

Quantifying Shape Variation in an Antisymmetrical Trait in the Tropical Fish Xenophallus umbratilis
Abstract
:1. Introduction
2. Materials and Methods
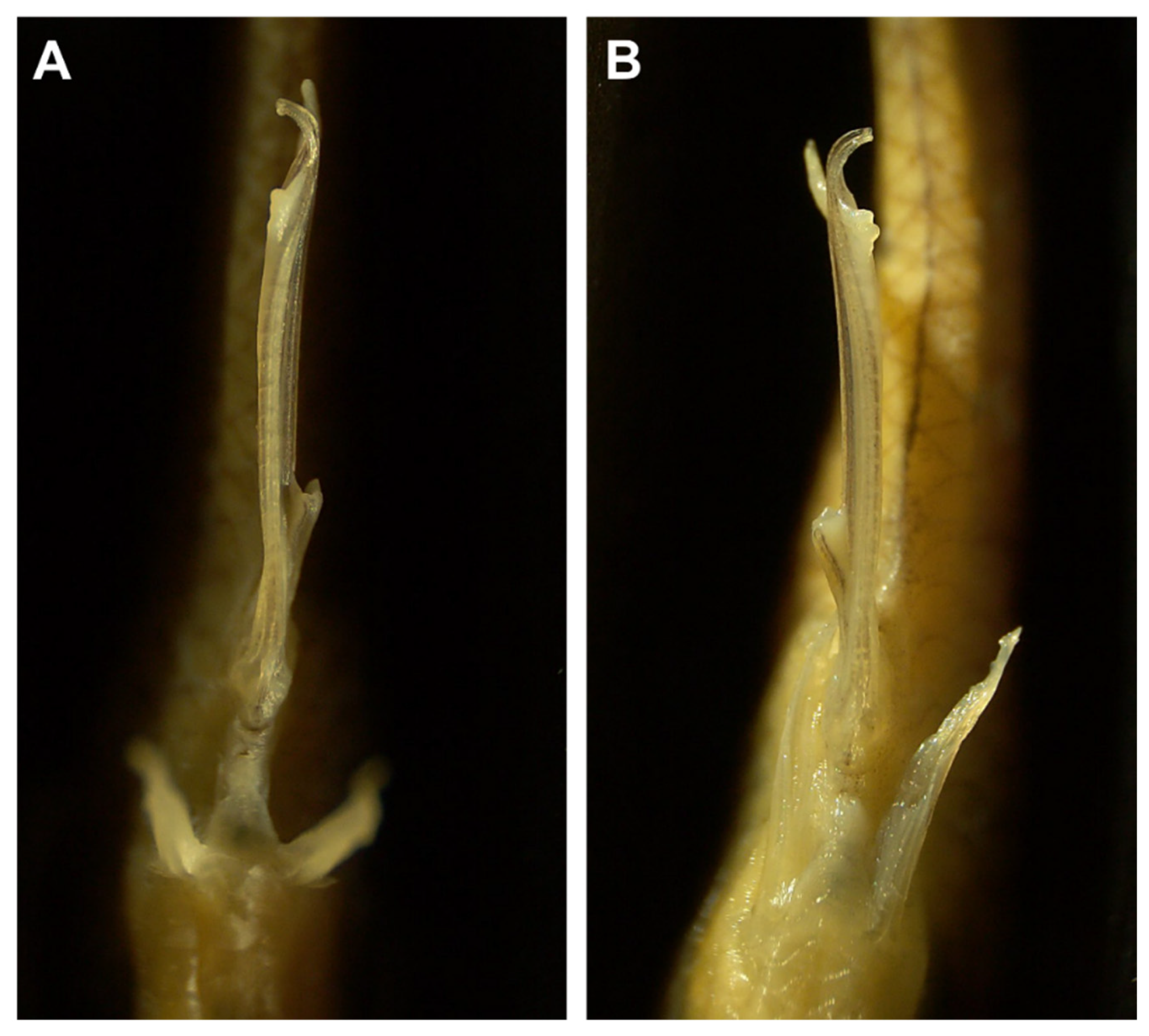
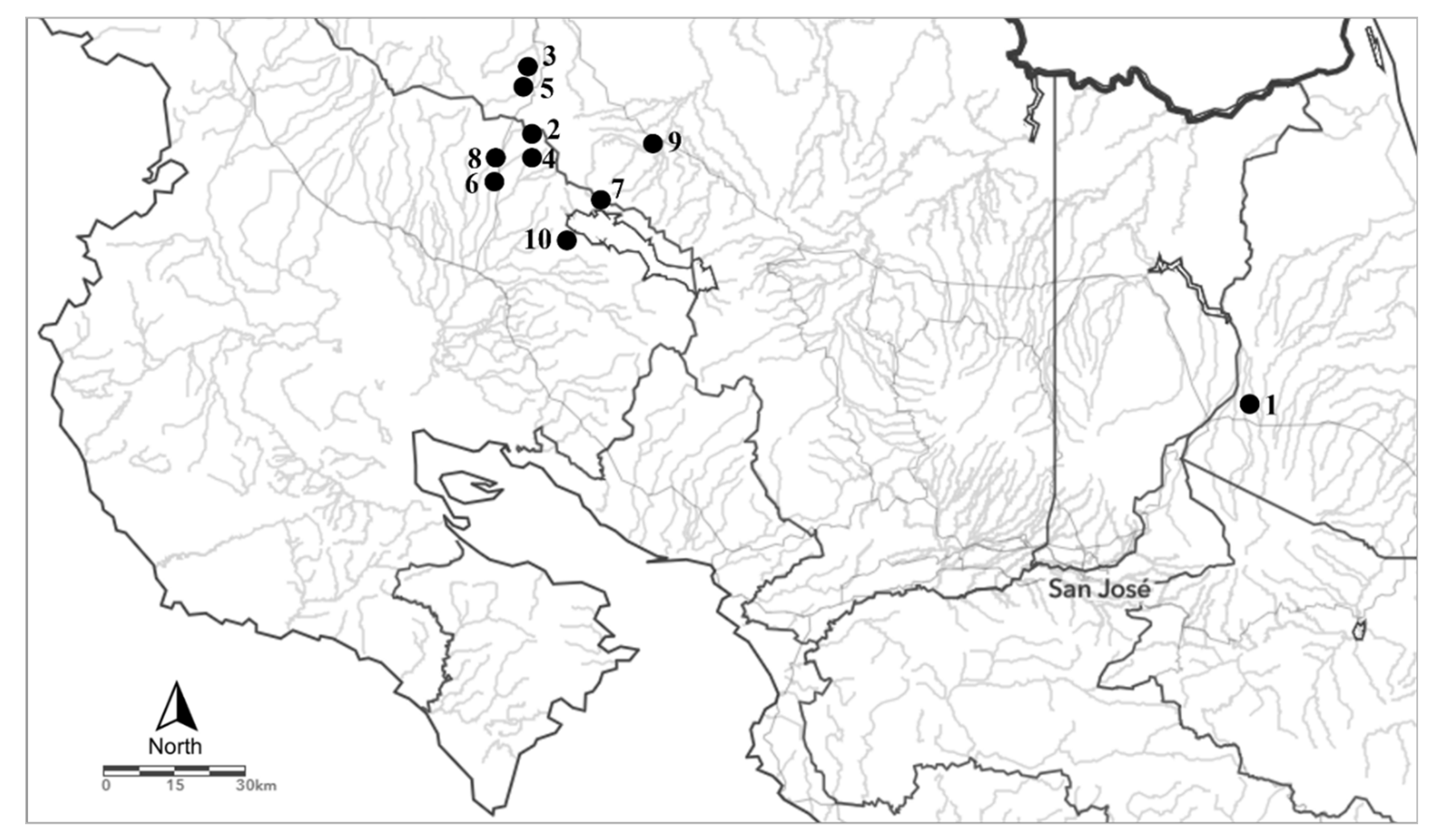
2.1. Study System and Sampling
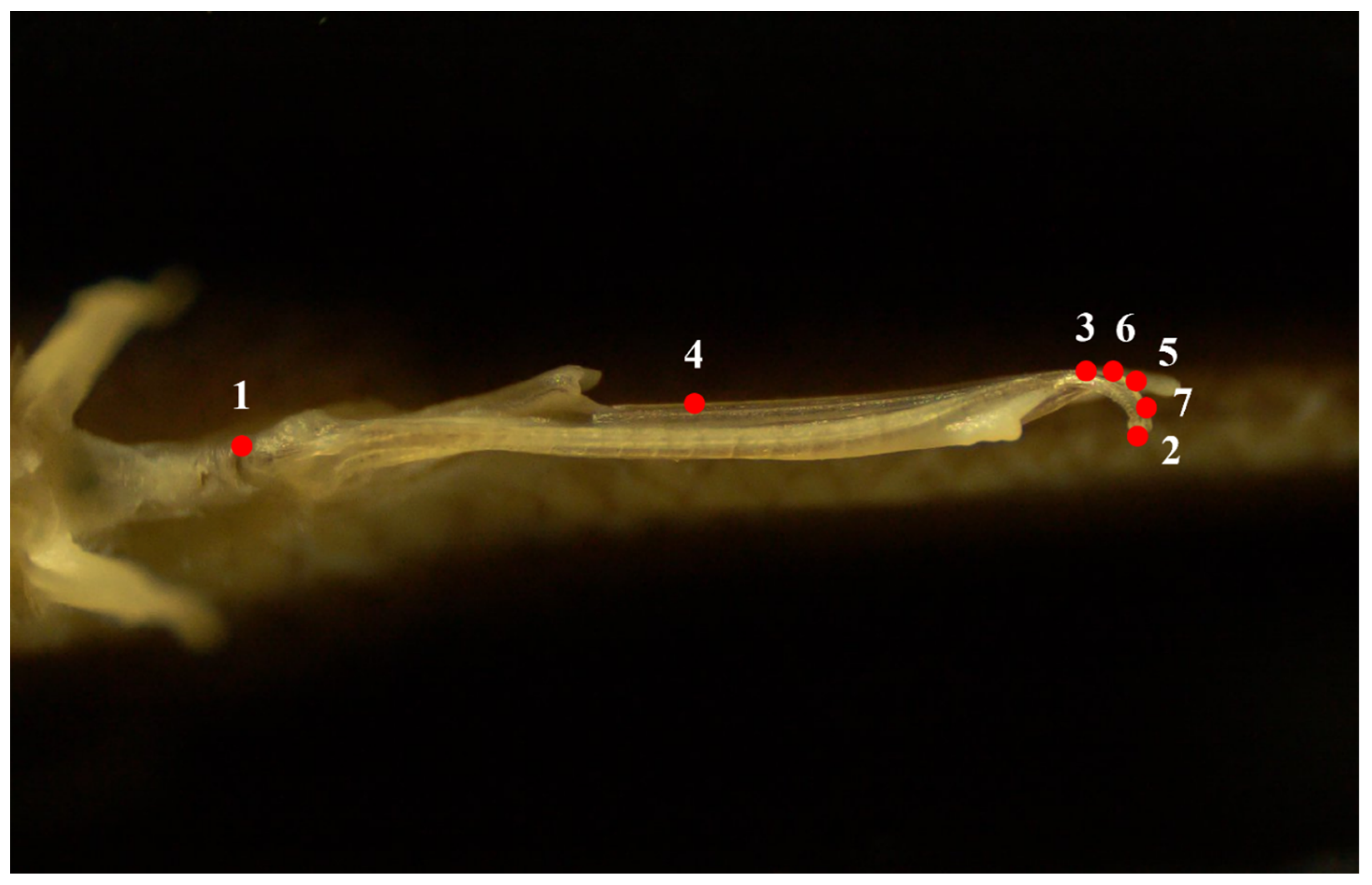
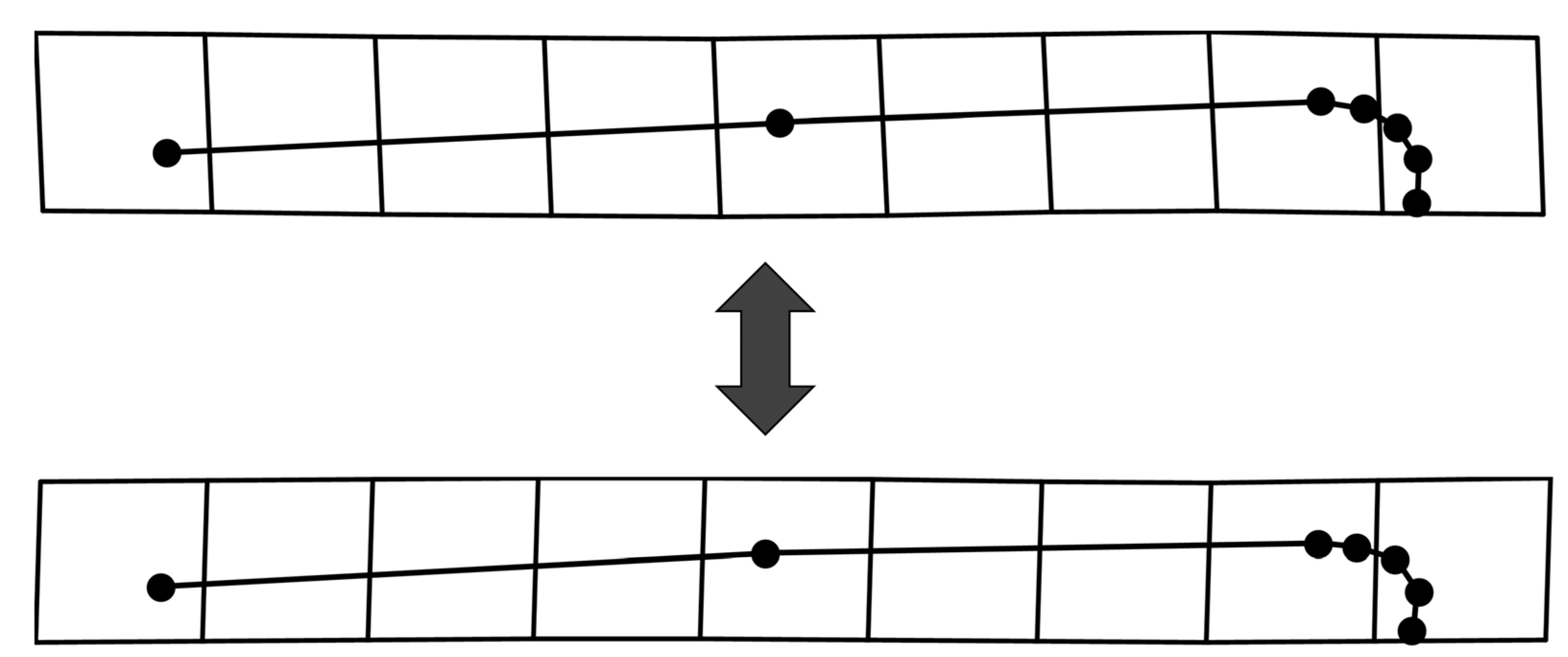
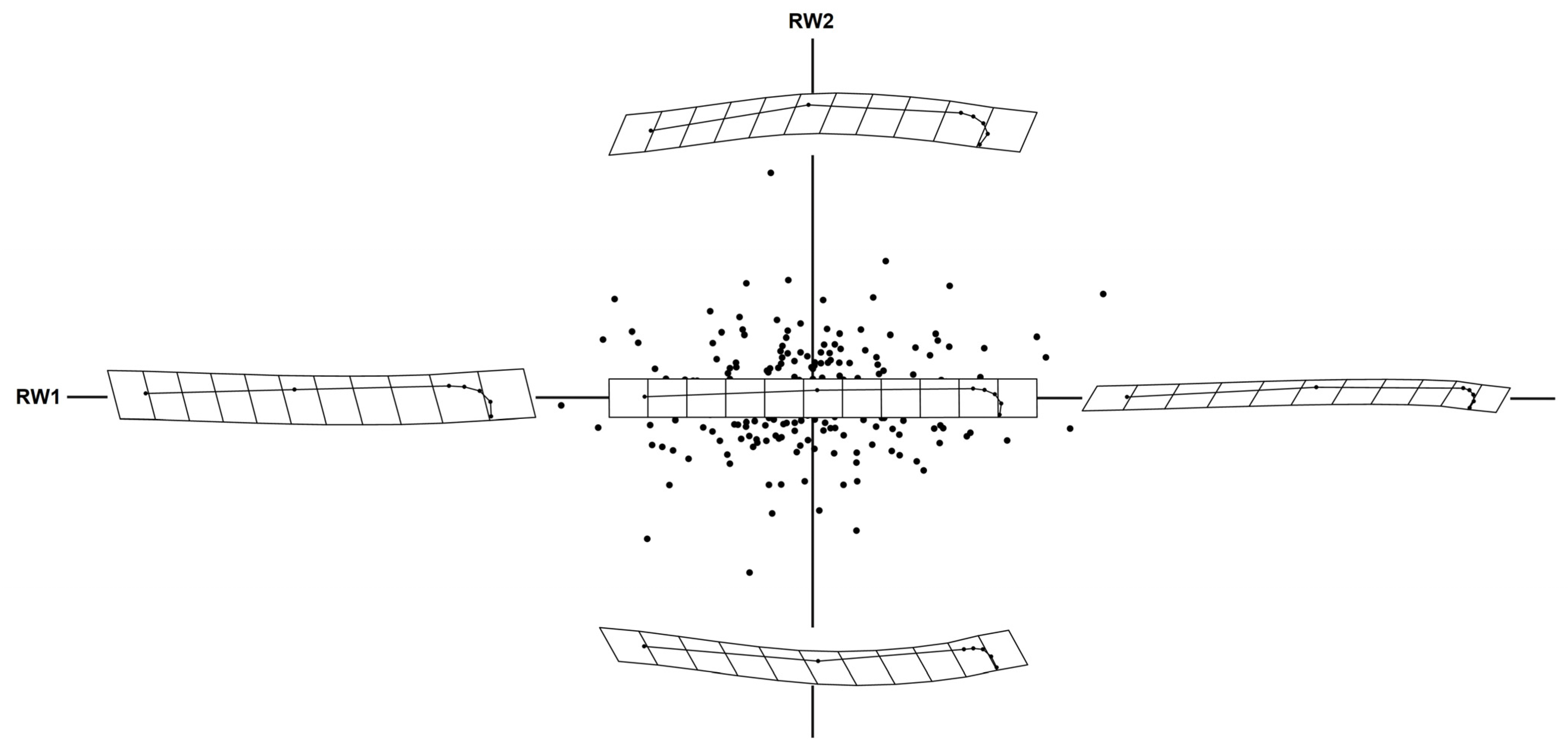
2.2. Geometric Morphometric Analysis
2.3. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Møller, A.; Milinski, M.; Slater, P. Stress and Behavior (Advances in the Study of Behavior); Academic Press: London, UK, 1998; Volume 27. [Google Scholar]
- Parsons, P. Fluctuating asymmetry: A biological monitor of environmental and genomic stress. Heredity 1992, 68, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Parasitism and developmental instability of hosts: A review. Oikos 1996, 77, 189–196. [Google Scholar] [CrossRef]
- Van Valen, L. A study of fluctuating asymmetry. Evolution 1962, 16, 125–142. [Google Scholar] [CrossRef]
- Ludwig, W. Das Rechts-Links-Problem im Tierreich und Beim Menschen: Mit Einem Anhang Rechts-Links-MERKMALE der Pflanzen; Springer: Berlin/Heidelberg, Germany, 2013; Volume 27. [Google Scholar]
- Møller, A. Fluctuating asymmetry in male sexual ornaments may reveal male quality. Mammal Rev. 1990, 5, 121–172. [Google Scholar] [CrossRef]
