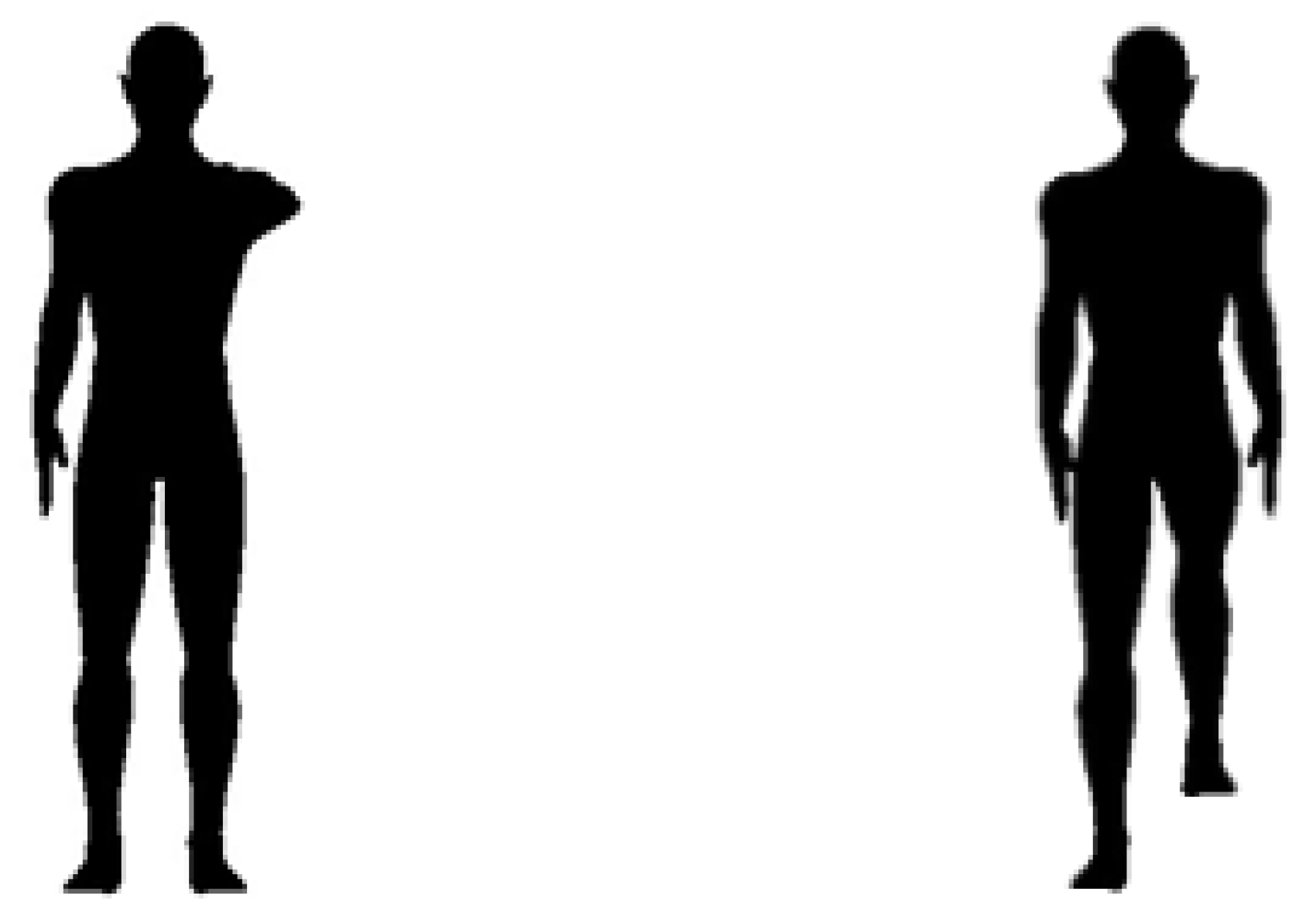
The Bias toward the Right Side of Others Is Stronger for Hands than for Feet

 , , ,
, , , 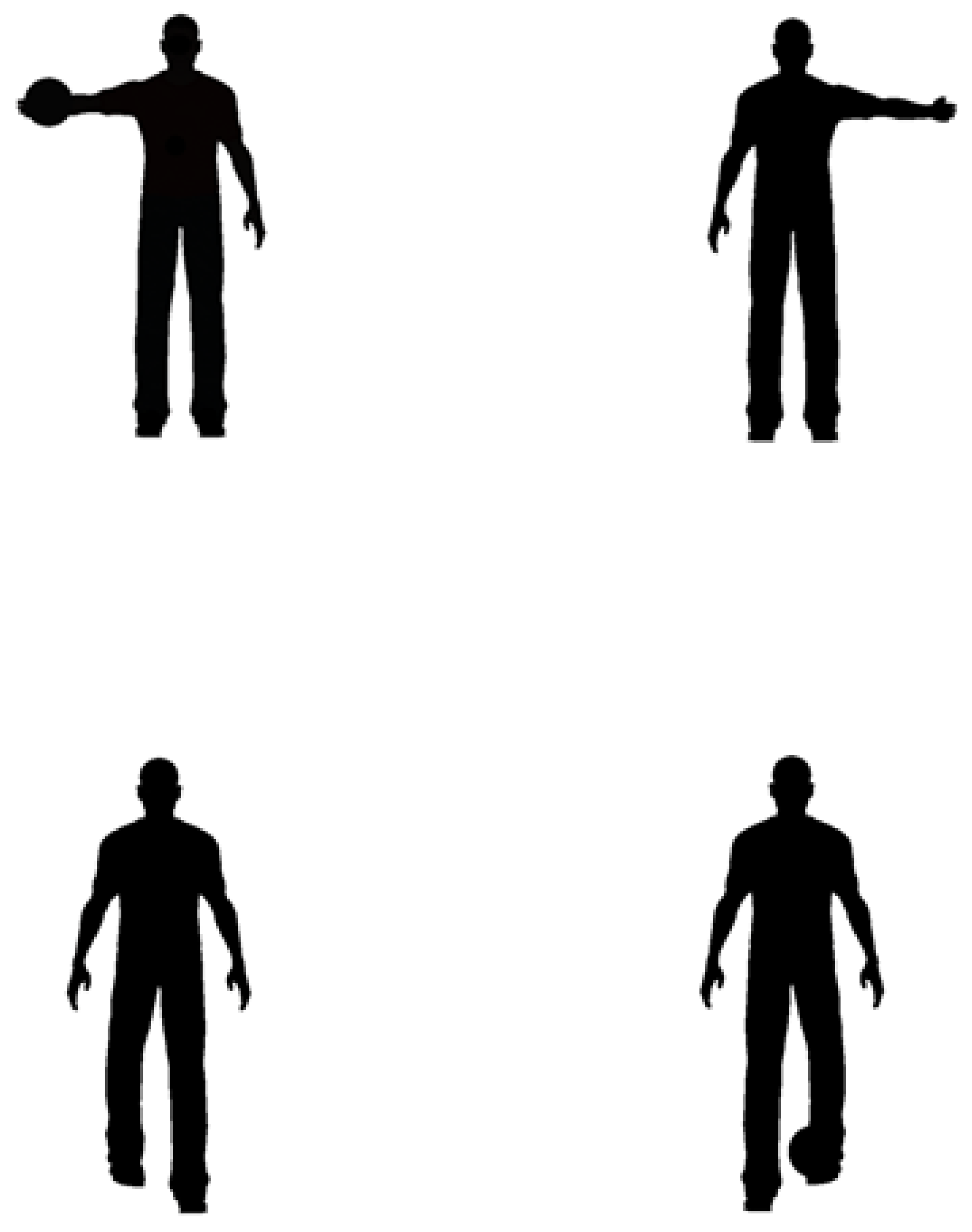{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experiment 1
2.1. Materials and Methods
2.1.1. Participants
2.1.2. Stimuli
2.1.3. Procedure
2.1.4. Data Analysis
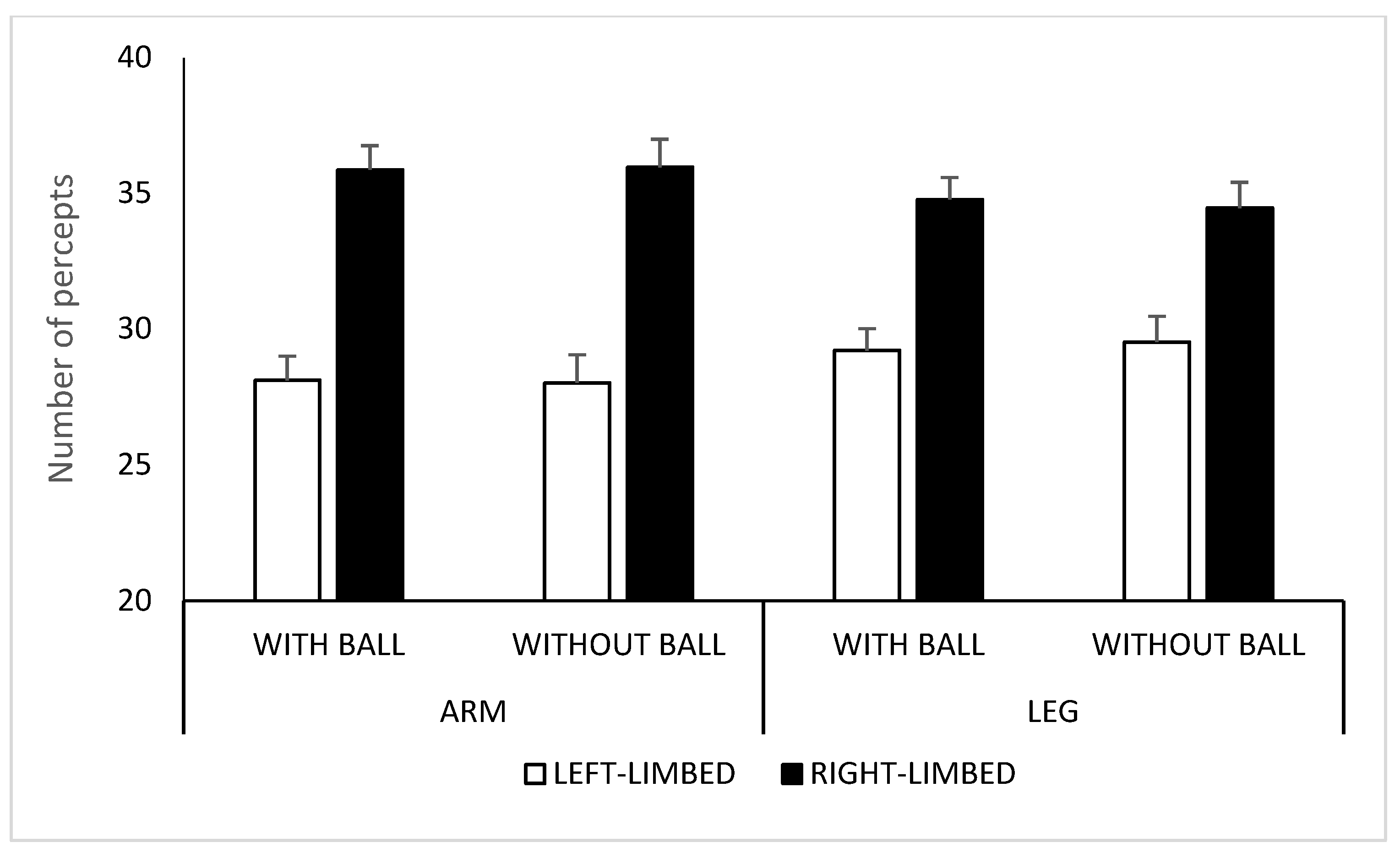
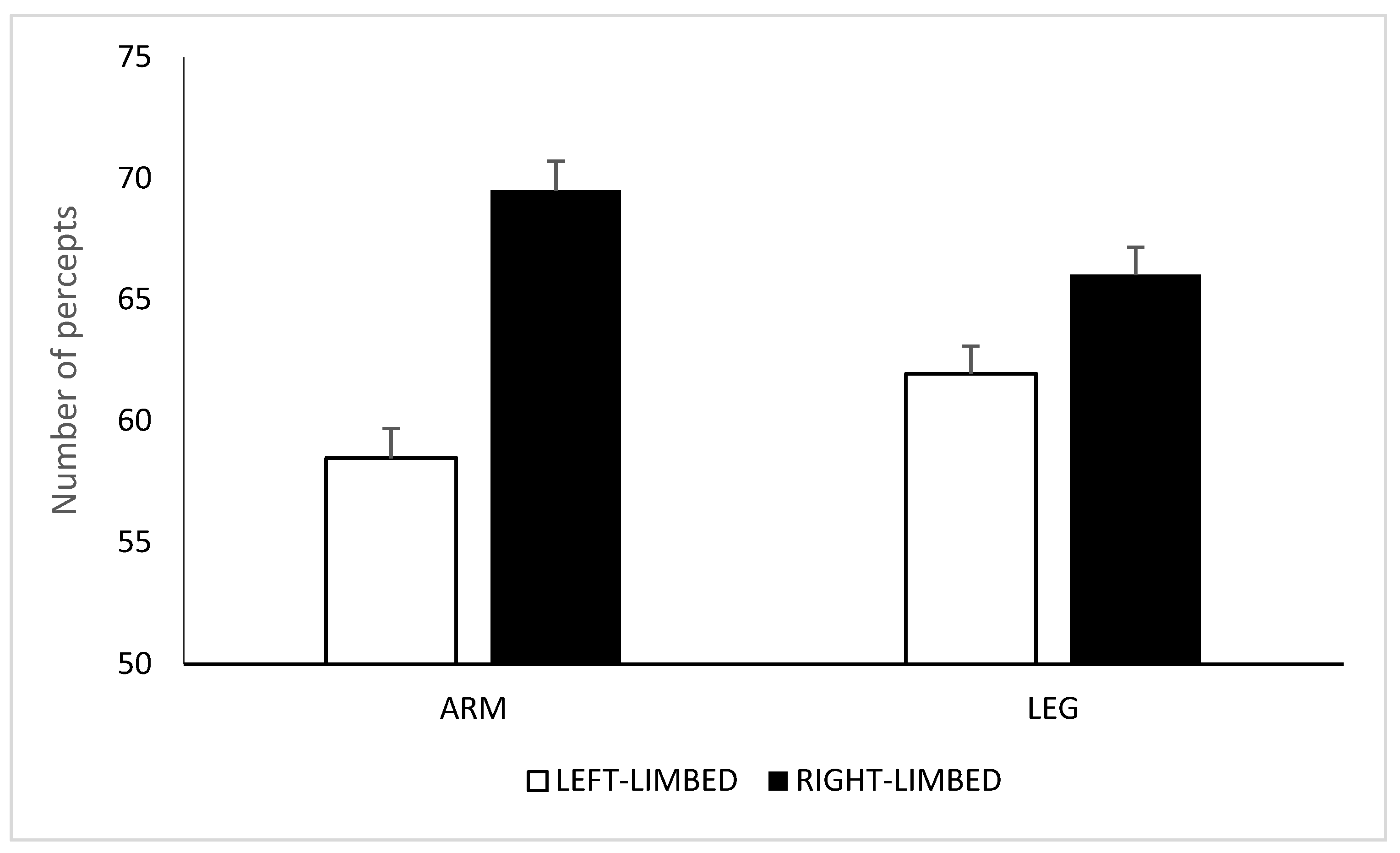
2.2. Results and Discussion
3. Experiment 2
3.1. Materials and Methods
3.1.1. Participants
3.1.2. Stimuli
3.1.3. Procedure
3.1.4. Data Analysis
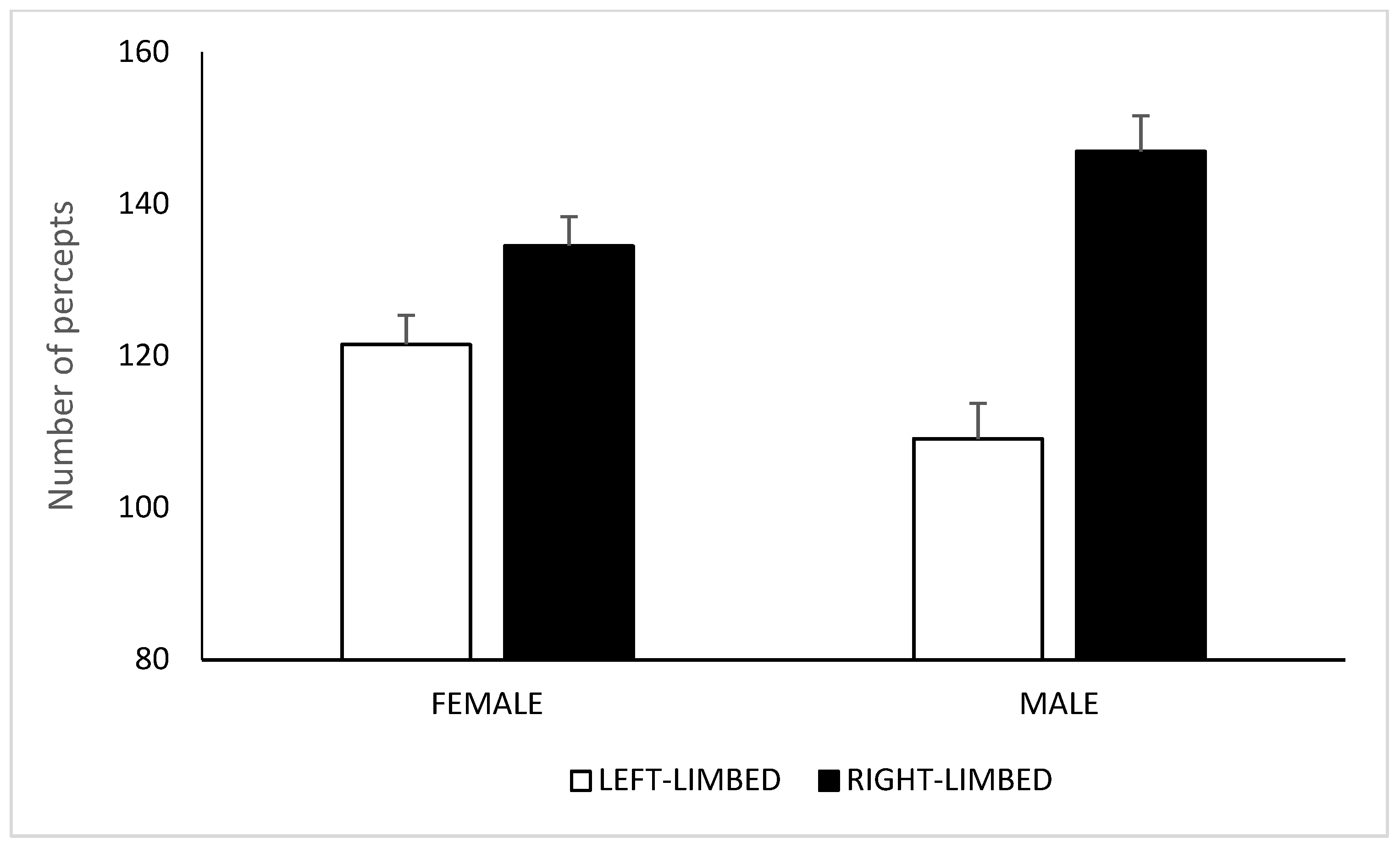
3.2. Results and Discussion
4. Experiment 3
4.1. Materials and Methods
4.1.1. Participants
4.1.2. Stimuli
4.1.3. Procedure
4.1.4. Data Analysis
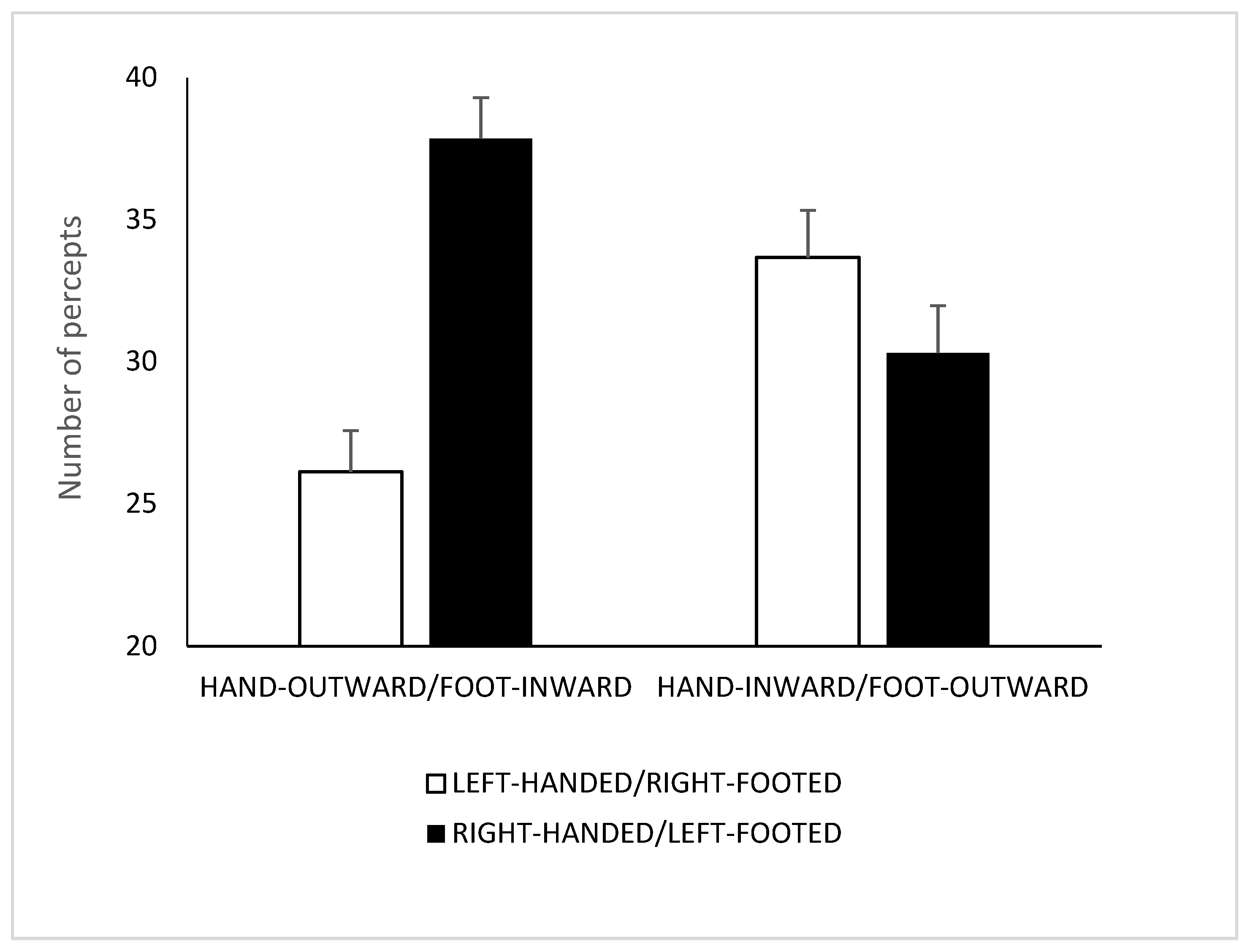
4.2. Results and Discussion
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bryden, M.P. Laterality: Functional Asymmetry in the Intact Brain; Academic Press: New York, NY, USA, 1982; ISBN 978-0-12-138180-6. [Google Scholar]
