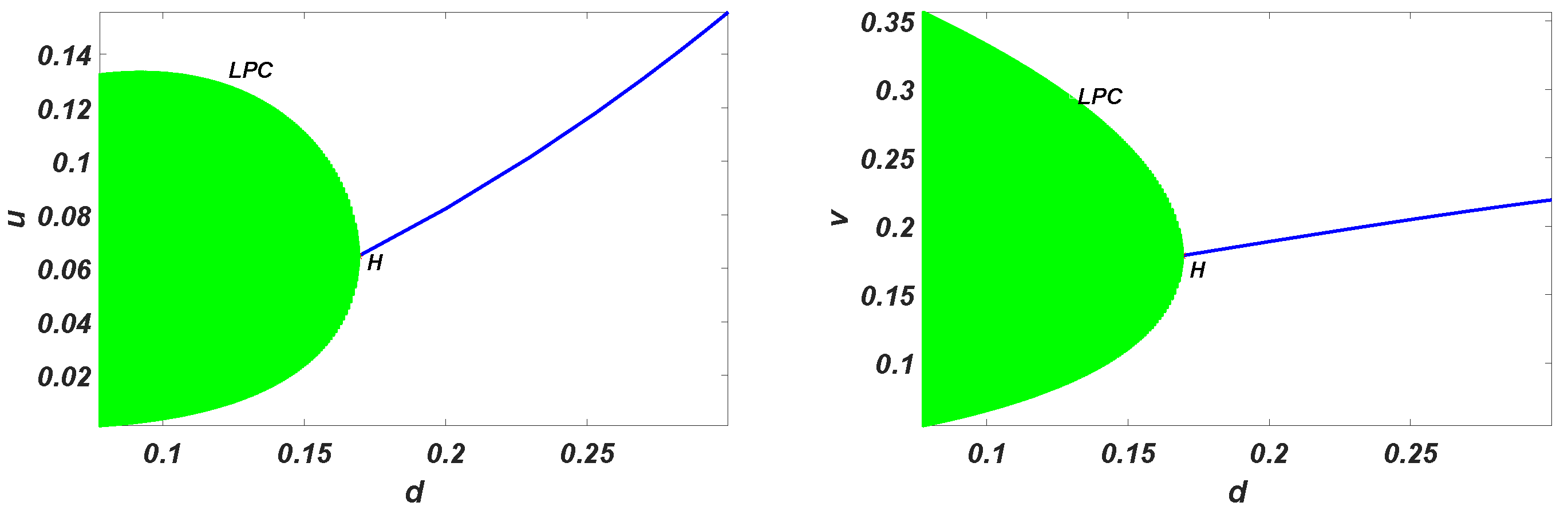
Hopf Bifurcation Analysis
- 1.
- 2.
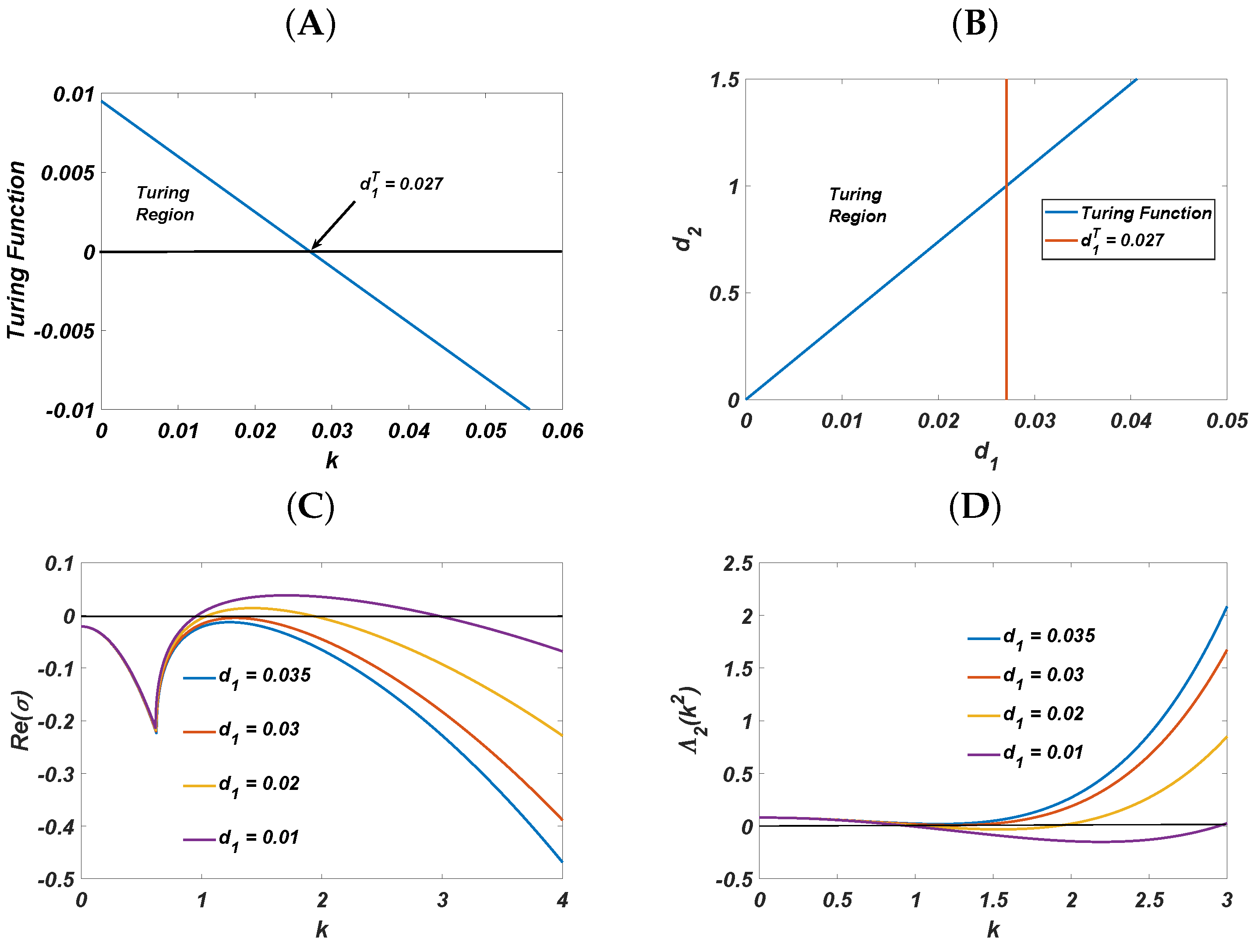
4. Diffusion-Driven Instability
- 1.
- and
- 2.
- ,
- 3.