- Andersson, M. Sexaul selection, natural selection, and quality advertisement. Biol. J. Linn. Soc. 1982, 17, 375–393. [Google Scholar] [CrossRef]
- Roldan, E.R.S.; Cassinello, J.; Abaigar, T.; Gomendio, M. Inbreeding, fluctuating asymmetry, and ejaculate quality in an endangered ungulate. Proc. R. Soc. London. Ser. B Biol. Sci. 1998, 265, 243–248. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry as a measure of developmental stability: Implications of non-normal distributions and power of statistical tests. Acta Zool. Fenn. 1992, 191, 13. [Google Scholar]
- Leamy, L. Morphometric studies in inbred and hybrid house mice. V. Directional and fluctuating asymmetry. Am. Nat. 1984, 123, 579–593. [Google Scholar] [CrossRef]
- Graham, J.H.; Freeman, D.C.; Emlen, J.M. Antisymmetry, directional asymmetry, and dynamic morphogenesis. In Developmental instability: Its Origins and Evolutionary Implications; Springer: Berlin/Heidelberg, Germany, 1994; pp. 123–139. [Google Scholar]
- Palmer, A.R. Animal asymmetry. Curr. Biol. 2009, 19, R473–R477. [Google Scholar] [CrossRef]
- Hansen, T.F.; Pélabon, C.; Houle, D. Heritability is not Evolvability. Evol. Biol. 2011, 38, 258–277. [Google Scholar] [CrossRef]
