
Asymmetries of Cerebellar Lobe in the Genus Homo


1
Joint International Research Laboratory of Environmental and Social Archaeology, Shandong University, Qingdao 266237, China
2
Institute of Cultural Heritage, Shandong University, Qingdao 266237, China
3
Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China
4
CAS Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Beijing 100044, China
*
Author to whom correspondence should be addressed.
Symmetry 2021, 13(6), 988; https://doi.org/10.3390/sym13060988
Submission received: 29 April 2021
/
Revised: 28 May 2021
/
Accepted: 31 May 2021
/
Published: 2 June 2021
(This article belongs to the Special Issue Symmetry in Human Evolution, from Biology to Behaviours)
Abstract
:The endocast was paid great attention in the study of human brain evolution. However, compared to that of the cerebrum, the cerebellar lobe is poorly studied regarding its morphology, function, and evolutionary changes in the process of human evolution. In this study, we define the major axis and four measurements to inspect possible asymmetric patterns within the genus Homo. Results show that significant asymmetry is only observed for the cerebellar length in modern humans and is absent in Homo erectus and Neanderthals. The influence of occipital petalia is obscure due to the small sample size for H. erectus and Neanderthals, while it has a significant influence over the asymmetries of cerebellar height and horizontal orientation in modern humans. Although the length and height of the Neanderthal cerebellum are comparable to that of modern humans, its sagittal orientation is closer to that of H. erectus, which is wider than that of modern humans. The cerebellar morphological difference between Neanderthals and modern humans is suggested to be related to high cognitive activities, such as social factors and language ability.
1. Introduction
Endocast, or brain endocast, is the cast made of the interior of the neurocranium of a skull [1]. Endocast is the only agent to investigate how the human brain evolved physically in the process of evolution [2] regarding its volume [3,4,5,6,7], surface features [8,9,10,11], or size and shape [12,13,14,15,16].
Asymmetry of the brain, as one of the most debated questions, exhibits at different levels, such as the Broca’s area [17,18], perisylvian region [19], central sulcus, cortical and subcortical regions, lobes, and hemispheres [20,21]. Brain asymmetry is often related to functional and evolutionary significance; for example, petalia and the Yakovlevian torque is a geometric distortion of the brain hemisphere, in which the left occipital lobe and the right frontal lobe are wider and longer than the opposite side [21]. While a combination of left occipital and right frontal petalia is common in modern humans and fossilized hominins and is regarded as evidence of right-handedness [22,23,24], such observations are less exaggerated or rarely consistent in great apes and other primates [15,25]. Also, capuchins display a leftward frontal petalia [26] while macaques show the rightward frontal petalia [27], and both were absent of a left occipital petalia.
Compared to that of the cerebrum, the cerebellum receives less attention and is still poorly understood. However, with the advances of neuroimaging technology and theoretical innovation, there is increasing study of the cerebellum regarding its morphology, function, and evolutionary changes. Moreover, evidence from neuroimaging uncovers the cerebellum as the “missing link” in many cognitive domains [28]. Sereno, et al. [29] revealed in a recent study that the cerebellar cortex covers almost 80% of the surface area of the cerebral cortex, and the expansion of the human cerebellar surface area even exceeds the cerebral cortex when compared to that of monkeys. Also, neuroimaging evidence suggests that the function of the cerebellum is also highly involved in cognitive and social activities [30,31,32,33], which is also supported by clinical studies, such as developmental dyslexia [34,35].
The evolutionary changes of the cerebellum were also hotly debated. Studies suggest that the cerebellum shows a similar asymmetric pattern as the cerebrum (the left-occipital, right-frontal petalia), with larger anterior lobules on the right side and larger posterior lobules on the left side [36,37]. Compared to that of humans, the cerebellar torque is opposite in chimpanzees [26], while absent in capuchins. MacLeod, Zilles, Schleicher, Rilling, and Gibson [31] found that hominins have a great increase in the lateral cerebellum compared to that of monkeys. It is also a popular idea that the cerebrum and cerebellum underwent several expansions and reorganizations in the process of human evolution [2]. Weaver [38] came up with a hypothesis that the evolution of human cerebellar/neocortical occurred in three stages; the first stage as early encephalization with an expansion of the neocortex during Early-to-Middle Pleistocene; the second stage as the dramatic encephalization primarily happened to the neocortex in Middle-to-Late Pleistocene humans, accompanied by a proliferation of cultural objects as well as an increase in complex behaviors, such as pyrotechnology and prepared core techniques, and the third stage happened in the late Late Pleistocene and Holocene, with an increase in cognitive efficiency as a result of expanded cerebellar capacity, which is the reason why modern humans can do more without an increase in net brain volume. Cerebellar specialization is thought to be an important component in the evolution of humanity’s advanced technological capacities and languages [39,40].
The cerebellum is classically divided into three lobes, namely, the anterior lobe, the posterior lobe, and the flocculonodular lobe; it also has three major surfaces, with the superior surface toward the tentorium cerebelli, the posterior surface toward the internal occipital bone, and the anterior surface toward the petrous pyramid [41]. The cerebellar lobe reconstructed from the cerebellar fossa is surrounded by transverse sinus, sigmoid sinus, and occasionally occipital and marginal sinus. Accurate and homologous landmarks are important in the study of endocasts, such as in the case of Taung australopithecine endocast, in which a vague position of the lunate sulcus caused great debate [1,42,43,44,45,46,47,48]. Unlike the cerebrum, there are no gyrus or sulcus that exist on the surface of the cerebellum, and hence, it is difficult to identify landmarks to measure.
In this study, we tentatively define the major axis and four measurements on the cerebellum to achieve a more accurate between-group comparison. The main aim of this study is (1) to test whether certain asymmetric patterns existed within the genus Homo; (2) to find out possible factors that affected the asymmetry, and to (3) provide morphological evidence for cerebellar reorganization and cognitive increases in the genus Homo.
