
An Eco-Epidemiological Model Incorporating Harvesting Factors


Abstract
:1. Introduction
2. Mathematical Model
3. Equilibrium Points and Their Stability
- this is the trivial equilibrium point and it is exists at any time.
- The disease and predator-free equilibrium point; , always exists (it is obvious from the conditions of the parameters of the system (1)) ).
- The predator free equilibrium point whereexists in –plane provided that the following condition holds
- The infected free equilibrium point where exists in –plane if and only if the following conditions hold:
- Since so Equation (11) has a positive root if the following condition holds
4. Stability
- The eigenvalues of are , and So is the hyperbolic saddle point with locally stable manifold in the direction and with locally unstable manifold in the –direction.
- The eigenvalues of are , and therefore is locally asymptotic stable provided that and do not exist.
- The eigenvalues of satisfy the following relationswhere Hence is asymptotically stable in provided
- The eigenvalues of satisfy the following relationsandHence is asymptotically stable in provided
- Finally, the Jacobian matrix of system (1) at the interior equilibirum point is given by wherewhere Then, the characteristic equation of is given bywhereThis if and only if the following condition holdsif and only ifForhence if and only if
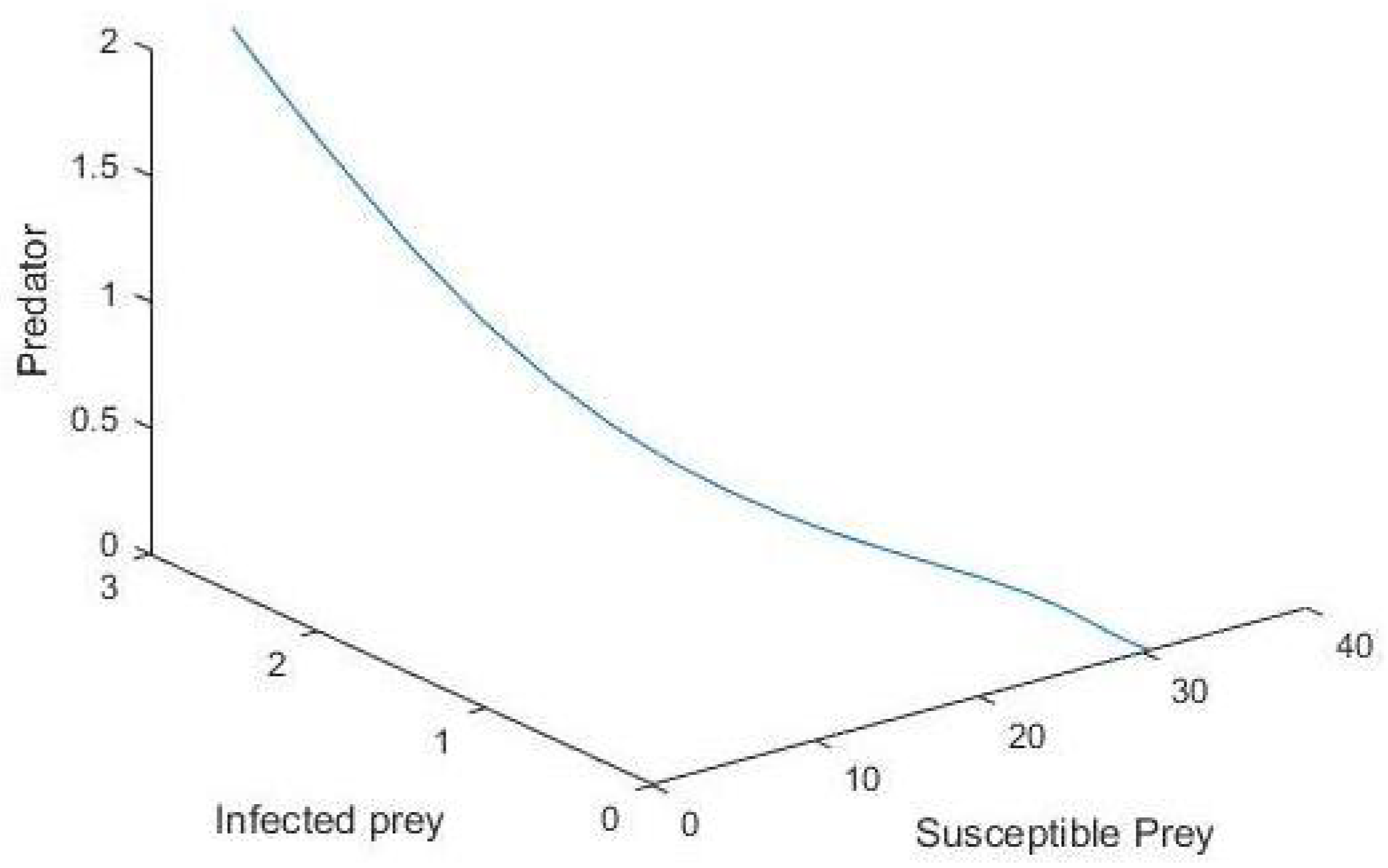
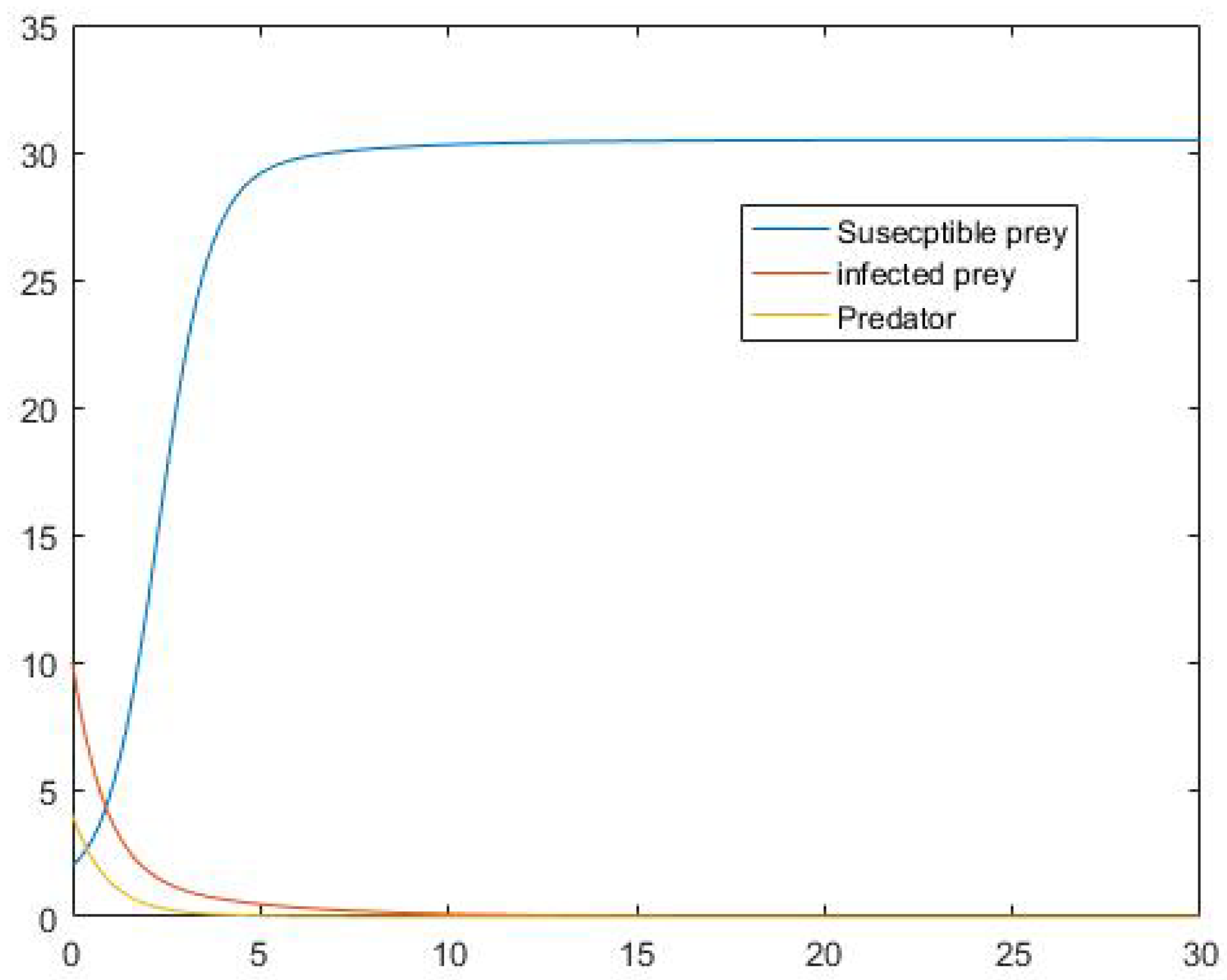
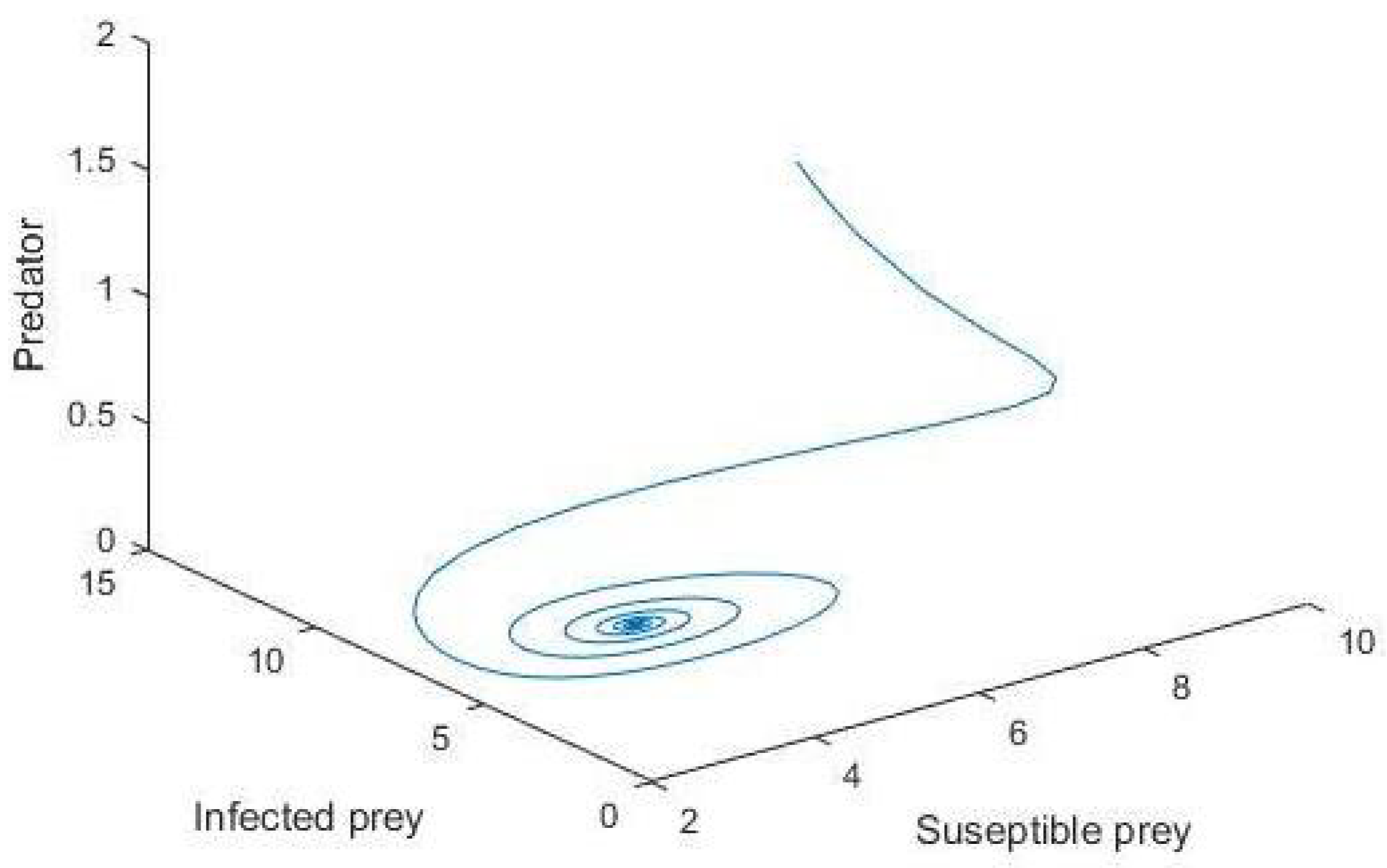