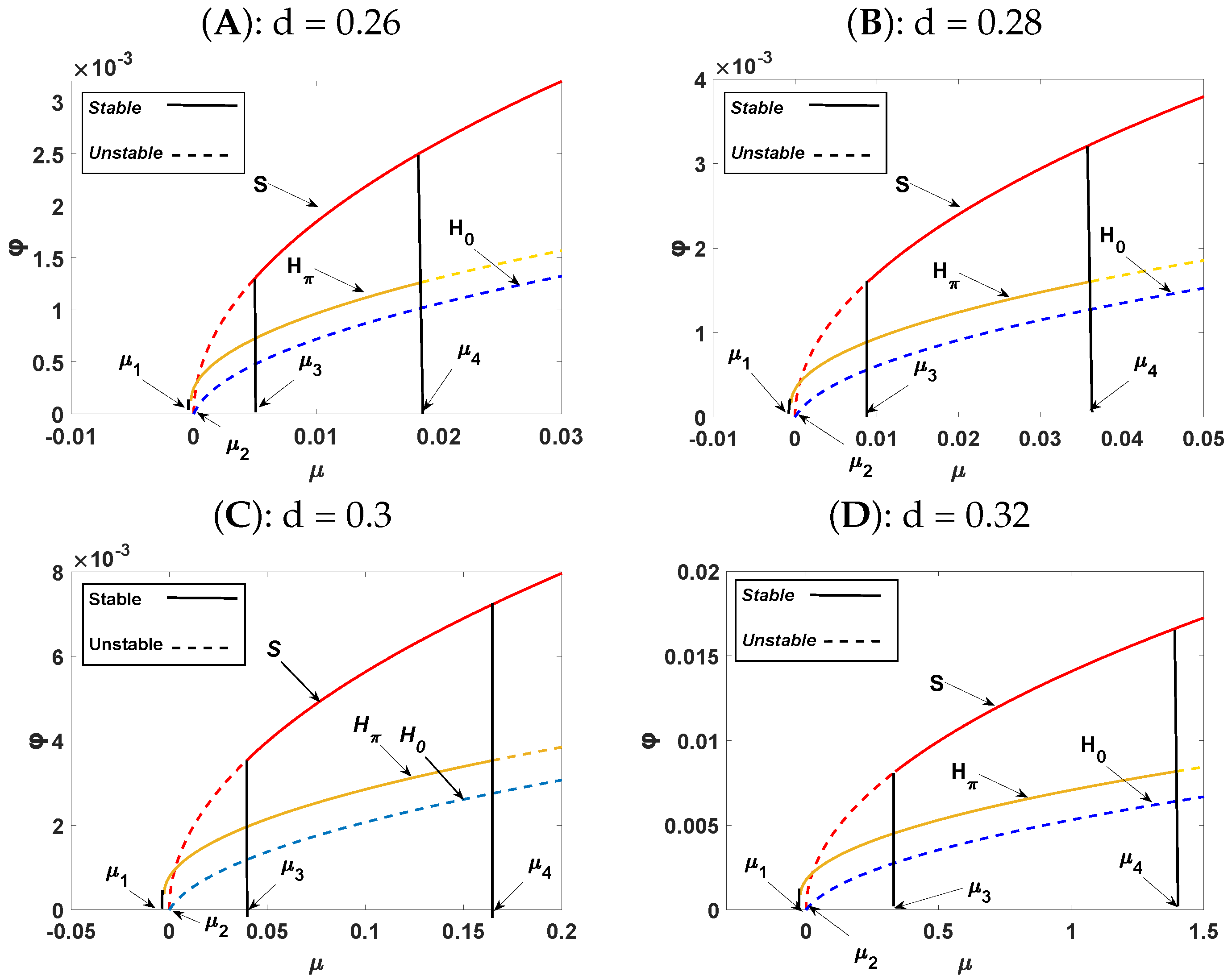
5. Weakly Nonlinear Analysis
- The first solution: The stationary state is represented by , which is stable for and unstable for .