- Bryden, P.J. Lateral Preference, Skilled Behaviour and Task Complexity: Hand and Foot. In Side Bias: A Neuropsychological Perspective; Mandal, M.K., Bulman-Fleming, M.B., Tiwari, G., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 225–248. ISBN 978-0-306-46884-1. [Google Scholar]
- Elias, L.J.; Bryden, M.P. Footedness Is a Better Predictor of Language Lateralisation than Handedness. Laterality 1998, 3, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Gabbard, C.; Hart, S. Examining the Notion of Foot Dominance. In Side Bias: A Neuropsychological Perspective; Mandal, M.K., Bulman-Fleming, M.B., Tiwari, G., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 249–265. ISBN 978-0-306-46884-1. [Google Scholar]
- Peters, M. Neuropsychological Identification of Motor Problems: Can We Learn Something from the Feet and Legs That Hands and Arms Will Not Tell Us? Neuropsychol. Rev. 1990, 1, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Porac, C.; Coren, S. Lateral Preferences and Human Behavior; Springer: New York, NY, USA, 1981; ISBN 978-1-4613-8141-9. [Google Scholar]
- Tran, U.S.; Stieger, S.; Voracek, M. Evidence for General Right-, Mixed-, and Left-Sidedness in Self-Reported Handedness, Footedness, Eyedness, and Earedness, and a Primacy of Footedness in a Large-Sample Latent Variable Analysis. Neuropsychologia 2014, 62, 220–232. [Google Scholar] [CrossRef]
- Strauss, E. Hand, Foot, Eye and Ear Preferences and Performance on a Dichotic Listening Test. Cortex 1986, 22, 475–482. [Google Scholar] [CrossRef]
- Augustyn, C.; Peters, M. On the Relation between Footedness and Handedness. Percept. Mot. Ski. 1986, 63, 1115–1118. [Google Scholar] [CrossRef]
- Chapman, J.P.; Chapman, L.J.; Allen, J.J. The Measurement of Foot Preference. Neuropsychologia 1987, 25, 579–584. [Google Scholar] [CrossRef]
- Clymer, P.E.; Silva, P.A. Laterality, Cognitive Ability and Motor Performance in a Sample of Seven Year Olds. J. Hum. Mov. Stud. 1985, 11, 59–68. [Google Scholar]