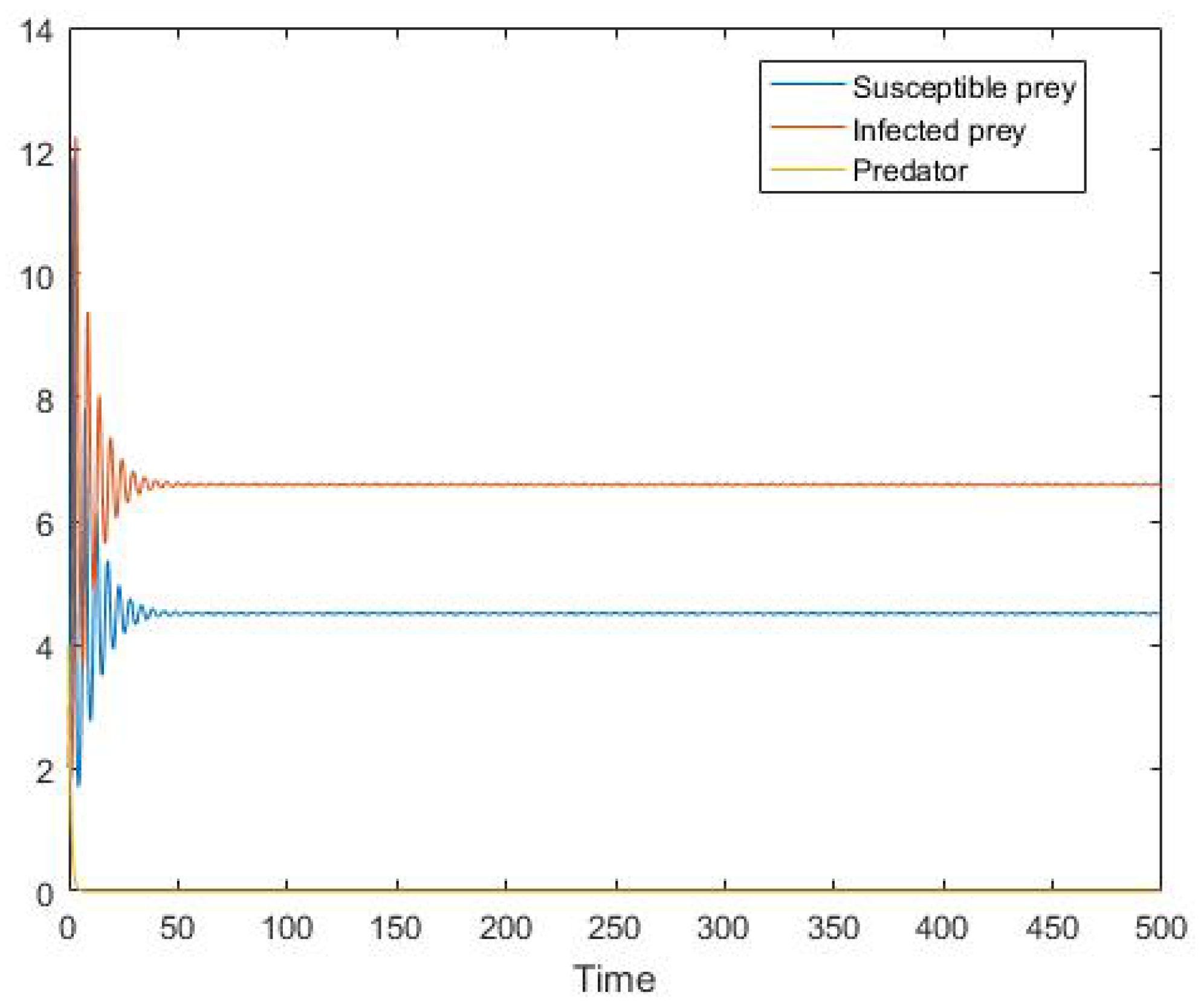
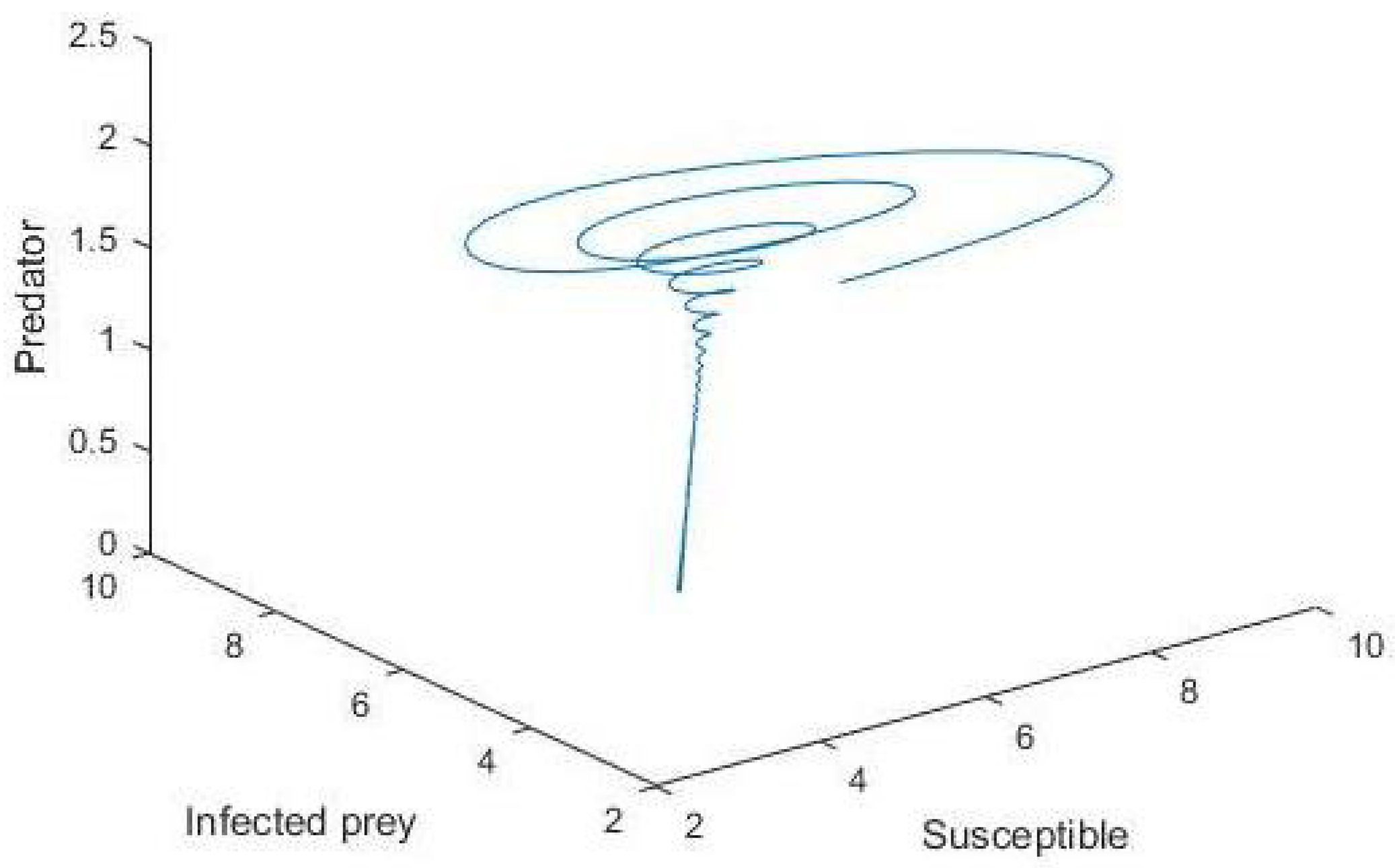
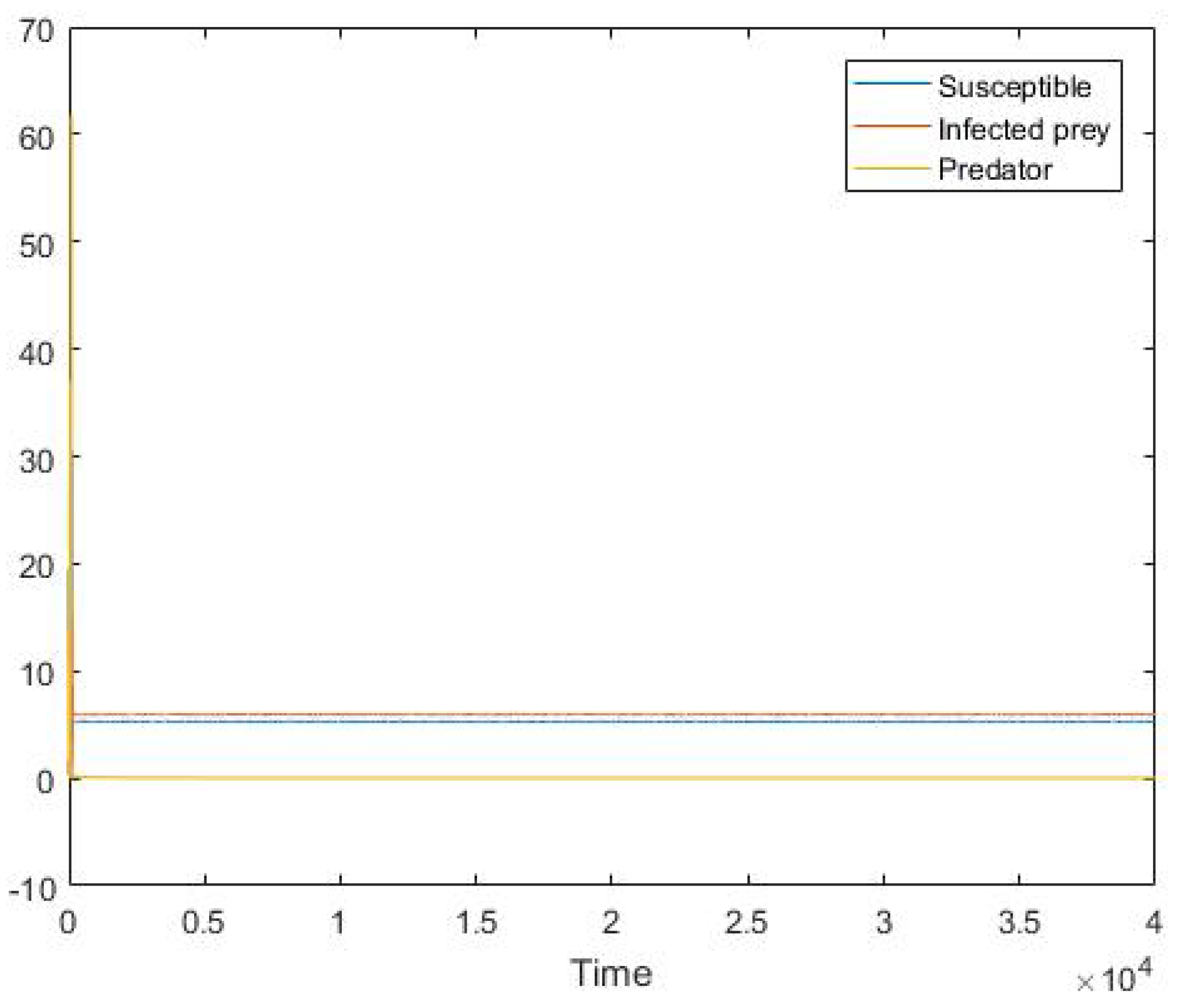
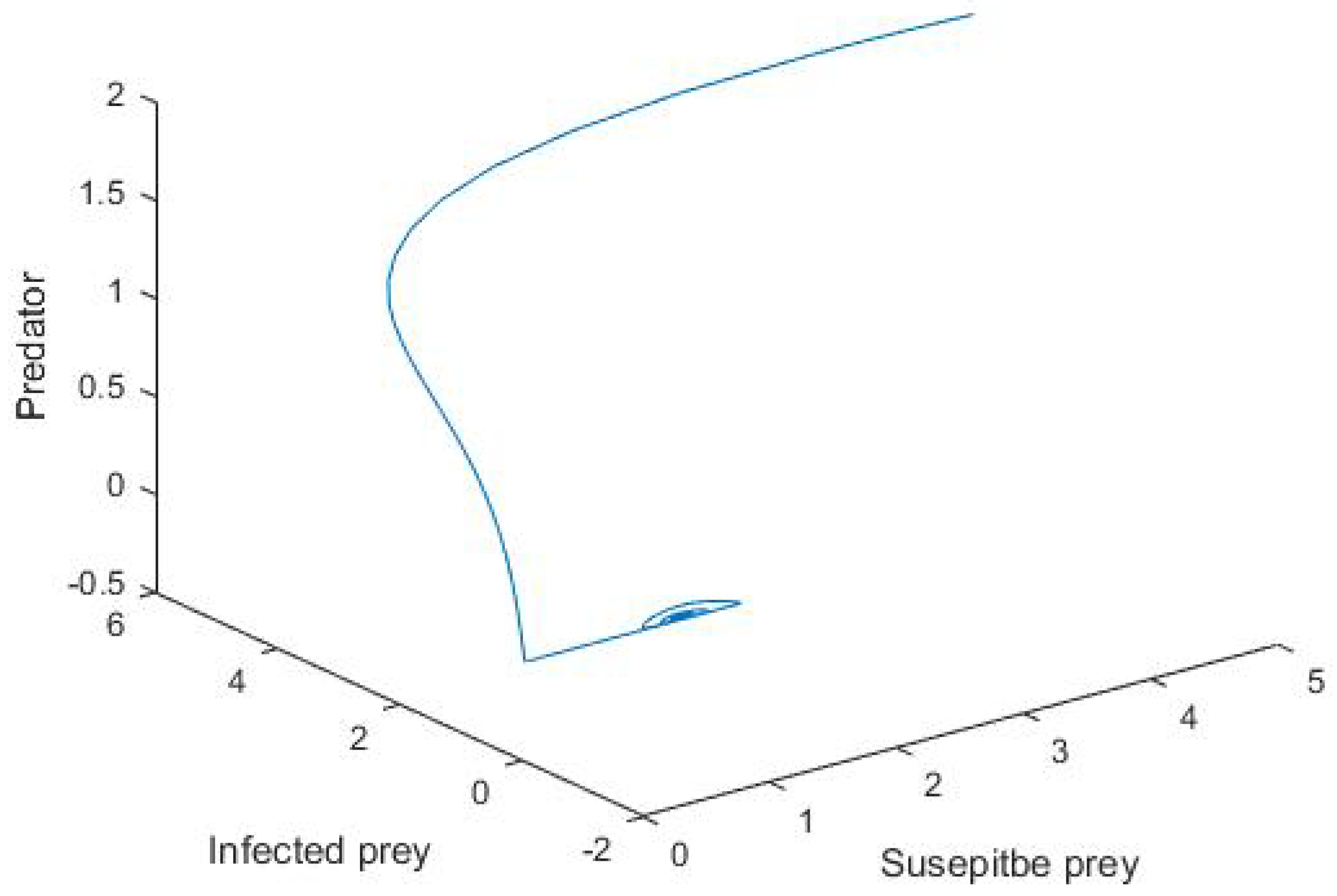
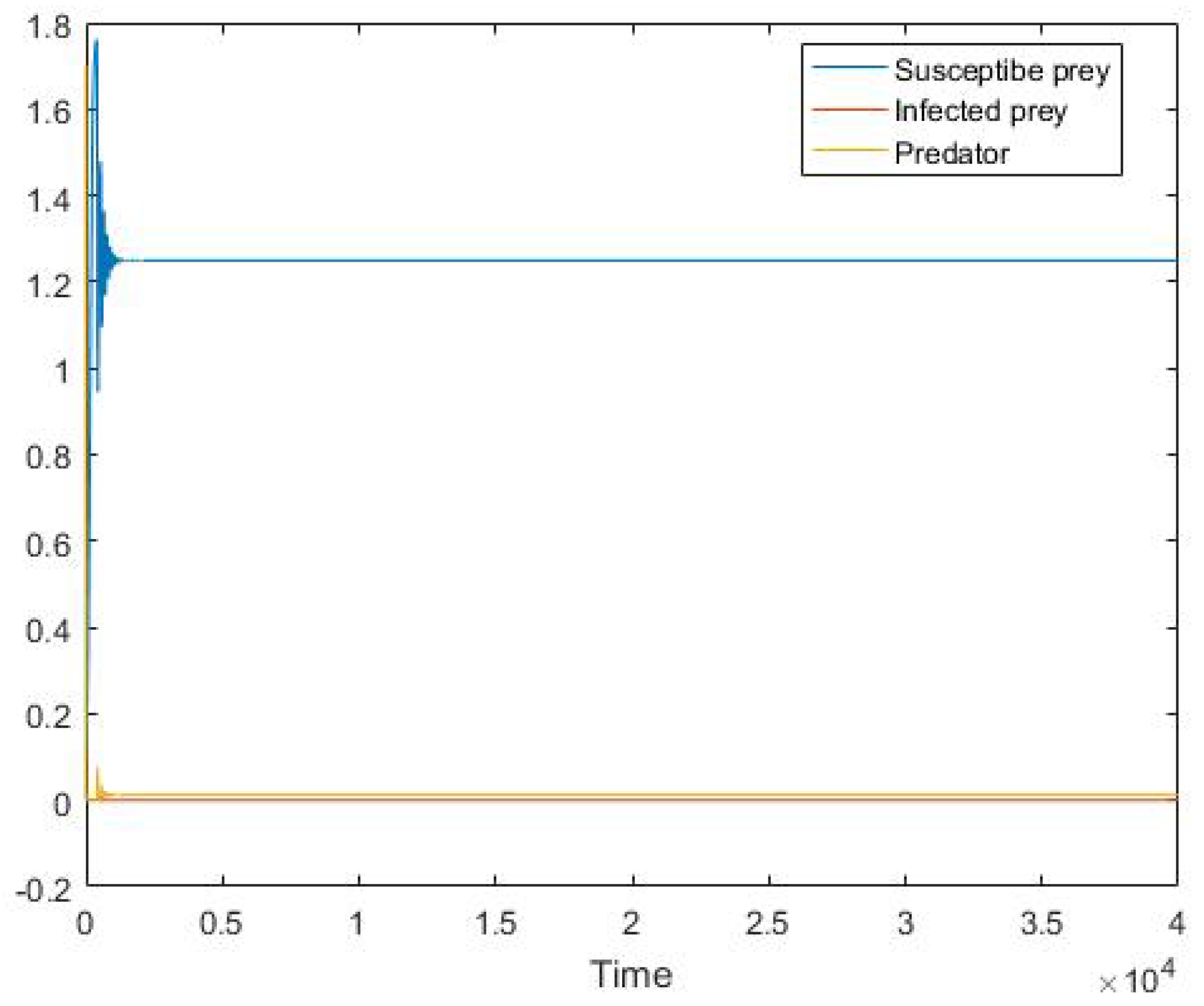
5. Numerical Simulation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nucci, M.C.; Leach, P.G.L. An integrable SIS model. J. Math. Anal. Appl. 2004, 290, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Nucci, M.C. Jacobi last multiplier and Lie symmetries: A novel application of an old relationship. J. Nonlinear Math. Phys. 2005, 12, 284–304. [Google Scholar] [CrossRef] [Green Version]
- Nucci, M.C. Using lie symmetries in epidemiology. J. Differ. Equ. 2005, 12, 87–101. [Google Scholar]
- Leach, P.G.L.; Andriopoulos, K. Application of symmetry and symmetry analyses to systems of first-order equations arising from mathematical modeling in epidemiology. Proc. Inst. Math. Nas Ukr. 2004, 50 Pt 1, 159–169. [Google Scholar]
- Nucci, M.C.; Leach, P.G.L. Singularity and symmetry analyses of mathematical models of epidemics. S. Afr. J. Sci. 2009, 105, 136–146. [Google Scholar] [CrossRef]
- Lotka, A.J. Elements of Physical Biology. Nature 1925, 116, 461. [Google Scholar] [CrossRef]
- Volterra, V. Variations and Fluctuations of the Number of Individuals in Animal Species living together. ICES J. Mar. Sci. 1928, 3, 3–51. [Google Scholar] [CrossRef]