- The second solution: The stripe pattern given by is stable for and unstable for .
- The third solution: When , two solutions exist: Hexagon patterns are given by with or , and exist when ; the solution is stable for , and is unstable.
- The mixed states: When with . This exists when and is always unstable.
- 1.
- Homogeneous solution: It is stable for and unstable for .
- 2.
- Stripe solution: It is stable for and unstable for .
- 3.
- Hexagonal solution: It exists when and is stable for and is always unstable.
- 4.
- Mixed solution: It exists but is always unstable when and .
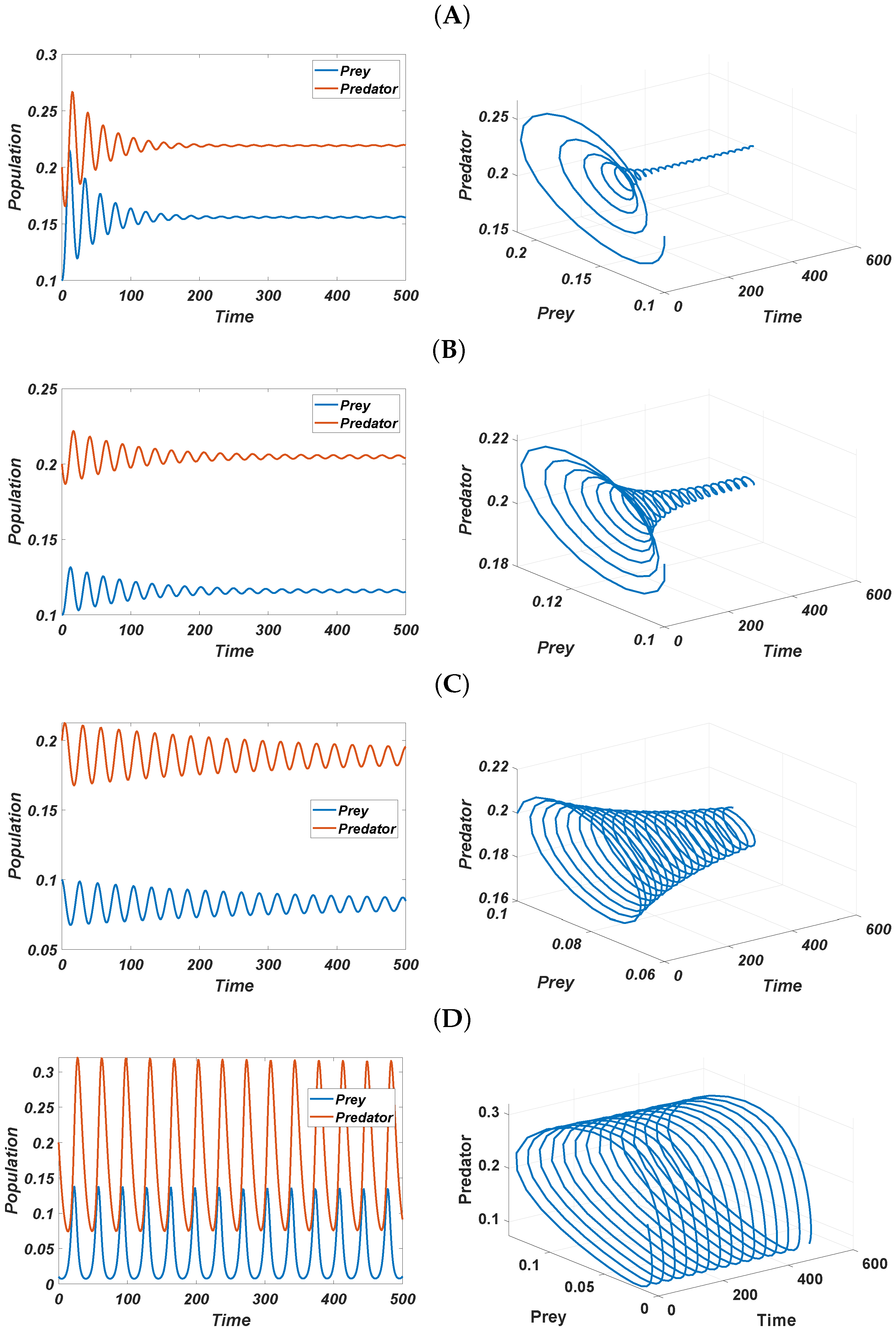
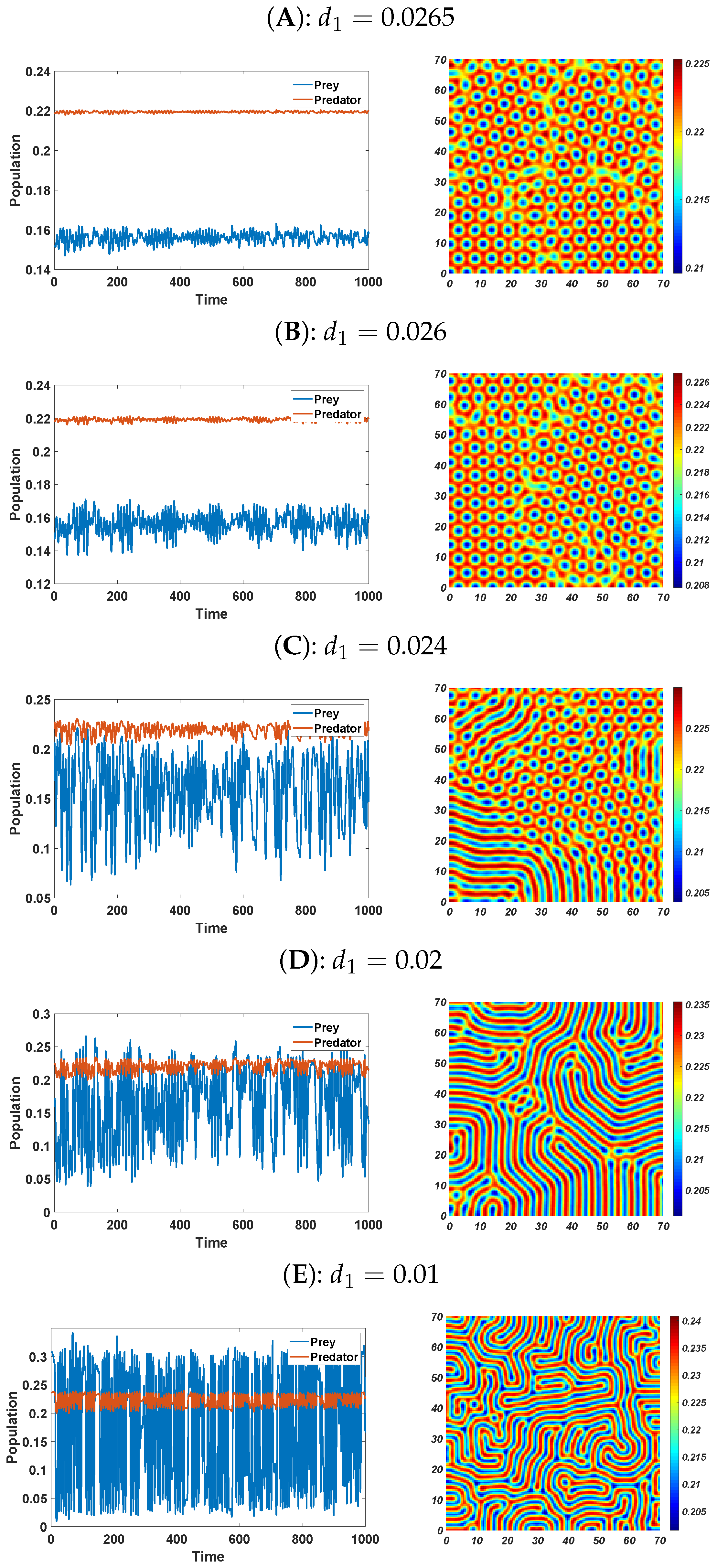
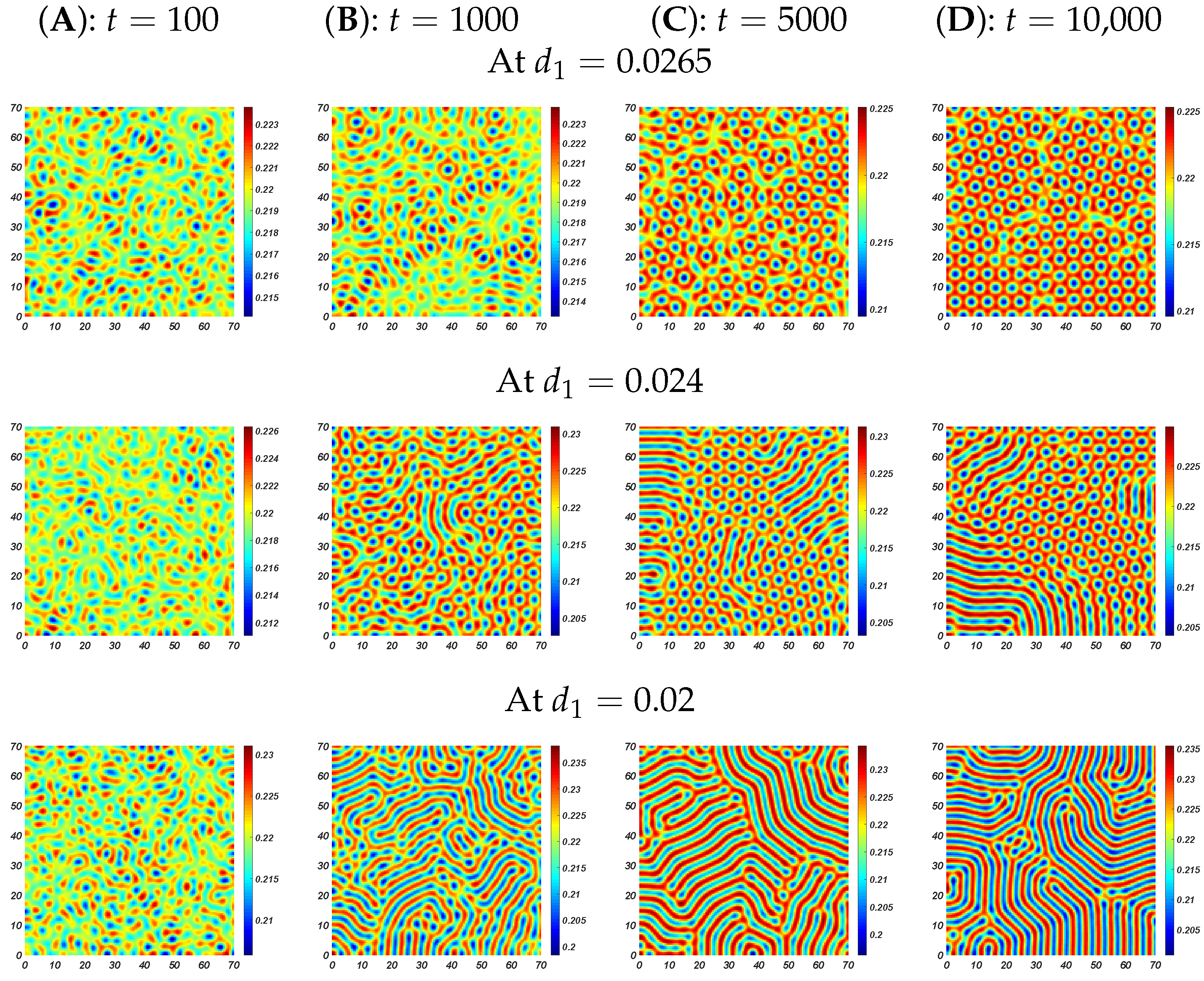
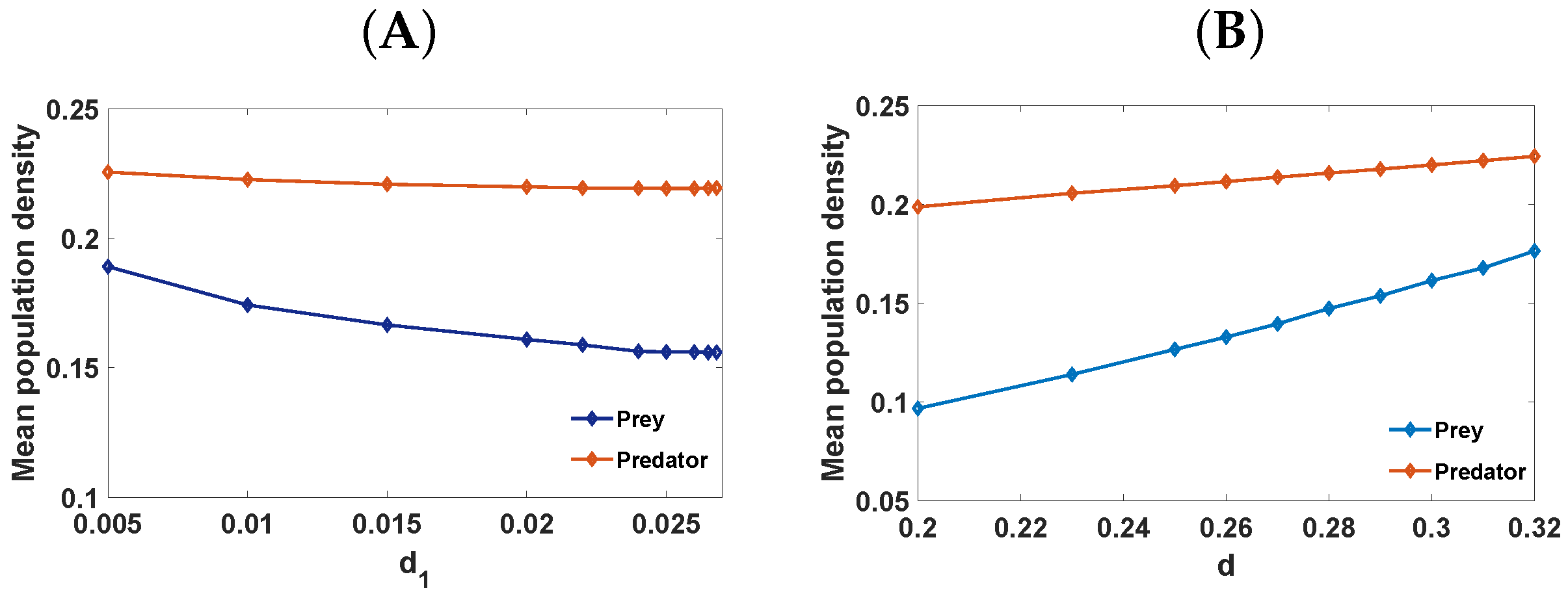
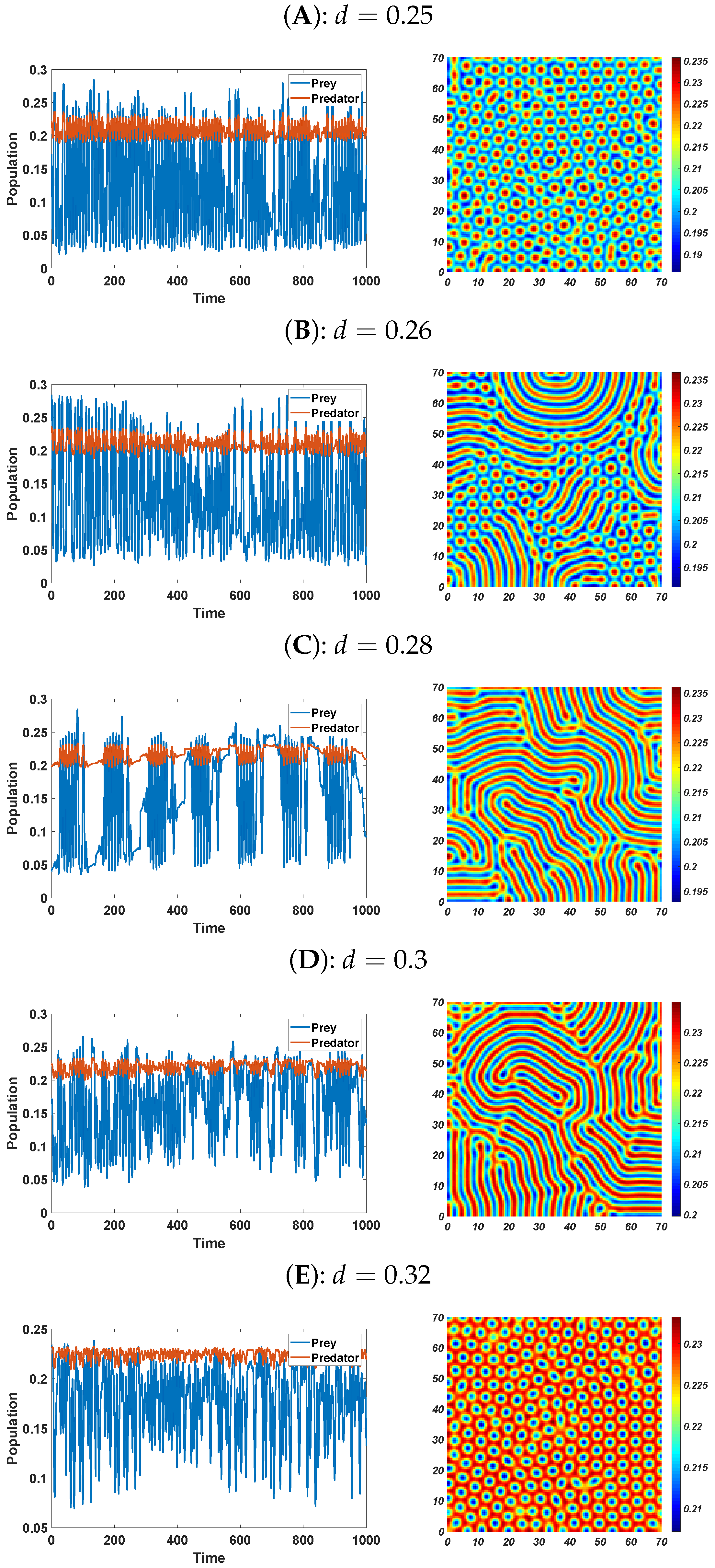
6. Numerical Simulations
6.1. Nonspatial Analysis
6.2. Pattern Selection
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cresswell, W.; Quinn, J.L. Faced with a choice, sparrowhawks more often attack the more vulnerable prey group. Oikos 2004, 104, 71–76. [Google Scholar] [CrossRef]
- Krause, J.; Godin, J.G.J. Predator preferences for attacking particular prey group sizes: Consequences for predator hunting success and prey predation risk. Anim. Behav. 1995, 50, 465–473. [Google Scholar] [CrossRef]
- Fitzgibbon, C.D. Mixed-species grouping in Thomson’s and Grant’s gazelles: The antipredator benefits. Anim. Behav. 1990, 39, 1116–1126. [Google Scholar] [CrossRef]
- Bailey, I.; Myatt, J.P.; Wilson, A.M. Group hunting within the Carnivora: Physiological, cognitive and environmental influences on strategy and cooperation. Behav. Ecol. Sociobiol. 2013, 67, 1–17. [Google Scholar] [CrossRef]