- Nachshon, I.; Denno, D. Birth Order and Lateral Preferences. Cortex 1986, 22, 567–578. [Google Scholar] [CrossRef]
- Peters, M.; Durding, B.M. Footedness of Left- and Right-Handers. Am. J. Psychol. 1979, 92, 133–142. [Google Scholar] [CrossRef]
- Marzoli, D.; Prete, G.; Tommasi, L. Perceptual Asymmetries and Handedness: A Neglected Link? Front. Psychol. 2014, 5, 163. [Google Scholar] [CrossRef] [Green Version]
- Faurie, C.; Raymond, M. Handedness, Homicide and Negative Frequency-Dependent Selection. Proc. Biol. Sci. 2005, 272, 25–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grouios, G.; Tsorbatzoudis, H.; Alexandris, K.; Barkoukis, V. Do Left-Handed Competitors Have an Innate Superiority in Sports? Percept. Mot. Ski. 2000, 90, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Grouios, G.; Koidou, I.; Tsorbatzoudis, H.; Alexandris, K. Handedness in Sport. J. Hum. Mov. Stud. 2002, 43, 347–361. [Google Scholar]
- Raymond, M.; Pontier, D.; Dufour, A.B.; Møller, A.P. Frequency-Dependent Maintenance of Left Handedness in Humans. Proc. Biol. Sci. 1996, 263, 1627–1633. [Google Scholar] [CrossRef]
- Groothuis, T.G.G.; McManus, I.C.; Schaafsma, S.M.; Geuze, R.H. The Fighting Hypothesis in Combat: How Well Does the Fighting Hypothesis Explain Human Left-Handed Minorities? Ann. N. Y. Acad. Sci. 2013, 1288, 100–109. [Google Scholar] [CrossRef]
- Dochtermann, N.A.; Gienger, C.M.; Zappettini, S. Born to Win? Maybe, but Perhaps Only against Inferior Competition. Anim. Behav. 2014, 96, e1–e3. [Google Scholar] [CrossRef]
- Pollet, T.V.; Stulp, G.; Groothuis, T.G. Born to Win? Testing the Fighting Hypothesis in Realistic Fights: Left-Handedness in the Ultimate Fighting Championship. Anim. Behav. 2013, 86, 839–843. [Google Scholar] [CrossRef] [Green Version]
- Richardson, T.; Gilman, R.T. Left-Handedness Is Associated with Greater Fighting Success in Humans. Sci. Rep. 2019, 9, 15402. [Google Scholar] [CrossRef] [Green Version]
