
From Neural Command to Robotic Use: The Role of Symmetry/Asymmetry in Postural and Locomotor Activities


 and
and 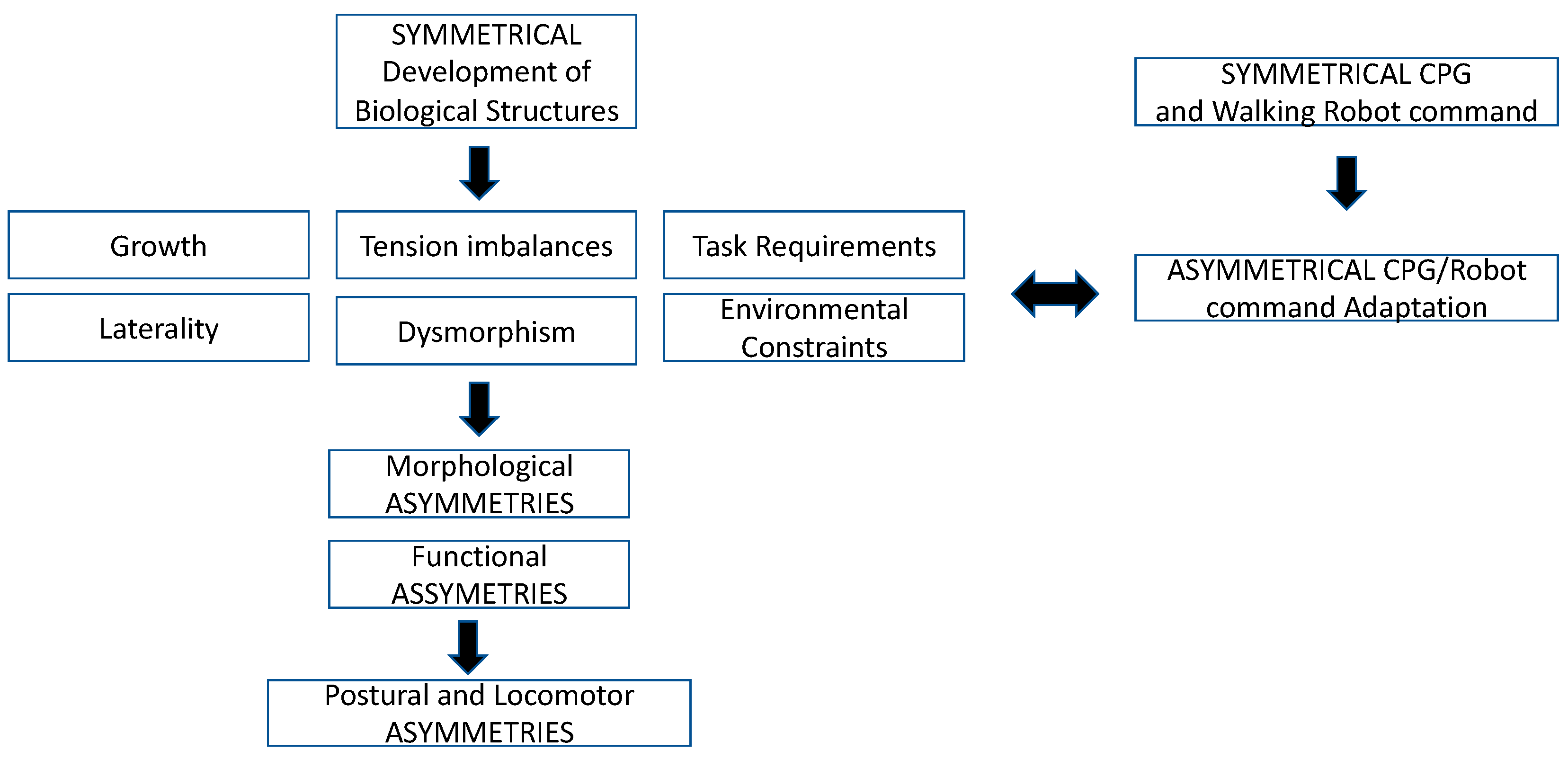{kind=link}
Abstract
:1. Introduction
2. Symmetry/Asymmetry in Human as Seen from Arts and Functional Anatomy
Symmetry and Asymmetry in Humans: From Uterine Development to Adult Life
3. A Central System for Locomotor Rhythm and Pattern Generation: Control of Symmetrical/Asymmetrical Activities by the Spinal Cord
4. Symmetry/Asymmetry from Robotic Point of View
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lindemann, U. Spatiotemporal gait analysis of older persons in clinical practice and research: Which parameters are relevant? Z. Gerontol. Geriatr. 2020, 53, 171–178. [Google Scholar] [CrossRef]
- Sadeghi, H.; Allard, P.; Prince, F.; Labelle, H. Symmetry and limb dominance in able-bodied gait: A review. Gait Posture 2000, 12, 34–45. [Google Scholar] [CrossRef]
- Sadeghi, H.; Prince, F.; Zabjek, K.; Labelle, H. Simultaneous, bilateral, and three-dimensional gait analysis of elderly people without impairments. Am. J. Phys. Med. Rehabil. 2004, 83, 112–123. [Google Scholar] [CrossRef]
- Tucker, C.; Hanley, B. Increases in speed do not change gait symmetry or variability in world-class race walkers. J. Sports Sci. 2020, 38, 2758–2764. [Google Scholar] [CrossRef] [PubMed]
- Weyl, H. Symmetry; Princeton University Press: Princeton, NJ, USA, 1952.thirty five years of its life. Int. J. Hum. Robot. 2004, 1, 157–173. [Google Scholar]
- Geismar, S. Mézières, Une Méthode, Une Femme. Le Dos Réinventé; Josette Lyon: France, 1993. [Google Scholar]
- Ingber, D.E. Cellular tensegrity: Defining new rules of biological design that govern the cytoskeleton. J. Cell Sci. 1993, 104, 613–627. [Google Scholar] [CrossRef]
- Scarr, G. Biotensegrity: The Structural Basis of Life; Handspring Publishing Limited: Pencarirland, UK, 2014. [Google Scholar]
- Scarr, G. Biotensegrity: What is the big deal? J. Bodyw. Mov. Ther. 2020, 24, 134–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, T.W. Anatomy Trains: Myofascial Meridians for Manual and Movement Therapists, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Stecco, C. Why are there so many discussions about the nomenclature of fasciae? J. Bodyw. Mov. Ther. 2014, 18, 441–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stecco, C.; Gagey, O.; Belloni, A.; Pozzuoli, A.; Porzionato, A.; Macchi, V.; Aldegheri, R.; De Caro, R.; Delmas, V. Anatomy of the deep fascia of the upper limb. Second part: Study of innervation. Morphologie. Bull. Assoc. Anat. 2007, 91, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Analyzing Fluctuating Asymmetry with Geometric Morphometrics: Concepts, Methods, and Applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef] [Green Version]
- Waitzman, K.A. The importance of positioning the near-term infant for sleep, play, and development. Newborn Infant Nurs. Rev. 2007, 7, 76–81. [Google Scholar] [CrossRef]
- Hadders-Algra, M. Variation and Variability: Key Words in Human Motor Development. Phys. Ther. 2010, 90, 1823–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aucott, S.; Donohue, P.K.; Atkins, E.; Allen, M.C. Neurodevelopmental care in the NICU. Mental Retard. Dev. Disabilities Res. Rev. 2002, 8, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Walicka-Cupry, K.; Szeliga, E.; Guzik, A.; Mrozkowiak, M.; Niewczas, M.; Ostrowski, P.; Tabaczek-Bejster, I. Evaluation of anterior-posterior spine curvatures and incidence of sagittal defects in children and adolescents practicing traditional karate. BioMed Res. Int. 2019, 2019, 9. [Google Scholar]