- Kermack, W.O.; McKendrick, A.G. Contributions to the mathematical theory of epidemics-part I. Proc. R. Soc. Edinb. Sect. A 1927, 115, 700–721. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.M.; May, R.M. The invasion, persistence, and spread of infectious diseases within animal and plant communities. Philos. Trans. R. Soc. Biol. Sci. 1986, 314, 533–570. [Google Scholar] [CrossRef]
- Molla, H.; Sarwardi, S.; Sajid, M. Predator-prey dynamics with Allee effect on predator species subject to intra-specific competition and nonlinear prey refuge. J. Math. Comput. Sci. 2021, 25, 150–165. [Google Scholar] [CrossRef]
- Mustafa, A.N.; Amin, S.F. A harvested modified Leslie-gower predator-prey model with SIS-disease in predator and prey refuge. J. Duhok-Univ.- Sect. Pure Eng. Sci. 2019, 22, 174–184. [Google Scholar] [CrossRef]
- Naji, R.K.; Mustafa, A.N. The dynamics of an eco-epidemiological model with nonlinear incidence rate. J. Appl. Math. 2012, 2012, 24. [Google Scholar] [CrossRef]
- Naji, R.K.; Yaseen, R.M. Modeling and stability analysis of an eco-epidemiological Model. Iraqi J. Sci. 2013, 54, 374–385. [Google Scholar]
- Greenhalgh, D.; Khan, Q.J.A.; Al-Kharousi, F.A. Eco-epidemiological model with fatal disease in the prey. Nonlinear Anal. Real World Appl. 2020, 53, 103072. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.; Shen, X. Stability and Hopf bifurcation of a fractional-order food chain model with disease and two delays. J. Comput. Nonlinear Dynam. 2020, 15, 034501. [Google Scholar] [CrossRef]