- Scheel, D. Profitability, encounter rates, and prey choice of African lions. Behav. Ecol. 1993, 4, 90–97. [Google Scholar] [CrossRef]
- Seo, G.; DeAngelis, D.L. A predator-prey model with a Holling type I functional response including a predator mutual interference. J. Nonlinear Sci. 2011, 21, 811–833. [Google Scholar] [CrossRef]
- Singh, T.; Dubey, R.; Mishra, V.N. Spatial dynamics of a predator-prey system with hunting cooperation in predators and type I functional response. AIMS Math. 2020, 5, 673–684. [Google Scholar] [CrossRef]
- Singh, T.; Banerjee, S. Spatial aspect of hunting cooperation in predators with Holling type II functional response. J. Biol. Syst. 2018, 26, 511–531. [Google Scholar] [CrossRef]
- Fu, S.; Zhang, H. Effect of hunting cooperation on the dynamic behavior for a diffusive Holling type II predator-prey model. Commun. Nonlinear Sci. Numer. Simul. 2021, 99, 105807. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, F.; Zhong, L. Stability analysis of a prey-predator model with Holling type III response function incorporating a prey refuge. Appl. Math. Comput. 2006, 182, 672–683. [Google Scholar] [CrossRef]
- Agarwal, M.; Pathak, R. Harvesting and Hopf Bifurcation in a prey-predator model with Holling Type IV Functional Response. Int. J. Math. Soft Comput. 2017, 2, 99. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Naji, R.K. Dynamics of a three species food chain model with Crowley-Martin type functional response. Chaos Solit. Fractals 2009, 42, 1337–1346. [Google Scholar] [CrossRef]