- Blanchet, M.; Marchand, D.; Cadoret, G. Postural control adjustments during progressive inclination of the support surface in children. Med. Eng. Phys. 2012, 34, 1019–1023. [Google Scholar] [CrossRef]
- Hadders-Algra, M.; Brogren, E.; Forssberg, H. Postural adjustments during sitting at preschool age: Presence of a transient toddling phase. Dev. Med. Child Neurol. 1998, 40, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Berger, W.; Discher, M.; Trippel, M.; Ibrahim, K.I.; Dietz, V. Developmental aspects of stance regulation, compensation and adaptation. Exp. Brain Res. 1992, 90, 610–619. [Google Scholar] [CrossRef]
- Kirshenbaum, N.; Riach, C.; Starkes, J. Non-linear development of postural control and strategy use in young children: A longitudinal study. Exp. Brain Res. 2001, 140, 420–431. [Google Scholar] [CrossRef]
- Changeux, J.P. Variation and selection in neural function. Trends Neurosci. 1997, 20, 291–293. [Google Scholar]
- Diamond, A. The interplay of biology and the environment broadly defined. Dev. Psychol. 2009, 45, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ismail, F.Y.; Fatemi, A.; Johnston, M.V. Cerebral plasticity: Windows of opportunity in the developing brain. Eur. J. Paediatr. Neurol. 2017, 21, 23–48. [Google Scholar] [CrossRef]
- Paillard, T. Plasticity of the postural function to sport and/or motor experience. Neurosci. Biobehav. Rev. 2017, 72, 129–152. [Google Scholar] [CrossRef]
- Shumway-Cook, A.; Woollacott, M.H. The growth of stability: Postural control from a development perspective. J. Mot. Behav. 1985, 17, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Hay, L.; Redon, C. Feedforward versus feedback control in children and adults subjected to a postural disturbance. Exp. Brain Res. 1999, 125, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Sigmundsson, H.; Whiting, H.T.; Loftesnes, J.M. Development of proprioceptive sensitivity. Exp. Brain Res. 2000, 135, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Riach, C.L.; Starkes, J.L. Stability limits of quiet standing postural control in children and adults. Gait Posture 1993, 1, 105–111. [Google Scholar] [CrossRef]
- Figura, F.; Cama, G.; Capranica, L.; Guidetti, L.; Pulejo, C. Assessment of static balance in children. J. Sports Med. Phys. Fit. 1991, 31, 235–242. [Google Scholar]
- Riach, C.L.; Starkes, J.L. Velocity of centre of pressure excursions as an indicator of postural control systems in children. Gait Posture 1994, 2, 167–172. [Google Scholar] [CrossRef]
- Rival, C.; Ceyte, H.; Olivier, I. Developmental changes of static standing balance in children. Neurosci. Lett. 2005, 376, 133–136. [Google Scholar] [CrossRef]
- Verbecque, E.; Vereeck, L.; Hallemans, A. Postural sway in children: A literature review. Gait Posture 2016, 49, 402–410. [Google Scholar] [CrossRef]
- Assaiante, C. Action and representation of action during childhood and adolescence: A functional approach. Clin. Neurophysiol. 2012, 42, 43–51. [Google Scholar] [CrossRef]
- Sabharwal, S.; Zhao, C. The Hip-Knee-Ankle Angle in Children: Reference Values Based on a Full-Length Standing Radiograph. J. Bone Joint Surg. Am. 2009, 91, 2461–2468. [Google Scholar] [CrossRef]