- Biswas, S.; Saifuddin, M.; Sasmal, S.K.; Samanta, S.; Pal, N.; Ababneh, F.; Chattopadhyay, J. Optimal harvesting and complex dynamics in a delayed eco-epidemiological model with weak Allee effects. Nonlinear Dynam. 2017, 87, 1553–1573. [Google Scholar] [CrossRef]
- Biswas, S.; Saifuddin, M.; Sasmal, S.K.; Samanta, S.; Pal, N.; Ababneh, F.; Chattopadhyay, J. A delayed prey-predator system with prey subject to the strong Allee effect and disease. Nonlinear Dynam. 2016, 84, 1569–1594. [Google Scholar] [CrossRef]
- Hu, G.P.; Li, X.L. Stability and Hopf bifurcation for a delayed predator-prey model with disease in the prey. Chaos Solitons Fractals 2012, 45, 229–237. [Google Scholar] [CrossRef]
- Nandi, S.K.; Mondal, P.K.; Jana, S.; Haldar, P.; Kar, T.K. Prey-predator model with two-stage infection in prey concerning pest control. J. Nonlinear Dynam. 2015, 2015, 948728. [Google Scholar] [CrossRef] [Green Version]
- Johri, A.; Trivedi, N.; Sisodiya, A.; Sing, B.; Jain, S. Study of a prey-predator model with disease prey. Int. J. Contemp. Math. Sci. 2012, 7, 489–498. [Google Scholar]
- Sharma, S.; Samanta, G.P. Analysis of a two prey one predator system with disease in the first prey population. Int. J. Dyn. Control 2015, 3, 210–224. [Google Scholar] [CrossRef]
- Jana, S.; Guria, S.; Das, U.; Kar, T.K.; Ghorai, A. Effect of harvesting and infection on predator in a prey-predator system. Nonlinear Dynam. 2015, 81, 1–14. [Google Scholar] [CrossRef]