- Zhang, X.C.; Sun, G.Q.; Jin, Z. Spatial dynamics in a predator-prey model with Beddington-DeAngelis functional response. Phys. Rev. E 2012, 85, 021924. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.B.; Hwang, T.W.; Kuang, Y. Global dynamics of a predator-prey model with Hassell-Varley type functional response. Discret. Contin. Dyn. Syst. Ser. B 2008, 10, 857. [Google Scholar] [CrossRef]
- Murray, J. A pre-pattern formation mechanism for animal coat markings. J. Theor. Biol. 1981, 88, 161–199. [Google Scholar] [CrossRef]
- Turing, A.M. The chemical basis of morphogenesis. Bull. Math. Biol. 1990, 52, 153–197. [Google Scholar] [CrossRef]
- Medvinsky, A.B.; Petrovskii, S.V.; Tikhonova, I.A.; Malchow, H.; Li, B.L. Spatiotemporal complexity of plankton and fish dynamics. SIAM Rev. 2002, 44, 311–370. [Google Scholar] [CrossRef]
- Segel, L.A.; Jackson, J.L. Dissipative structure: An explanation and an ecological example. J. Theor. Biol. 1972, 37, 545–559. [Google Scholar] [CrossRef]
- Agiza, H.N.; Elabbasy, E.M.; ElMetwally, H.; Elsadany, A.A. Chaotic dynamics of a discrete prey-predator model with Holling type II. Nonlinear Anal. Real World Appl. 2009, 10, 116–129. [Google Scholar] [CrossRef]
- Capone, F.; Carfora, M.F.; De Luca, R.; Torcicollo, I. Turing patterns in a reaction diffusion system modeling hunting cooperation. Math. Comput. Simul. 2019, 165, 172–180. [Google Scholar] [CrossRef]
- Hao, W.; Xue, C. Spatial pattern formation in reaction-diffusion models: A computational approach. J. Math. Biol. 2020, 80, 521–543. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Pal, N.; Samanta, S.; Chattopadhyay, J. Effect of hunting cooperation and fear in a predator-prey model. Ecol. Complex. 2019, 39, 100770. [Google Scholar] [CrossRef]