- Kernell, D. The final common pathway in postural control developmental perspective. Neurosci. Biobehav. Rev. 1998, 22, 479–484. [Google Scholar] [CrossRef]
- Samsom, J.; DeGroot, L.; Bezemer, P.; Lafeber, H.; Fetter, W. Muscle power development during the first year of life predicts neuromotor behaviour at 7 years in pretermborn. Early Hum. Dev. 2002, 68, 103–118. [Google Scholar] [CrossRef]
- Parfitt, A.M.; Travers, R.; Rauch, F.; Glorieux, F.H. Structural and cellular changes during bone growth in healthy children. Bone 2000, 27, 487–494. [Google Scholar] [CrossRef]
- Kannus, P.; Haapasalo, H.; Sankelo, M.; Sievanen, H.; Pasanen, M.; Heinonen, A.; Oja, P.; Vuori, I. Effect of Starting Age of Physical Activity on Bone Mass in the Dominant Arm of Tennis and Squash Players. Ann. Int. Med. 1995, 123, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Fagard, J. Droitiers/Gauchers: Des asymmetries dans tous les sens. De Boeck superieur Solal: Marseille, France, 2004. [Google Scholar]
- Brown, E.R.; Taylor, P.J. Handness, footness and eyeness. Percept. Mot. Skills 1988, 66, 183–186. [Google Scholar] [CrossRef]
- Gilberta, G.; Nassogneb, M. Mise au point face à l’acquisition d’une latéralité précoce de l’enfant. Focus on early acquisition of lateralization in children. Kinesither. Rev. 2020, 20, 25–28. [Google Scholar]
- Massion, J.; Popov, K.; Fabre, J.C.; Rage, P.; Gurfinkel, V. Is the erect posture in microgravity based on the control of trunk orientation or center of mass position? Exp. Brain Res. 1997, 114, 384–389. [Google Scholar] [CrossRef]
- Assaiante, C.; Barlaam, F.; Cignetti, F.; Vaugoyeau, M. Body schema building during childhood and adolescence: A neurosensory approach. Neurophysiol. Clin. 2014, 44, 3–12. [Google Scholar] [CrossRef]
- Naito, E.; Morita, T.; Amemiyaa, K. Body representations in the human brain revealed by kinesthetic illusions and their essential contributions to motor control and corporeal awareness. Neurosci. Res. 2016, 104, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.T. The intrinsic factors in the act of progression in the mammal. Proc. R. Soc. Lond. 1911, 84, 309–319. [Google Scholar]
- Brown, G.T. On the nature of the fundamental activity of the nervous centres; together with an analysis of the conditioning of rhythmic activity in progression, and a theory of the evolution of function in the nervous system. J. Physiol. 1914, 48, 18–46. [Google Scholar] [CrossRef]
- Grillner, S.; Zangger, P. Locomotor movements generated by the deafferented spinal cord. Acta Physiol. Scand 1974, 91, 38A–39A. [Google Scholar]
- Grillner, S.; Zangger, P. On the central generation of locomotion in the low spinal cat. Exp. Brain Res. 1979, 34, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E.; Jukes, M.; Lund, S.; Lundberg, A. The effect of DOPA on the spinal cord. 5. Reciprocal organization of pathways transmitting excitatory action to alpha motoneurones of flexors and extensors. Acta Physiol. Scand. 1967, 70, 369–388. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E.; Jukes, M.; Lund, S.; Lundberg, A. The effect of DOPA on the spinal cord. 6. Half-centre organization of interneurones transmitting effects from the flexor reflex afferents. Acta Physic. Scand. 1967, 70, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Guertin, P. Central pattern generator for locomotion: Anatomical, physiological, and pathophysiological considerations. Front. Neurol. 2013, 3, 183. [Google Scholar] [CrossRef] [Green Version]