- Shaikh, A.A.; Das, H.; Ali, N. Study of LG-Holling type III predator-prey model with disease in predator. J. Appl. Math. Comput. 2018, 58, 235–255. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Zhang, S.H. Modelling and analysis of a delayed predator-prey model with disease in the predator. Appl. Math. Comput. 2013, 224, 372–386. [Google Scholar] [CrossRef]
- Zhang, J.S.; Sun, L. Analysis of eco-epidemiological model with epidemic in the predator. J. Biomath. 2005, 20, 157–164. [Google Scholar]
- Ko, W.; Ryu, K. Qualitative analysis of a predator–prey model with Holling type II functional response incorporating a prey refuge. J. Differ. Equ. 2006, 231, 534–550. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chen, F.; Chen, L. Qualitative analysis of a predator– prey model with Holling type II functional response incorporating a constant prey refuge. Nonlinear Anal. Real World Appl. 2010, 11, 246–252. [Google Scholar] [CrossRef]
- Selvam, A.G.M.; Jacob, S.B. Analysis of prey–predator model with holling type II functional response in discrete time. In Proceedings of the International Conference on Current Scenario in Pure and Applied Mathematics (ICCSPAM-2019), Tamil Nadu, India, 3 January 2019; Volume 2, pp. 19–44. [Google Scholar]
- Jana, S.; Kar, T.K. Modeling and analysis of a prey–predator system with disease in the prey. Chaos Solitons Fractals 2013, 47, 42–53. [Google Scholar] [CrossRef]
- Prasad, B.S.R.V.; Banerjee, M.; Srinivasu, P.D.N. Dynamics of additional food provided predator-prey system with mutually interfering predators. Math. Biosci. 2013, 246, 176–190. [Google Scholar] [CrossRef]