- Falconer, D.S. Introduction to Quantitative Genetics; Pearson Education India: Noida, India, 1996. [Google Scholar]
- Houle, D. Comparing evolvability and variability of quantitative traits. Genetics 1992, 130, 195–204. [Google Scholar] [CrossRef]
- Meek, S.E. New Species of Fishes from Costa Rica; Field Museum of Natural History: Chicago, IL, USA, 1912. [Google Scholar] [CrossRef]
- Rosen, D.E.; Bailey, R.M. The poeciliid fishes (Cyprinodontiformes): Their structure, zoogeography, and systematics. Bull. Am. Mus. Nat. Hist. 1963, 126, 1–17. [Google Scholar]
- Langerhans, R.B. Genital evolution. Ecol. Evol. Poeciliid Fishes 2011, 572, 228–240. [Google Scholar]
- Rosen, D.E.; Gordon, M. Functional anatomy and evolution of male genitalia in Poeciliid fishes. Zool. Sci. Contrib. N. Y. Zool. Soc. 1953, 38, 1–47. [Google Scholar] [CrossRef]
- Bussing, W.A. Freshwater Fishes of Costa Rica/Peces de Las Aguas Continentales de Costa Rica, 2nd ed.; Editorial de la Universidad de Costa Rica: San José, Costa Rica, 1998. [Google Scholar]
- Johnson, E.S.; Nielsen, M.-E.; Johnson, J.B. Does asymmetrical gonopodium morphology predict lateralized behavior in the fish Xenophallus umbratilis? Front. Ecol. Evol. 2020, 8, 606856. [Google Scholar] [CrossRef]
- AVMA. AVMA Guidelines on Euthanasia, Formerly Report of the AVMA Panel on Euthanasia. 20; AVMA: Washington, DC, USA, 2007. [Google Scholar]