2. Materials and Methods
2.1. Materials
A total of 45 specimens were used in this study (as illustrated in Table 1), including Homo erectus (n = 11), Homo neanderthalensis (n = 4), and extant Homo sapiens (modern humans, n = 30). H. erectus specimens were mostly sourced from Asia and Indonesia to avoid significant regional differences. The 3D data of the fossil hominins were laser-scanned from endocast models housed in the American Museum of Natural History (AMNH) and the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) with a resolution of 0.5 mm or higher. Endocasts of extant H. sapiens were reconstructed from CT scans with a resolution of 160 μm. Frontal pole to occipital pole length was also measured in this study and compared to that of those from literature to ascertain whether no great discrepancy existed.
2.2. Cerebellar Metrics
Due to lacking prominent anatomical features, homologous landmarks on the cerebellar lobe are difficult to recognize and define. Although previous studies used transverse sinus and sigmoid sinus defining measurements of the cerebellum [1], the great variation suggests that the sinus was not an adequate reference.
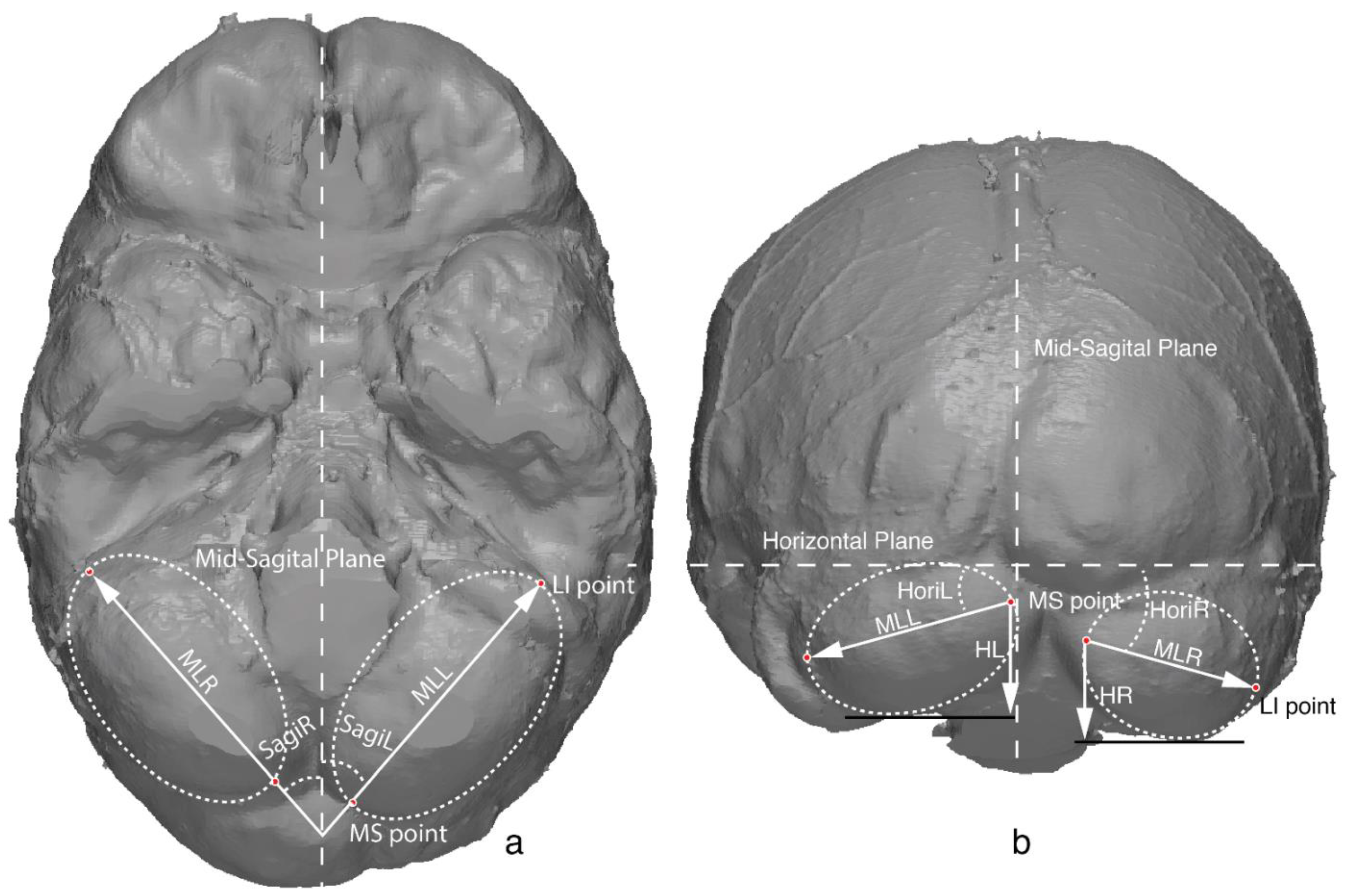
Considering the ellipsoid shape of the cerebellar lobe, the size and shape can be best depicted by the major axis and two points it passes through (as illustrated in Figure 1). Here, we define the major axis as a straight line that divides the cerebellar lobe into two halves from both the inferior view and the posterior view. The point that the major axis passes through at the anterior part is defined as the most lateral and inferior point (LI point), which in most cases is medial to the sigmoid sinus. The point that the major axis passes through at the posterior part is defined as the most medial and superior point (MS point), which is close to the internal occipital protuberance point and is often asymmetric because of the occipital petalia. With the endocast at the standard position using the front pole to occipital pole as the horizontal plane [1], four measurements were defined upon the major axis (as illustrated in Figure 1, Table 2), namely, the cerebellar length, cerebellar height, sagittal orientation (the orientation of the major axis relative to the sagittal plane), and horizontal orientation (orientation of the major axis relative to the horizontal plane). Asymmetric parameters were also calculated as the difference between the left and right sides of the same measurement (L–R) [15], namely, the ML.lr, H.lr, Sagi.lr, and Hori.lr. The determination of the landmarks and the measurements were performed in Rapidform XOR3.