- Rowland, R. Principle of Human Locomotion; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2020. [Google Scholar]
- Steuer, I.; Guertin, P. Central pattern generators in the brainstem and spinal cord: An overview of basic principles, similarities, and differences. Rev. Neurosci. 2019, 30, 107–164. [Google Scholar] [CrossRef]
- Tan, U. Two families with quadrupedalism, mental retardation, no speech, and infantile hypotonia (Uner Tan Syndrome Type-II); a novel theory for the evolutionary emergence of human bipedalism. Front Neurosci. 2014, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Guertin, P.; Dubuc, R. Effects of stimulating the reticular formation during fictive locomotion in lampreys. Brain Res. 1996, 753, 328–334. [Google Scholar] [CrossRef]
- Wannier, T.; Deliagina, G.; Orlovsky, G.; Grillner, S. Differential effects of the reticular system on locomotion in lamprey. J. Neurophysiol. 1998, 80, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Shik, M.L.; Severin, F.V.; Orlovsky, G.N. Control of walking and running by means of electrical stimulation of the midbrain. Biofizika 1966, 11, 659–666. [Google Scholar]
- Shik, M.L.; Severin, F.V.; Orlovsky, G.N. Control of walking and running by means of electrical stimulation of the mesencephalon. Electroencephalogr. Clin. Neurophysiol. 1969, 26, 549. [Google Scholar] [PubMed]
- Forssberg, H.; Grillner, S.; Halbertsma, J.; Rossignol, S. The locomotion of the low spinal cat. II. Interlimb coordination. Acta Physiol. Scan. 1980, 108, 283–295. [Google Scholar] [CrossRef]
- Frigon, A.; Desrochers, É.; Thibaudier, Y.; Hurteau, M.; Dambreville, C. Left-right coordination from simple to extreme conditions during split-belt locomotion in the chronic spinal adult cat. J. Physiol. 2017, 585, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Forssberg, H.; Grillner, S.; Rossignol, S. Phase-dependent reflex reversal during walking in chronic spinal cats. Brain Res. 1975, 85, 103–107. [Google Scholar] [CrossRef]
- Guertin, P.; Angel, M.; Perreault, M.-C.; McCrea, D. Ankle extensor group I afferents excite extensors throughout the hindlimb during fictive locomotion in the cat. J. Physiol. 1995, 487, 197–209. [Google Scholar] [CrossRef]
- Perreault, M.-C.; Angel, M.; Guertin, P.; Mccrea, D. Effects of stimulation of hindlimb flexor group II afferents during fictive locomotion in the cat. J. Physiol. 1995, 487, 211–220. [Google Scholar] [CrossRef]
- Griffin, M.; Olney, S.; McBride, I. Role of symmetry in gait performance of stroke subjects with hemiplegia. Gait Posture 1995, 3, 132–142. [Google Scholar] [CrossRef]
- Kagawa, T.; Nomura, T.; Kondo, S. Interlimb Parallel-link Powered Orthosis (IPPO): Compact Wearable, Robot with Lateral Weight Bearing Mechanisms for Gait Assistance. IEEE Trans. Med. Robot. Bionics 2020, 2, 300–308. [Google Scholar] [CrossRef]
- Meng, S.; Jin, S.; Li, J.; Hashimoto, K.; Guo, S.; Dai, S. The Analysis of HumanWalking Stability Using ZMP in Sagittal Plane. In Proceedings of the IEEE Intertional Conference on Cybernetics and Intelligent Systems, Ningbo, China, 19–21 November 2017; pp. 296–501. [Google Scholar]