- Nickum, J.; Bart, H., Jr.; Bowser, P.; Greer, I.; Hubbs, C.; Jenkins, J.; MacMillan, J.; Rachlin, J.; Rose, J.; Sorensen, P. Guidelines for the Use of Fishes in Research; American Fisheries Society: Bethesda, MD, USA, 2004. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar] [CrossRef]
- Olympus. Olympus CellSens Software, RRID:SCR_014551; Olympus: Tokyo, Japan, 2016.
- Rohlf, F.J. The tps series of software. Hystrix Ital. J. Mammal. 2015, 26, 9–12. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Slice, D. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst. Biol. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Roth-Monzón, A.J.; Belk, M.C.; Zúñiga-Vega, J.J.; Johnson, J.B. Beyond pairwise interactions: Multispecies character displacement in Mexican freshwater fish communities. Am. Nat. 2020, 195, 983–996. [Google Scholar] [CrossRef]
- Williams, T.J.; Johnson, J.B.; Belk, M.C. Interaction between predation environment and diet constrains body shape in Utah chub, Gila atraria (Cypriniformes: Cyprinidae). Biol. J. Linn. Soc. 2017, 122, 147–156. [Google Scholar] [CrossRef]
- Mosimann, J.E. Size allometry: Size and shape variables with characterizations of the lognormal and generalized gamma distributions. J. Am. Stat. Assoc. 1970, 65, 930–945. [Google Scholar] [CrossRef]