2.3. Descriptive Statistics of the Asymmetries
To demonstrate overall asymmetries of the cerebellum, all the measurements were summarized regarding different populations and measurements, with mean value, standard deviation, and coefficient of variation (CV) being presented. Paired t-test was conducted to test the difference between the two sides with bootstrap considering the small sample size.
2.4. Analysis of Covariance
Cerebellar size and shape may be influenced by occipital petalia and brain size (allometry) [49]. Therefore, those two factors were recorded and analyzed in this study. Endocasts with left occipital petalia, right occipital petalia, or equally bilateral situation are recorded as L, R, and B, respectively. To test how cerebellar asymmetries were influenced by those two factors, analysis of covariance (ANCOVA) was conducted within each population group, with asymmetric parameters as the dependent variable, natural log-transformed cranial capacity as the covariate, and occipital petalia as the independent variable.
3. Error Evaluation
To avoid interobserver error, all measurements were measured by the same author Y.Z. Also, to assess intraobserver error, three specimens were chosen and measured six times repeatedly on six different days. The coefficient of variation (CV) is calculated with the repeated measurements (as illustrated in Table 3). The intraobserver error is well controlled within most of the measurements except for the horizontal angle. All of the measurements were included in the formal analysis, although the results of the horizontal angle should be taken with great caution.
4. Results
4.1. Description of Cerebellar Asymmetries
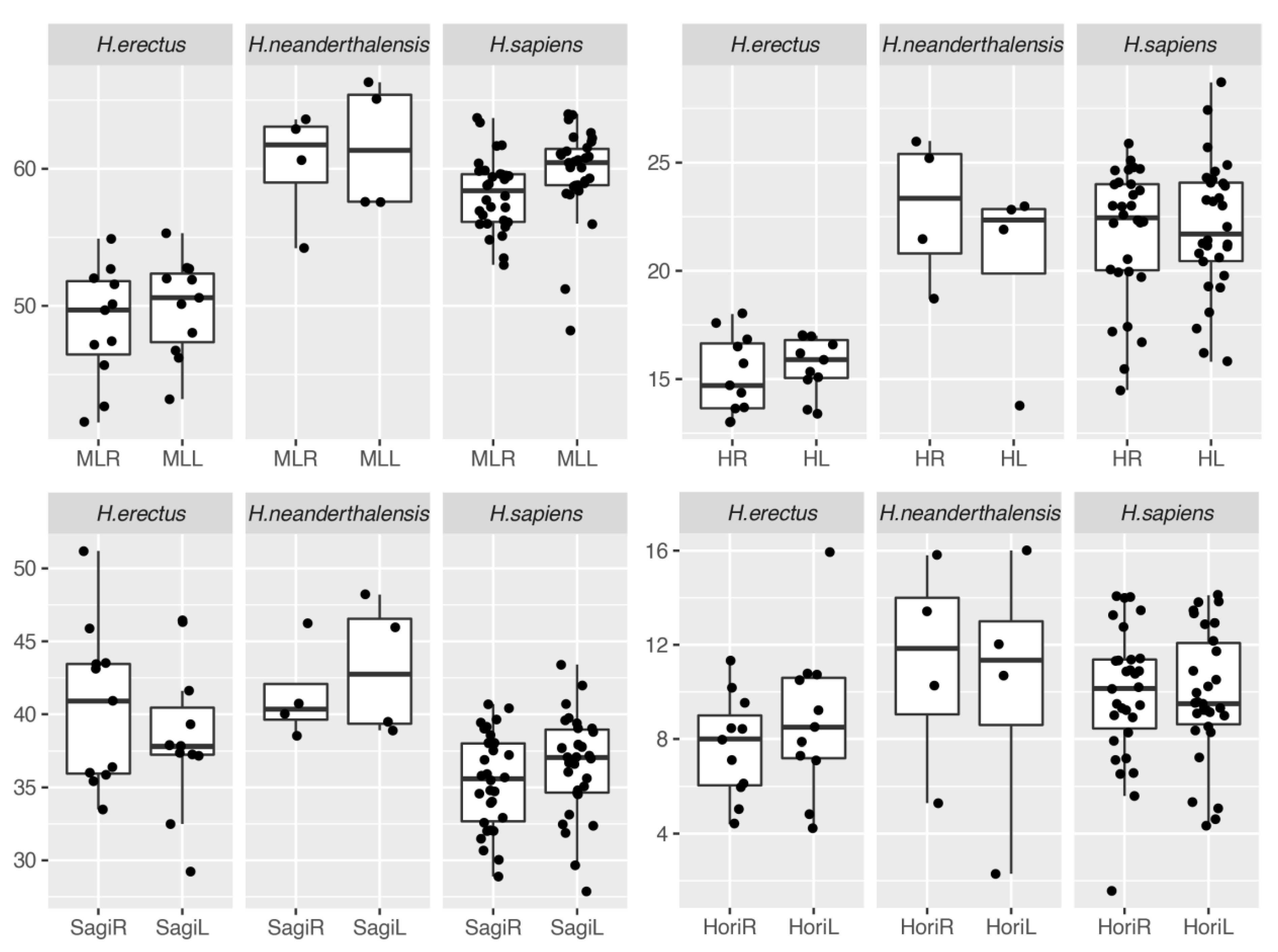
Results of the descriptive statistics and bootstrapped t-test were shown in Table 4. No significant asymmetry was observed among the four measurements within H. erectus and H. neanderthalensis. However, the result of Neanderthals should be taken with caution as only a small sample size is available. Within the H. sapiens group, only the cerebellar length is significantly larger on the left side.