- Wang, X.; Zanette, L.; Zou, X. Modelling the fear effect in predator– prey interactions. J. Math. Biol. 2016, 73, 1179–1204. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.A.E.S.; Jang, B. Dynamic analysis of fractional-order predator–prey biological economic system with Holling type II functional response. Nonlinear Dyn. 2019, 96, 407–416. [Google Scholar] [CrossRef]
- Shaikh, A.A.; Das, H.; Sarwardi, S. Dynamics of an eco-epidemiological system with disease in competitive prey species. J. Appl. Math. Comput. 2020, 62, 525–545. [Google Scholar] [CrossRef]
- Bera, S.P.; Maiti, A.; Samanta, G.P. Dynamics of a food chain model with herd behaviour of the prey. Model. Earth Syst. Environ. 2016, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Mahato, P.; Mahato, S.K. Disease control prey–predator model incorporating prey refuge under fuzzy uncertainty. Model. Earth Syst. Environ. 2021, 7, 2149–2166. [Google Scholar] [CrossRef]
- Das, K.P. A study of harvesting in a predator-prey model with disease in both populations. Math. Methods Appl. Sci. 2016, 39, 2853–2870. [Google Scholar] [CrossRef]
- Kant, S.; Kumar, V. Stability analysis of predator-prey system with migrating prey and disease infection in both specise. Appl. Math. Model. 2017, 42, 509–539. [Google Scholar] [CrossRef]
- Dai, G.; Tang, M. Coexistence region and global dynamics of a harvested predator-prey system. SIAM J. Appl. Math. 1998, 58, 193–210. [Google Scholar] [CrossRef]