- Searle, P.C.; Mercer, M.; Habit, E.; Belk, M.C. Ontogenetic shape trajectory of Trichomycterus areolatus varies in response to water velocity environment. PLoS ONE 2021, 16, e0252780. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J.D.; Reznick, D.N. Evolution of juvenile growth rates in female guppies (Poecilia reticulata): Predator regime or resource level? Proc. R. Soc. B Biol. Sci. 2005, 272, 333–337. [Google Scholar] [CrossRef]
- Reznick, D.; Hrbek, T.; Caura, S.; De Greef, J.; Roff, D. Life history of Xenodexia ctenolepis: Implications for life history evolution in the family Poeciliidae. Biol. J. Linn. Soc. 2007, 92, 77–85. [Google Scholar] [CrossRef]
- Wiper, M.L. Evolutionary and mechanistic drivers of laterality: A review and new synthesis. Laterality 2017, 22, 740–770. [Google Scholar] [CrossRef]
- Nielsen, M.-E.; Johnson, E.S.; Johnson, J.B. Does male gonopodial morphology affect male-female mating positioning in the livebearing fish Xenophallus umbratilis? PLoS ONE 2023, 18, e0281267. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Museum ID | Tributary/River | Coordinates |
|---|---|---|---|
| 1 | 009294c | Rio Corinto | 10°12.674′ N 83°53.114′ W |
| 2 | 009301b | Rio Esquivetto | 10°41.231′ N 85°04.002′ W |
| 3 | 009302 | Trib. to Rio Bijagua | 10°43.887′ N 85°03.318′ W |