Together with the boxplot (as illustrated in Figure 2), a preliminary evolutionary change of the cerebellar metrics can be summarized. There is an obvious growth of the cerebellar length and cerebellar height for Neanderthals and modern humans over the H. erectus. However, the sagittal orientation of the cerebellar is wider for Neanderthals and H. erectus while this value is quite small for modern humans. For the horizontal orientation, Neanderthals and modern humans are relatively steep while it is rather flat in H. erectus. The CVs of horizontal orientation are high in the process of error evaluation and may contribute to within-group variation here.
4.2. ANCOVA
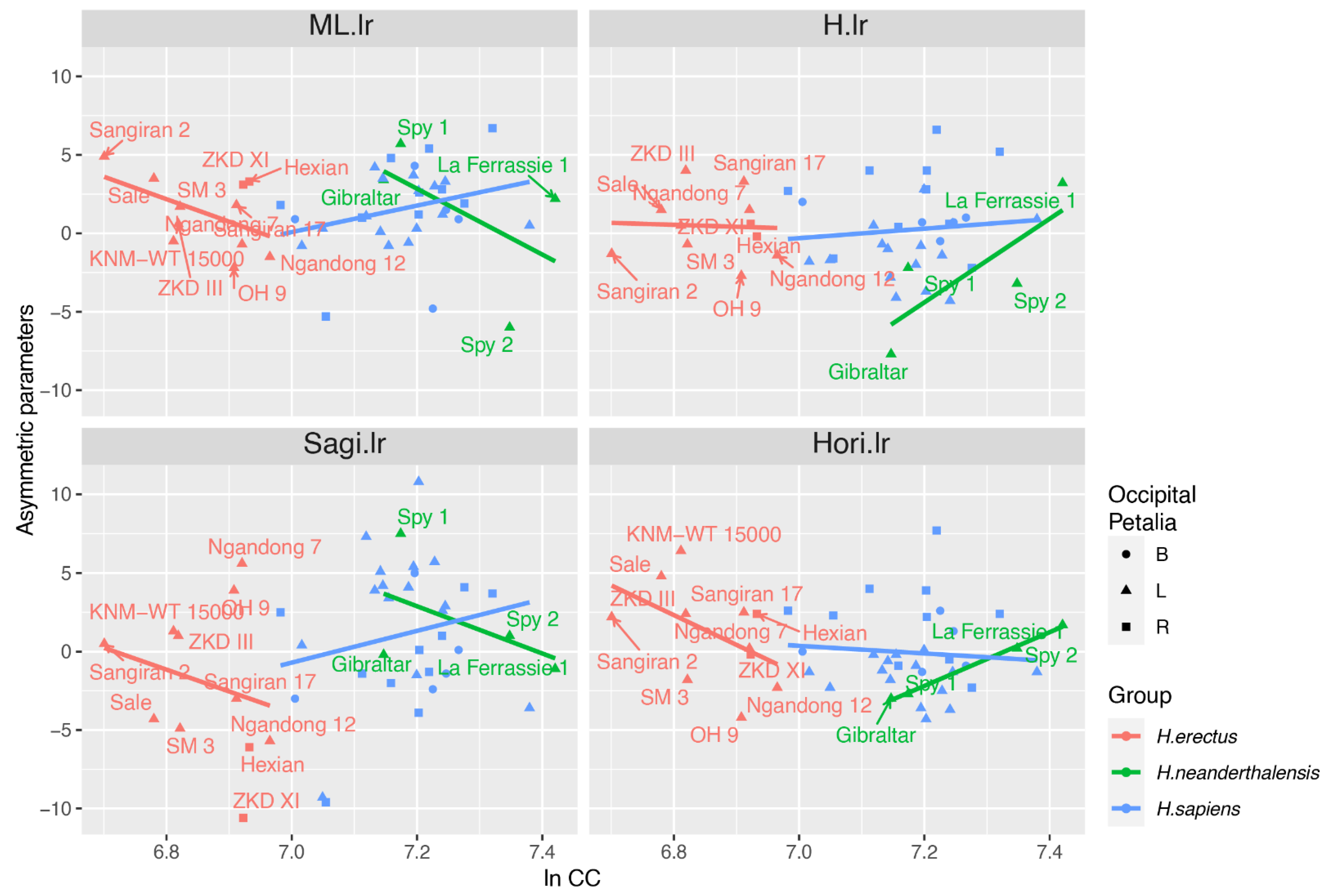
In H. erectus, the ANCOVA (as illustrated in Table 5) revealed a significant relationship between cerebellar length asymmetric parameter (ML.lr) and both cranial capacity and occipital petalia while other parameters had no such relationship. From the scatterplot (as illustrated in Figure 3), negative allometry can be observed between ML.lr and cranial capacity in H. erectus, indicating that individuals with small brain size will have a larger cerebellar length on the left side. However, only two specimens of H. erectus have right occipital petalia (as illustrated in Figure 3), making the relationship between cerebellar length and occipital petalia questionable.
Within H. sapiens, cranial capacity does not affect the asymmetric parameter significantly. Occipital petalia had a significant influence on cerebellar height asymmetric parameter (H.lr) and horizontal orientation asymmetric parameter (Hori.lr). Endocasts with right occipital petalia tend to have large positive H.lr and Hori.lr, indicating a tall and steep-orientated cerebellar lobe on the left side.
Because all of the four Neanderthal specimens were left occipital petalia, we performed linear regression instead of the ANCOVA in Neanderthals. Results show that Hori.lr is significantly affected by the cranial capacity. A large brain size would be accompanied by a steep cerebellar lobe, as revealed in the scatterplot (as illustrated in Figure 3).