- Ng, K.; Mehdizadeh, S.; Laboni, A.; Mansfield, A.; Flint, A.; Taati, B. Measuring gait variables using computer vision to assess mobility and fall risk in older adults with dementia. IEEE J. Transl. Eng. Health Med. 2020, 8, 2100609. [Google Scholar] [CrossRef]
- Rodano, R.; Santambrogio, G. Quantitative comparison of locomotor performance in different race walkers. In Proceedings of the 5th International Symposium on Biomechanics in Sports. Conference Proceeding Archive, Athens, Greece, 1987; pp. 122–134. [Google Scholar]
- Chodera, J.D.; Levell, R.W. Footprint Patterns during Walking. In Perspectives in Biomedical Engineering: Proceedings of a Symposium Organised in Association with the Biological Engineering Society and Held in the University of Strathclyde, Glasgow, June 1972; Kenedi, R.M., Ed.; Palgrave Macmillan: London, UK, 1973; pp. 81–90. [Google Scholar]
- Gundersen, L.; Valle, D.; Barr, A.; Danoff, J.; Stanhope, S.; Snyder-Mackler, L. Bilateral analysis of the knee and ankle during gait: An examination of the relationship between lateral dominance and symmetry. Phys. Ther. 1989, 69, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Reisman, D.; Wityk, R.; Silver, K.; Bastian, A. Locomotor adaptation on a split-belt treadmill can improve walking symmetry post-stroke. Brain 2007, 130, 1861–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forczek, W.; Staszkiewicz, R. An evaluation of symmetry in the lower limb joints during the able-bodied gait of women and men. J. Hum. Kinetics 2012, 35, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Hannah, R.; Morrison, J.; Chapman, A. Kinematic symmetry of the lower limbs. Arch. Phys. Med. Rehabil. 1984, 65, 155–158. [Google Scholar] [PubMed]
- Karamanidis, K.; Arampatzis, A.; Bruggemann, G. Symmetry and reproducibility of kinematic parameters during various running techniques. Med. Sci. Sports Exerc. 2003, 35, 1009–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, H. Microcomputer-based low-cost method for measurement of spatial and temporal parameters of gait. J. Biomed. Eng. 1987, 9, 115–120. [Google Scholar] [CrossRef]
- Hamill, J.; Bates, B.; Knutzen, K. Ground reaction force symmetry during walking and running. Res. Q. Exerc. Sport 1984, 55, 289–293. [Google Scholar] [CrossRef]
- Herzog, W.; Nigg, B.; Read, L.; Olsson, E. Asymmetries in ground reaction force patterns in normal human gait. Med. Sci. Sports Exerc. 1989, 21, 110–114. [Google Scholar] [CrossRef]
- Hsiao-Wecksler, E.; Polk, J.; Rosengren, K.; Sosnoff, J.; Hong, S. A review of new analytic techniques for quantifying symmetry in locomotion. Symmetry 2010, 2, 1135–1155. [Google Scholar] [CrossRef] [Green Version]
- Arsenault, A.; Winter, D.; Marteniuk, R. Is there a ‘normal’ profile of EMG activity in gait? Med. Biol. Eng. Comput. 1986, 24, 337–343. [Google Scholar] [CrossRef]
- Õunpuu, S.; Winter, D. Bilateral electromyographical analysis of the lower limbs during walking in normal adults. Electroencephalogr. Clin. Neurophysiol. 1989, 72, 429–438. [Google Scholar] [CrossRef]
- Balakrishan, S.; Thornton-Trump, A. Integral parameters in human locomotion. In Proceedings of the Process 2nd Biannual Conference of the Canadian Society for Biomechanics, Kingston, ON, Canada, 31 August–3 September 1982; pp. 12–13. [Google Scholar]
- Damholt, V.; Termansen, N. Asymmetry of plantar flexion strength in the foot. Acta Orthop. 1978, 49, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, C. Are joint torques the Holy Grail of human gait analysis? Hum. Mov. Sci. 1996, 15, 423–443. [Google Scholar] [CrossRef]