| 4 | 009310 | Rio Tenerio | 10°41.285′ N 85°04.561′ W |
| 5 | 009320 | Trib. to Rio Bijagua | 10°43.453′ N 85°03.982′ W |
| 6 | 009325 | Quebrada Hormiguero | 10°41.454′ N 85°05.019′ W |
| 7 | 009338c | Quebrada La Palma | 10°33.614′ N 84°56.442′ W |
| 8 | 009339 | Quebrada Hormiguero | 10°41.445′ N 85°05.036′ W |
| 9 | 009340 | Quebrada Isabel | 10°38.387′ N 84°50.757′ W |
| 10 | 009354 | Quebrada Azul | 10°29.955′ N 84°59.138′ W |
| Response Variable | Random Effect | Predictor Variable | |
|---|---|---|---|
| Model 1 | Relative Warps | -- | Centroid Size |
| Index | |||
| Centroid Size × Index | |||
| Model 2 | Relative Warps | -- | Locality |
| Centroid Size | |||
| Centroid Size × Locality | |||
| Index | |||
| Locality × Index | |||
| Centroid Size × Index | |||
| Locality × Centroid Size × Index | |||
| Model 3 | Relative Warps | Locality | Morph |
| Centroid Size | |||
| Centroid Size × Morph | |||
| Index | |||
| Morph × Index | |||
| Centroid Size × Index | |||
| Morph × Centroid Size × Index |
| Predictor Variable | Degrees of Freedom | F-Value | p-Value | |
|---|---|---|---|---|
| Model 1 | Centroid Size | 1, 702 | 6.44 | 0.0113 |
| Index | 5, 657 | 4.9 | 0.0002 | |
| Centroid Size × Index | 5, 657 | 4.82 | 0.0002 | |
| Model 2 | Locality | 9, 794 | 1.02 | 0.4226 |