- Singh, T.; Dubey, R.; Mishra, V.N.; Abdel-Aty, M. Modeling of Diffusive Patterns in Predator-Prey System using Turing Instability and Amplitude Equations. Inf. Sci. Lett. 2021, 10, 5–16. [Google Scholar]
- Song, Q.; Yang, R.; Zhang, C.; Tang, L. Bifurcation analysis of a diffusive predator-prey model with Monod-Haldane functional response. Int. J. Bifurc. Chaos 2019, 29, 1950152. [Google Scholar] [CrossRef]
- Wang, J.S.; Wu, Y.P.; Li, L.; Sun, G.Q. Effect of mobility and predator switching on the dynamical behavior of a predator-prey model. Chaos Solit. Fractals 2020, 132, 109584. [Google Scholar] [CrossRef]
- Yuan, S.; Xu, C.; Zhang, T. Spatial dynamics in a predator-prey model with herd behavior. Chaos 2013, 23, 033102. [Google Scholar] [CrossRef]
- Zhang, T.; Xing, Y.; Zang, H.; Han, M. Spatio-temporal dynamics of a reaction-diffusion system for a predator-prey model with hyperbolic mortality. Nonlinear Dyn. 2014, 78, 265–277. [Google Scholar] [CrossRef]
- Kumar, V.; Kumari, N. Stability and bifurcation analysis of Hassell-Varley prey-predator system with fear effect. Int. J. Appl. Comput. Math. 2020, 6, 150. [Google Scholar] [CrossRef]
- Li, H.; Tian, Y. Dynamic behavior analysis of a feedback control predator-prey model with exponential fear effect and Hassell-Varley functional response. J. Frankl. Inst. 2023, 360, 3479–3498. [Google Scholar] [CrossRef]
- Du, B.; Hu, M.; Lian, X. Dynamical behavior for a stochastic predator-prey model with HV type functional response. Bull. Malays. Math. Sci. Soc. 2017, 40, 487–503. [Google Scholar] [CrossRef]