5. Discussion
5.1. Cerebellar Asymmetric Pattern
The cerebellar asymmetry is only observed in H. sapiens while absent in H. erectus and possibly H. neanderthalensis. In H. sapiens, the cerebellar length is significantly longer on the left side, while its height, sagittal orientation, and horizontal orientation do not differ between left and right side (as illustrated in Figure 4).
Further analysis found that the petalia and cranial capacity did not influence the cerebellar length and sagittal orientation in H. sapiens. However, the occipital petalia is significantly related to cerebellar height and horizontal orientation in H. sapiens. Individuals with right occipital petalia would have high and steep orientated cerebellar on the left side (as illustrated in Figure 4). The prevalence of cerebellum contralateral to the occipital lobe is suggested to be a spatial compensation [52,53], supported by the fact that the occipital petalia can be used as a predictive sign for the transverse sinus [54]. This is not repeated in the cerebellar length and sagittal orientation, indicating that the occipital petalia only affects the superior–inferior dimension instead of the anterior–posterior and medial–lateral dimension of the cerebellum.
5.2. Cerebellar Asymmetry in the Genus Homo
Although cerebellar expansion can be traced back to great apes [38], the asymmetries of the cerebellum appear quite recently in H. sapiens, which is absent in H. erectus and possibly H. neanderthalensis. Also, this differs from the fact that the cerebral laterality is distinguished in early hominins compared to that of the great apes [22,25]. We provide more evidence for the difference of evolutionary trajectories between the cerebrum and cerebellum.
A flat parietal lobe, elongated occipital lobe, and flat cerebellar lobe are thought to be unique features among Neanderthal endocasts [12]. Compared to that of Neanderthals, the endocast of modern humans is much more globular, including the parietal and the cerebellum lobe [55]. The globularity of the endocast of modern humans mainly originates from the parietal expansion rather than the cerebellum [12]. Congruent with previous studies, we find that modern humans do not differ from Neanderthals much at the cerebellar length and height (without size correction), and horizontal orientation. Our results support the idea that the tentorium cerebelli prevented the horizontal dimensions, such as the cerebellar height and horizontal orientation, from diverging greatly between Neanderthals and modern humans [12]. Meanwhile, the sagittal orientation is significantly narrower in modern humans when compared to that of Neanderthals. The small sample size of the Neanderthals aside, this may reflect a species-specific feature. If so, such difference possibly occurs during the “globularization-phase” when critical features of the human brain were established [55]. This might provide new evidence for the different rates and timing of brain development between Neanderthals and modern humans. However, such a statement needs more study and ontogenetic evidence. We suggest this to be related to higher cognitive abilities, such as social factors [56] and language abilities [57], that differ mostly between H. sapiens and Neanderthals [58].
5.3. Limitations
Considering the difficulty in obtaining homologous landmarks on the cerebellum, this is still a very preliminary study. The morphological information obtained is limited, and the uncertainty of the landmarks also introduced errors, especially in the measurement of the horizontal orientation. Further studies with landmark-free methods are suggested to reduce measurement error and extract more information from the whole cerebellar surface. Also, the basilar part of the cranium is rarely well preserved in human fossils and led to the small sample size in the analysis, which also prohibited us from obtaining convincing results.
Author Contributions
Conceptualization, Y.Z. and X.W.; methodology, Y.Z.; software, Y.Z. and X.W.; validation, Y.Z. and X.W.; formal analysis, Y.Z.; resources, Y.Z. and X.W., data curation, Y.Z. and X.W.; writing—original draft preparation, Y.Z.; writing—review and editing, Y.Z.; visualization, Y.Z.; funding acquisition, Y.Z. and X.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Key Laboratory of Vertebrate Evolution and Human Origins, IVPP, CAS (NO. LVEHO20005), the Strategic Priority Research Program of Chinese Academy of the Sciences (XDB26000000), and the Shandong University Humanities and Social Science Major Project (No. 19RWZD08).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used in the study are available from the authors upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Holloway, R.L.; Broadfield, D.C.; Yuan, M.S.; Schwartz, J.H.; Tattersall, I. The Human Fossil Record, Brain Endocasts--The Paleoneurological Evidence, 1st ed.; Wiley-Liss: New York, NY, USA, 2004; p. 315. [Google Scholar]
- Holloway, R.L. Evolution of the Human Brain; Lock, A., Peters, C.R., Eds.; Clarendon Press: Oxford, UK, 1996; pp. 74–125. [Google Scholar]
- Holloway, R.L. Cranial capacity, neural reorganization, and hominid evolution: A search for more suitable parameters. Am. Anthrop. 1966, 68, 103–121. [Google Scholar]
- Holloway, R.L. New Endocranial Values for the Australopithecines. Nature 1970, 227, 199–200. [Google Scholar] [CrossRef]
- Holloway, R.L. New Endocranial Values for the East African Early Hominids. Nature 1973, 243, 97–99. [Google Scholar] [CrossRef]