- Guin, L.N.; Acharya, S. Dynamic behaviour of a reaction-diffusion predator-prey model with both refuge and harvesting. Nonlinear Dynam. 2017, 88, 1501–1533. [Google Scholar] [CrossRef]
- Clark, C.W. Mathematical Bioeconomics: The Optimal Management of Renewable Resources, 2nd ed.; Wiley: New York, NY, USA, 1990. [Google Scholar]
- Bairagi, N.; Chaudhuri, S.; Chattopadhyay, J. Harvesting as a disease control measure in an ecoepidemiological system—A theoretical study. Math. Biosci. 2009, 217, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Hethcote, H.W.; Wang, W.; Han, L.; Ma, Z. A predator-prey model with infected prey. Theor. Popul. Biol. 2014, 34, 849–858. [Google Scholar] [CrossRef]
- Azar, C.; Holmberg, J.; Lindgren, K. Stability analysis of harvesting in a predator–prey model. J. Theor. Biol. 1995, 174, 13–19. [Google Scholar] [CrossRef]
- Ge, Z.; Yan, J. Hopf bifurcation of a predator-prey system with stage structure and harvesting. Nonlinear Anal. Theory Methods Appl. 2011, 74, 652–660. [Google Scholar] [CrossRef]
- Brauer, F.; Sanchez, D. Constant rate population harvesting: Equilibrium and stability. Theor. Popul. Biol. 1975, 8, 12–30. [Google Scholar] [CrossRef]
- Brauer, F.; Soudack, A.C. Stability regions and transition phenomena for harvested predator–prey systems. J. Math. Biol. 1979, 7, 319–337. [Google Scholar] [CrossRef]
- Xiao, D.; Ruan, S. Bogdanov–Takens bifurcations in predator–prey systems with constant rate harvesting. Fields Inst. Commun. 1999, 21, 493–506. [Google Scholar]