- Xie, X.; Zhang, C.; Chen, X.; Chen, J. Almost periodic sequence solution of a discrete Hassell-Varley predator-prey system with feedback control. Appl. Math. Comput. 2015, 268, 35–51. [Google Scholar] [CrossRef]
- Kim, H.K.; Baek, H. The dynamical complexity of a predator-prey system with Hassell-Varley functional response and impulsive effect. Math. Comput. Simul. 2013, 94, 1–14. [Google Scholar] [CrossRef]
- Pathak, S.; Maiti, A.; Samanta, G.P. Rich dynamics of a food chain model with Hassell-Varley type functional responses. Appl. Math. Comput. 2009, 208, 303–317. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, S.; Fan, K.; Wang, Q. Asymptotic behavior of a non-autonomous predator-prey model with Hassell-Varley type functional response and random perturbation. J. Appl. Math. Comput. 2015, 49, 573–594. [Google Scholar] [CrossRef]
- Luo, D.; Wang, D. Impact of discontinuous harvesting policies on prey-predator system with Hassell-Varley type functional response. Int. J. Biomath. 2017, 10, 1750048. [Google Scholar] [CrossRef]
- Chen, D.; Wang, D. Almost periodic dynamics of delayed prey-predator model with discontinuous harvesting policies and Hassell-Varley type functional response. Int. J. Biomath. 2018, 11, 1850083. [Google Scholar] [CrossRef]
- Wang, K.; Zhu, Y.L. Permanence and global asymptotic stability of a delayed predator-prey model with Hassell-Varley type functional response. Bull. Iran. Math. Soc. 2011, 37, 197–215. [Google Scholar]
- Xu, C.; Li, P. Oscillations for a delayed predator-prey model with Hassell-Varley-type functional response. C. R. Biol. 2015, 338, 227–240. [Google Scholar] [CrossRef]