- Conroy, G.C.; Vannier, M.W.; Tobias, P.V. Endocranial features of Australopithecus africanus revealed by 2- and 3-D computed tomography. Science 1990, 247, 838–841. [Google Scholar] [CrossRef]
- Conroy, G.C.; Weber, G.W.; Seidler, H.; Tobias, P.V.; Kane, A.; Brunsden, B. Endocranial capacity in an early hominid cranium from Sterkfontein, South Africa. Science 1998, 280, 1730–1731. [Google Scholar] [CrossRef] [PubMed]
- Falk, D.; Conroy, G.C. The cranial venous sinus system in Australopithecus afarensis. Nature 1983, 306, 779–781. [Google Scholar] [CrossRef]
- Falk, D. Meningeal arterial patterns in great apes: Implications for hominid vascular evolution. Am. J. Phys. Anthropol. 1993, 92, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Weidenreich, F. The ramification of the middle meningeal artery in fossil hominids: And its bearing upon phylogenetic problems. Palaeontol. Sin. 1938, 110, 1–16. [Google Scholar]
- Bruner, E.; Sherkat, S. The middle meningeal artery: From clinics to fossils. Child’s Nerv. Syst. 2008, 24, 1289–1298. [Google Scholar] [CrossRef]
- Bruner, E. Geometric morphometrics and paleoneurology: Brain shape evolution in the genus Homo. J. Hum. Evol. 2004, 47, 279–303. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Pedro, A.S.; Bruner, E.; Gunz, P.; Neubauer, S. A morphometric comparison of the parietal lobe in modern humans and Neanderthals. J. Hum. Evol. 2020, 142, 102770. [Google Scholar] [CrossRef]
- Wu, X.-j.; Bruner, E. The endocranial anatomy of maba 1. Am. J. Phys. Anthropol. 2016, 160, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Balzeau, A.; Holloway, R.L.; Grimaud-Hervé, D. Variations and asymmetries in regional brain surface in the genus Homo. J. Hum. Evol. 2012, 62, 696–706. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E.; Holloway, R.L.; Prima, S.; Grimaud-Hervé, D. Variations in size, shape and asymmetries of the third frontal convolution in hominids: Paleoneurological implications for hominin evolution and the origin of language. J. Hum. Evol. 2014, 76, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, C.C.; Broadfield, D.C.; Holloway, R.L.; Gannon, P.J.; Hof, P.R. Variability of Broca’s area homologue in African great apes: Implications for language evolution. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2003, 271, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Bruner, E.; Holloway, R.L. A bivariate approach to the widening of the frontal lobes in the genus Homo. J. Hum. Evol. 2010, 58, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Xiang, L.; Crow, T.J.; Leroy, F.; Rivière, D.; Mangin, J.-F.; Roberts, N. Measurement of Sylvian Fissure asymmetry and occipital bending in humans and Pan troglodytes. Neuroimage 2019, 184, 855–870. [Google Scholar] [CrossRef] [Green Version]
- LeMay, M. Morphological Cerebral Asymmetries of Modern Man, Fossil Man, and Nonhuman Primate. Ann. N. Y. Acad. Sci. 1976, 280, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Rentería, M.E. Cerebral asymmetry: A quantitative, multifactorial, and plastic brain phenotype. Twin Res. Hum. Genet. 2012, 15, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Holloway, R.L.; De La Costelareymondie, M.C. Brain endocast asymmetry in pongids and hominids: Some preliminary findings on the paleontology of cerebral dominance. Am. J. Phys. Anthropol. 1982, 58, 101–110. [Google Scholar] [CrossRef]
- Galaburda, A.M.; LeMay, M.; Kemper, T.L.; Geschwind, N. Right-left asymmetrics in the brain. Science 1978, 199, 852–856. [Google Scholar] [CrossRef]
- Hervé, P.-Y.; Crivello, F.; Perchey, G.; Mazoyer, B.; Tzourio-Mazoyer, N. Handedness and cerebral anatomical asymmetries in young adult males. Neuroimage 2006, 29, 1066–1079. [Google Scholar] [CrossRef]
- Li, X.; Crow, T.J.; Hopkins, W.D.; Gong, Q.; Roberts, N. Human torque is not present in chimpanzee brain. Neuroimaging 2018, 165, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Phillips, K.A.; Sherwood, C.C. Cerebral petalias and their relationship to handedness in capuchin monkeys (Cebus apella). Neuropsychologia 2007, 45, 2398–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falk, D.; Hildebolt, C.; Cheverud, J.; Vannier, M.; Helmkamp, R.C.; Konigsberg, L. Cortical asymmetries in frontal lobes of rhesus monkeys (Macaca mulatta). J. Brain Res. 1990, 512, 40–45. [Google Scholar] [CrossRef]
- MacLeod, C. Chapter 8—The missing link: Evolution of the primate cerebellum. In Progress in Brain Research; Hofman, M.A., Falk, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 195, pp. 165–187. [Google Scholar]
- Sereno, M.I.; Diedrichsen, J.; Tachrount, M.; Testa-Silva, G.; d’Arceuil, H.; De Zeeuw, C. The human cerebellum has almost 80% of the surface area of the neocortex. Proc. Natl. Acad. Sci. USA 2020, 117, 19538–19543. [Google Scholar] [CrossRef]
- Tanabe, H.C.; Kubo, D.; Hasegawa, K.; Kochiyama, T.; Kondo, O. Cerebellum: Anatomy, Physiology, Function, and Evolution. In Digital Endocasts: From Skulls to Brains; Bruner, E., Ogihara, N., Tanabe, H.C., Eds.; Springer: Tokyo, Japan, 2018; pp. 275–289. [Google Scholar]
- MacLeod, C.E.; Zilles, K.; Schleicher, A.; Rilling, J.K.; Gibson, K.R. Expansion of the neocerebellum in Hominoidea. J. Hum. Evol. 2003, 44, 401–429. [Google Scholar] [CrossRef]
- Kim, S.; Ugurbil, K.; Strick, P. Activation of a cerebellar output nucleus during cognitive processing. Science 1994, 265, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; Fiez, J.A.; Videen, T.O.; MacLeod, A.-M.K.; Pardo, J.V.; Fox, P.T.; Petersen, S.E. Practice-related changes in human brain functional anatomy during nonmotor learning. Cereb. Cortex 1994, 4, 8–26. [Google Scholar] [CrossRef]