- Beddington, J.R.; Cooke, J.G. Harvesting from a prey–predator complex. Ecol. Model. 1982, 14, 155–177. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Units | Description |
|---|---|---|
| r | per day | Intrinsic growth rate constant |
| no. per unit area | Carrying capacity of the prey species in the absence of predation and harvesting | |
| m | per day | Infection rate |
| per day | The amount of handled susceptible prey in a unit time | |
| per day | Harvesting efforts for the susceptible prey | |
| per day | Maximum attack rate | |
| per day | The death rates of the infected prey | |
| per day | Harvesting efforts for the infected prey | |
| per day | The death rates of the predator | |
| g | per day | Growth rate of the predator due to predation of susceptible prey |
| h | per day | Growth rate of predator due to perdation of infected prey |
| per day | Harvesting efforts for the predator |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, K.; Mustafa, A.; Hama, M. An Eco-Epidemiological Model Incorporating Harvesting Factors. Symmetry 2021, 13, 2179. https://doi.org/10.3390/sym13112179
Hassan K, Mustafa A, Hama M. An Eco-Epidemiological Model Incorporating Harvesting Factors. Symmetry. 2021; 13(11):2179. https://doi.org/10.3390/sym13112179
Chicago/Turabian StyleHassan, Kawa, Arkan Mustafa, and Mudhafar Hama. 2021. "An Eco-Epidemiological Model Incorporating Harvesting Factors" Symmetry 13, no. 11: 2179. https://doi.org/10.3390/sym13112179