- Crowe, A.; Schiereck, P.; De Boer, R.; Keessen, W. Characterization of human gait by means of body center of mass oscillations derived from ground reaction forces. IEEE Trans. Biomed. Eng. 1995, 42, 293–303. [Google Scholar] [CrossRef]
- Giakas, G.; Baltzopoulos, V. Time and frequency domain analysis of ground reaction forces during walking: An investigation of variability and symmetry. Gait Posture 1997, 5, 189–197. [Google Scholar] [CrossRef]
- Rice, J.; Seeley, M. An investigation of lower-extremity functional asymmetry for non-preferred able-bodied walking speeds. Int. J. Exerc. Sci. 2010, 3, 182–188. [Google Scholar] [PubMed]
- Seeley, M.; Umberger, B.; Shapiro, R. A test of the functional asymmetry hypothesis in walking. Gait Posture 2008, 28, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Carpes, F.; Mota, C.; Faria, I. On the bilateral asymmetry during running and cycling: A review considering leg preference. Phys. Ther. Sport 2010, 11, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.; Figueroa, A.; Fernhall, B.; Kanaley, J.A. Energy expenditure of walking and running: Comparison with prediction equations. Med. Sci. Sports Exerc. 2004, 36, 2128–2134. [Google Scholar] [CrossRef] [Green Version]
- Sardian, P.; Bessonnet, G. Forces acting on a biped robot. Center of pressure-zero moment point. Trans. Syst. Man Cybern. Part A Syst. Hum. 2004, 34, 630–637. [Google Scholar] [CrossRef]
- Vukobratovic, M.; Borovac, B. Zero-moment point–thirty five years of its life. Int. J. Hum. Robot. 2004, 1, 157–173. [Google Scholar] [CrossRef]
- Vukobratovic, M.; Frank, A.; Juricic, D. On the stability of biped locomotion. IEEE Trans. Biomed. Eng. 1970, BME-17, 25–36. [Google Scholar] [CrossRef]
- Wada, Y.; Kanekoa, Y.; Nakano, E.; Osu, R.; Kawato, M. Quantitative examinations for multi joint arm trajectory planning using a robust calculation algorithm of the minimum commanded torque change trajectory. Neural Netw. 2001, 14, 381–393. [Google Scholar] [CrossRef]
- Li, Q.; Takanishi, A.; Kato, I. Learning control for a biped walking robot with a trunk. In Proceedings of the Process of 1993 IEEE-RSJ International Conference on Intelligent Robots and Systems IROS 93, Yokohama, Japan, 26–30 July 1993. [Google Scholar]
- Fujimoto, Y.; Kawamura, A. Three dimensional digital simulation and autonomous walking control for eight-axis biped robot. In Proceedings of the IEEE Intenational Conference on Robotics and Automation, Nagoya, Japan, 21–27 May 1995; pp. 2877–2884. [Google Scholar]
- Hirai, K.; Hirose, M.; Haikawa, Y.; Takenaka, T. The development of Honda humanoid robot. In Proceedings of the IEEE International Conference on Robotics and Automation (ICRA), Leuven, Belgium, 16–20 May 1998; pp. 1321–1326. [Google Scholar]
- Takanishi, A.; Ishida, M.; Yamazaki, Y.; Kato, I. The realization of dynamic walking by the biped walking robot wl-10rd. In Proceedings of the IEEE International Conference on Robotics and Automation, St. Louis, MO, USA, 25 March 1985; pp. 459–466. [Google Scholar]
- Nishiwaki, K.; Kagami, S.; Kuniyoshi, Y.; Inaba, M.; Inoue, H. Online generation of humanoid walking motion based on a fast generation method of motion pattern that follows desired ZMP. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems, Lausanne, Switzerland, 30 September–4 October 2002; pp. 2684–2689. [Google Scholar]