- Rae, C.; Harasty, J.A.; Dzendrowskyj, T.E.; Talcott, J.B.; Simpson, J.M.; Blamire, A.M.; Dixon, R.M.; Lee, M.A.; Thompson, C.H.; Styles, P. Cerebellar morphology in developmental dyslexia. Neuropsychologia 2002, 40, 1285–1292. [Google Scholar] [CrossRef]
- Eckert, M.A.; Leonard, C.M.; Richards, T.L.; Aylward, E.H.; Thomson, J.; Berninger, V.W. Anatomical correlates of dyslexia: Frontal and cerebellar findings. Brain 2003, 126, 482–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diedrichsen, J.; Balsters, J.H.; Flavell, J.; Cussans, E.; Ramnani, N. A probabilistic MR atlas of the human cerebellum. Neuroimage 2009, 46, 39–46. [Google Scholar] [CrossRef]
- Snyder, P.J.; Bilder, R.M.; Wu, H.; Bogerts, B.; Lieberman, J.A. Cerebellar volume asymmetries are related to handedness: A quantitative MRI study. Neuropsychologia 1995, 33, 407–419. [Google Scholar] [CrossRef]
- Weaver, A.H. Reciprocal evolution of the cerebellum and neocortex in fossil humans. Proc. Natl. Acad. Sci. USA 2005, 102, 3576–3580. [Google Scholar] [CrossRef] [Green Version]
- Barton, R.A. Embodied cognitive evolution and the cerebellum. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2097–2107. [Google Scholar] [CrossRef] [Green Version]
- Barton, R.A.; Venditti, C. Rapid Evolution of the Cerebellum in Humans and Other Great Apes. Curr. Biol. 2014, 24, 2440–2444. [Google Scholar] [CrossRef] [Green Version]
- Naidich, T.P.; Duvernoy, H.M.; Delman, B.N.; Sorensen, A.G.; Kollias, S.S.; Haacke, E.M. Duvernoy’s Atlas of the Human Brain Stem and Cerebellum; Springer: Vienna, Austria, 2009. [Google Scholar]
- Falk, D. Apples, oranges, and the lunate sulcus. Am. J. Phys. Anthropol. 1985, 67, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Falk, D. A reanalysis of the South African australopithecine natural endocasts. Am. J. Phys. Anthropol. 1980, 53, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Falk, D. Ape-like endocast of “ape-man” Taung. Am. J. Phys. Anthropol. 1989, 80, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Falk, D. The Taung endocast: A reply to Holloway. Am. J. Phys. Anthropol. 1983, 60, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Holloway, R.L. Revisiting the South African Taung australopithecine endocast: The position of the lunate sulcus as determined by the stereoplotting technique. Am. J. Phys. Anthropol. 1981, 56, 43–58. [Google Scholar] [CrossRef]
- Holloway, R.L. The taung endocast and the lunate sulcus: A rejection of the hypothesis of its anterior position. Am. J. Phys. Anthropol. 1984, 64, 285–287. [Google Scholar] [CrossRef]
- Holloway, R.L.; Clarke, R.J.; Tobias, P.V. Posterior lunate sulcus in Australopithecus africanus: Was Dart right? Comptes Rendus Palevol 2004, 3, 287–293. [Google Scholar] [CrossRef]
- Rightmire, G.P. Homo erectus and middle pleistocene hominins: Brain size, skull form, and species recognition. J. Hum. Evol. 2013, 65, 223–252. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Enlow, D.H.; Hans, M. Essentials of Facial Growth; W.B. Saunders Company: Philadelphia, PA, USA, 1996. [Google Scholar]
- Enlow, D.H.; Kuroda, T.; Lewis, A.B. The morphological and morphogenetic basis for craniofacial form and pattern. Angle Orthod. 1971, 41, 161–188. [Google Scholar] [PubMed]
- Arsava, E.Y.; Arsava, E.M.; Oguz, K.K.; Topcuoglu, M.A. Occipital petalia as a predictive imaging sign for transverse sinus dominance. Neurol. Res. 2019, 41, 306–311. [Google Scholar] [CrossRef]
- Gunz, P.; Neubauer, S.; Golovanova, L.; Doronichev, V.; Maureille, B.; Hublin, J.-J. A uniquely modern human pattern of endocranial development. Insights from a new cranial reconstruction of the Neandertal newborn from Mezmaiskaya. J. Hum. Evol. 2012, 62, 300–313. [Google Scholar] [CrossRef]
- Burke, A. Spatial abilities, cognition and the pattern of Neanderthal and modern human dispersals. Quat. Int. 2012, 247, 230–235. [Google Scholar] [CrossRef]
- Lieberman, P. The evolution of human speech-Its anatomical and neural bases. Curr. Anthropol. 2007, 48, 39–66. [Google Scholar] [CrossRef] [Green Version]
- Kochiyama, T.; Ogihara, N.; Tanabe, H.C.; Kondo, O.; Amano, H.; Hasegawa, K.; Suzuki, H.; Ponce de León, M.S.; Zollikofer, C.P.E.; Bastir, M.; et al. Reconstructing the Neanderthal brain using computational anatomy. Sci. Rep. 2018, 8, 6296. [Google Scholar] [CrossRef]
Figure 1.
Diagram of major axis and four measurements of cerebellum on endocast from inferior (a) and posterior view (b). MLL and MLR, cerebellar length on the left and right side; MS point, medial-superior point; LI point, lateral-inferior point; SagiL and SagiR, orientation of major axis relative to sagittal and horizontal plane; HL and HR, cerebellar height on left and right side.
Figure 1.
Diagram of major axis and four measurements of cerebellum on endocast from inferior (a) and posterior view (b). MLL and MLR, cerebellar length on the left and right side; MS point, medial-superior point; LI point, lateral-inferior point; SagiL and SagiR, orientation of major axis relative to sagittal and horizontal plane; HL and HR, cerebellar height on left and right side.

Figure 2.
Boxplot of cerebellar metrics.

Figure 3.
Allometric trend of asymmetric parameters in three populations; B, bilateral; L, left occipital petalia; R, right occipital petalia.