- Skubachevskii, A.L. On necessary conditions for the Fredholm solvability of nonlocal elliptic problems. Proc. Steklov Inst. Math. 2008, 260, 238–253. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Biological Meaning |
|---|---|
| r | is an intrinsic growth rate of prey |
| K | is carrying capacity |
| c | is attack rate |
| is Hassell–Varley (H-V) constant | |
| m | is half-saturation constant |
| f | is conversion rate |
| D | is death rate of predators |
| S.No. | d | |||||||
|---|---|---|---|---|---|---|---|---|
| 1. | 0.26 | 0.035 | 2930.65 | 5745.24 | −0.000219 | 0 | 0.00468 | 0.0185 |
| 2. | 0.28 | 0.031 | 3472.19 | 7091.72 | −0.000478 | 0 | 0.00895 | 0.0362 |
| 3. | 0.3 | 0.027 | 3151.04 | 6871.92 | −0.002559 | 0 | 0.03935 | 0.1645 |
| 4. | 0.32 | 0.022 | 5040.15 | 10,773.10 | −0.020674 | 0 | 0.33716 | 1.3950 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shivam; Aljrees, T.; Singh, T.; Varshney, N.; Kumar, M.; Singh, K.U.; Vimal, V. Influence of Spatial Dispersal among Species in a Prey–Predator Model with Miniature Predator Groups. Symmetry 2023, 15, 986. https://doi.org/10.3390/sym15050986
Shivam, Aljrees T, Singh T, Varshney N, Kumar M, Singh KU, Vimal V. Influence of Spatial Dispersal among Species in a Prey–Predator Model with Miniature Predator Groups. Symmetry. 2023; 15(5):986. https://doi.org/10.3390/sym15050986
Chicago/Turabian StyleShivam, Turki Aljrees, Teekam Singh, Neeraj Varshney, Mukesh Kumar, Kamred Udham Singh, and Vrince Vimal. 2023. "Influence of Spatial Dispersal among Species in a Prey–Predator Model with Miniature Predator Groups" Symmetry 15, no. 5: 986. https://doi.org/10.3390/sym15050986