- Huang, Q.; Peng, Z.; Zhang, W.; Zhang, L.; Li, K. Design of humanoid complicated dynamic motion based on human motion capture. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems, Edmonton, AB, Canada, 2–6 August 2005. [Google Scholar]
- Gorce, P. Dynamic postural control method for biped in unknown environment. IEEE Trans. Syst. Man Cybern. 1999, 29, 616–626. [Google Scholar] [CrossRef]
- Gorce, P.; Guihard, M. On dynamics control of pneumatic bipeds. J. Robotic Syst. 1998, 15, 421–433. [Google Scholar] [CrossRef]
- Gorce, P.; Hafi, E.l.; Coronado, J.L. Dynamic control of walking cycle with initiation process for humanoid robot. J. Intell. Robotics Syst. 2001, 31, 321–337. [Google Scholar] [CrossRef]
- Gorce, P.; Vanel, O.; Ribreau, C. Equilibrium study of human robot. In Proceedings of the IEEE International Conference on Systems Man and Cybernetics, Vancouver, BC, Canada, 22–25 October 1995; pp. 1309–1314. [Google Scholar]
- Hirukawa, H.; Hattori, S.; Kajita, S.; Harada, K.; Kaneko, K.; Kanehiro, F.; Morisawa, M.; Nakaoka, S. A pattern generator of humanoid robots walking on a rough terrain. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007; pp. 2181–2187. [Google Scholar]
- Kim, S.; Murakami, T. An Approach to Human Walking Analysis Based on Balance, Symmetry and Stability Using COG, ZMP and CP. Appl. Sci. 2020, 10, 7307. [Google Scholar] [CrossRef]
- Goswami, A. Postural Stability of Biped Robots and the Foot-Rotation Indicator (FRI) Point. Int. J. Robot. Res. 1999, 18, 523–533. [Google Scholar] [CrossRef]
- Ortiz, A.; Ibarra, J. Walk stability control for position-controlled servo actuated humanoid robot. In Proceedings of the IEEE International Conference on Electrical Engineering, Computing Science and Automatic Control, Mexico City, Mexico, 11–13 September 2019; pp. 1–6. [Google Scholar]
- Raoufi, M.; Edrisi, M. Designing a Fractional Order Back-Stepping Controller Based on GPI Observer for a 3D Biped Robot. In Proceedings of the IEEE International Conference on Robotics and Mechatronics, Tehran, Iran, 20–21 November 2019; pp. 211–216. [Google Scholar]
- Soliman, A.; Sendur, P.; Ugurlu, B. 3-D DynamicWalking Trajectory Generation for a Bipedal Exoskeleton with Underactuated Legs: A Proof of Concept. In Proceedings of the IEEE International Conference on Rehabilitation Robotics, Toronto, ON, Canada, 24–28 June 2019; pp. 599–604. [Google Scholar]
- Tesio, L.; Rota, V. The motion of body center of mass during walking: A review oriented to clinical applications. Front. Neurol. 2019, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanchet, M.; Guertin, P.; Pilon, F.; Gorce, P.; Prince, F. From Neural Command to Robotic Use: The Role of Symmetry/Asymmetry in Postural and Locomotor Activities. Symmetry 2021, 13, 1773. https://doi.org/10.3390/sym13101773
Blanchet M, Guertin P, Pilon F, Gorce P, Prince F. From Neural Command to Robotic Use: The Role of Symmetry/Asymmetry in Postural and Locomotor Activities. Symmetry. 2021; 13(10):1773. https://doi.org/10.3390/sym13101773
Chicago/Turabian StyleBlanchet, Mariève, Pierre Guertin, Francine Pilon, Philippe Gorce, and François Prince. 2021. "From Neural Command to Robotic Use: The Role of Symmetry/Asymmetry in Postural and Locomotor Activities" Symmetry 13, no. 10: 1773. https://doi.org/10.3390/sym13101773