Figure 3.
Allometric trend of asymmetric parameters in three populations; B, bilateral; L, left occipital petalia; R, right occipital petalia.

Figure 4.
Cerebellar asymmetric pattern represented by Ngandong 7, La Ferrassie 1, and modern human.
Figure 4.
Cerebellar asymmetric pattern represented by Ngandong 7, La Ferrassie 1, and modern human.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of specimens used in the study.
| Populations | Number | Specimens and Source |
|---|---|---|
| Homo erectus | 11 | ZKD III, ZKD XI, Hexian (IVPP); OH 9, WT 15000, Sale, Sangiran 2, Sangiran 17, Ngandong 7, Ngandong 12, Sambungmacan 3 (AMNH) |
| Homo neanderthalensis | 4 | La Ferrassie 1, Gibraltar, Spy 1, Spy 2 (AMNH) |
| Homo sapiens | 30 | Modern Chinese (IVPP) |
Table 2.
Definition and abbreviation of measurements.
| Measurements | Abbreviation (Right/Left) | Definition |
|---|---|---|
| Cerebellar length | MLR/MLL | Length of the cerebellar major axis |
| Cerebellar height | HR/HL | Height of the cerebellum, measured from the MS point to the lowest margin of the cerebellum |
| Sagittal orientation | SagiR/SagiL | Orientation of the major axis relative to the sagittal plane, depicting how the cerebellar lobe orientated medial-laterally |
| Horizontal orientation | HoriR/HoriL | Orientation of the major axis relative to the horizontal plane, depicting how the cerebellar lobe orientated superior-inferiorly |
| Asymmetric parameters | ML.lr, H.lr, Sagi.lr, Hori.lr | Difference between the left and right side of the same measurement, calculated as (L–R) |
Table 3.
CV calculated from repeated measurements of chosen specimens.
| Specimen | Side | ML | H | Sagi | Hori |
|---|---|---|---|---|---|
| Ngandong 7 | right | 2.42 | 7.95 | 8.88 | 16.33 |
| Ngandong 7 | left | 0.73 | 2.44 | 2.01 | 22.46 |
| Gibraltar | right | 1.75 | 8.34 | 3.70 | 40.03 |
| Gibraltar | left | 3.52 | 4.15 | 4.36 | 33.63 |
| yno4f | right | 0.86 | 4.12 | 3.29 | 24.41 |
| yno4f | left | 1.03 | 1.33 | 5.49 | 19.23 |
Table 4.
Descriptive results of measurements and bootstrapped t-test.
| Population | Side | Metrics | ML | H | Sagi | Hori |
|---|---|---|---|---|---|---|
| H. erectus (n = 12) | right | mean | 48.68 | 15.18 | 40.47 | 7.68 |
| sd | 4.21 | 1.83 | 5.49 | 2.18 | ||
| cv | 8.65 | 12.05 | 13.56 | 28.42 | ||
| left | mean | 49.95 | 15.65 | 38.45 | 8.81 | |
| sd | 3.56 | 1.30 | 5.11 | 3.23 | ||
| cv | 7.13 | 8.30 | 13.29 | 36.66 | ||
| p | 0.10 | 0.45 | 0.19 | 0.26 | ||
| H. neanderthalensis (n = 4) | right | mean | 60.33 | 22.85 | 41.35 | 11.20 |
| sd | 4.28 | 3.39 | 3.36 | 4.53 | ||
| cv | 7.09 | 14.84 | 8.13 | 40.47 | ||
| left | mean | 61.65 | 20.38 | 43.15 | 10.25 | |
| sd | 4.70 | 4.41 | 4.66 | 5.76 | ||
| cv | 7.63 | 21.64 | 10.79 | 56.19 | ||
| p | 0.64 | 0.43 | 0.25 | 0.37 | ||
| H. sapiens (n = 30) | right | mean | 58.17 | 21.78 | 35.40 | 9.90 |
| sd | 2.65 | 3.02 | 3.27 | 2.82 | ||
| cv | 4.55 | 13.86 | 9.24 | 28.52 | ||
| left | mean | 59.75 | 22.01 | 36.50 | 9.83 | |
| sd | 3.33 | 3.05 | 3.46 | 2.77 | ||
| cv | 5.57 | 13.85 | 9.48 | 28.14 | ||
| p | 0.01 * | 0.64 | 0.21 | 0.91 |
* Significance level is lower than 0.05.
Table 5.
p-value of asymmetric parameters from ANCOVA.
| Asymmetric Parameter | Term | H. erectus | H. sapiens | H. neanderthalensis |
|---|---|---|---|---|
| ML.lr | ln CC | 0.03 * | 0.11 | 0.45 |
| Occipital Petalia | 0.01 * | 0.42 | ||
| H.lr | ln CC | 0.89 | 0.51 | 0.21 |
| Occipital Petalia | 0.89 | 0.01 * | ||
| Sagi.lr | ln CC | 0.42 | 0.26 | 0.49 |
| Occipital Petalia | 0.06 | 0.12 | ||
| Hori.lr | ln CC | 0.14 | 0.56 | 0.00 * |
| Occipital Petalia | 0.51 | 0.01 * |
* Significance level is lower than 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Wu, X. Asymmetries of Cerebellar Lobe in the Genus Homo. Symmetry 2021, 13, 988. https://doi.org/10.3390/sym13060988
AMA Style
Zhang Y, Wu X. Asymmetries of Cerebellar Lobe in the Genus Homo. Symmetry. 2021; 13(6):988. https://doi.org/10.3390/sym13060988
Chicago/Turabian StyleZhang, Yameng, and Xiujie Wu. 2021. "Asymmetries of Cerebellar Lobe in the Genus Homo" Symmetry 13, no. 6: 988. https://doi.org/10.3390/sym13060988
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.