| Centroid Size | 1, 794 | 1.25 | 0.2365 | |
| Centroid Size × Locality | 9, 794 | 1.04 | 0.4092 | |
| Index | 5, 659 | 2.04 | 0.0718 | |
| Locality × Index | 45, 1183 | 0.98 | 0.5024 | |
| Centroid Size × Index | 5, 659 | 2.07 | 0.0671 | |
| Locality × Centroid Size × Index | 45, 1183 | 0.98 | 0.5188 | |
| Model 3 | Morph | 1, 704 | 0.01 | 0.9238 |
| Centroid Size | 1, 619 | 5.09 | 0.0244 | |
| Centroid Size × Morph | 1, 703 | 0.04 | 0.8391 | |
| Index | 5, 653 | 3.26 | 0.0065 | |
| Morph × Index | 5, 653 | 1.39 | 0.2273 | |
| Centroid Size × Index | 5, 653 | 3.07 | 0.0095 | |
| Morph × Centroid Size × Index | 5, 653 | 1.23 | 0.0095 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, M.-E.; Johnson, J.B. Quantifying Shape Variation in an Antisymmetrical Trait in the Tropical Fish Xenophallus umbratilis. Symmetry 2023, 15, 489. https://doi.org/10.3390/sym15020489
Nielsen M-E, Johnson JB. Quantifying Shape Variation in an Antisymmetrical Trait in the Tropical Fish Xenophallus umbratilis. Symmetry. 2023; 15(2):489. https://doi.org/10.3390/sym15020489
Chicago/Turabian StyleNielsen, Mary-Elise, and Jerald B. Johnson. 2023. "Quantifying Shape Variation in an Antisymmetrical Trait in the Tropical Fish Xenophallus umbratilis" Symmetry 15, no. 2: 489. https://doi.org/10.3390/sym15020489