- Cingoz, Y.E.; Gursoy, R.; Ozan, M.; Hazar, K.; Dalli, M. Research on the Relation between Hand Preference and Success in Karate and Taekwondo Sports with Regards to Gender. Adv. Phys. Educ. 2018, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Gursoy, R. Effects of Left- or Right-Hand Preference on the Success of Boxers in Turkey. Br. J. Sports Med. 2009, 43, 142–144. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N. Pushing through Evolution? Incidence and Fight Records of Left-Oriented Fighters in Professional Boxing History. Laterality Asymmetries Body Brain Cogn. 2015, 20, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Ziyagil, M.A.; Gursoy, R.; Dane, S.; Yuksel, R. Left-Handed Wrestlers Are More Successful. Percept. Mot. Ski. 2010, 111, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Bisiacchi, P.S.; Ripoll, H.; Stein, J.; Simonet, P.; Azemar, G. Left-Handedness in Fencers: An Attentional Advantage? Percept. Mot. Ski. 1985, 61, 507–513. [Google Scholar] [CrossRef]
- Harris, L.J. In Fencing, What Gives Left-Handers the Edge? Views from the Present and the Distant Past. Laterality 2010, 15, 15–55. [Google Scholar] [CrossRef]
- Breznik, K. On the Gender Effects of Handedness in Professional Tennis. J. Sports Sci. Med. 2013, 12, 346–353. [Google Scholar]
- Holtzen, D.W. Handedness and Professional Tennis. Int. J. Neurosci. 2000, 105, 101–119. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N.; Strauss, B. Left-Handedness in Professional and Amateur Tennis. PloS ONE 2012, 7, e49325. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, S.R.; Young, C.A. “Evolutionary” Stable Strategy of Handedness in Major League Baseball. J. Comp. Psychol. 1996, 110, 164. [Google Scholar] [CrossRef]
- Lawler, T.P.; Lawler, F.H. Left-Handedness in Professional Basketball: Prevalence, Performance, and Survival. Percept. Mot. Ski. 2011, 113, 815–824. [Google Scholar] [CrossRef]
- Brooks, R.; Bussière, L.F.; Jennions, M.D.; Hunt, J. Sinister Strategies Succeed at the Cricket World Cup. Proc. Biol. Sci. 2004, 271, S64–S66. [Google Scholar] [CrossRef]
- Akpınar, S.; Bicer, B. Why Left-Handers/Footers Are Overrepresented in Some Sports? Montenegrin J. Sports Sci. Med. 2014, 3, 33–38. [Google Scholar]
- Tran, U.S.; Voracek, M. Footedness Is Associated with Self-Reported Sporting Performance and Motor Abilities in the General Population. Front. Psychol. 2016, 7, 1199. [Google Scholar] [CrossRef] [PubMed]
- McMorris, T.; Colenso, S. Anticipation of Professional Soccer Goalkeepers When Facing Right-and Left-Footed Penalty Kicks. Percept. Mot. Ski. 1996, 82, 931–934. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N. Zum Einfluss Des Anlaufwinkels Und Der Füßigkeit Des Schützen Auf Die Antizipation von Elfmeterschüssen. Z. Sportpsychol. 2015, 21, 63–73. [Google Scholar] [CrossRef]
- Thornton, I.M.; Vuong, Q.C.; Bülthoff, H.H. A Chimeric Point-Light Walker. Perception 2003, 32, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Lucafò, C.; Marzoli, D.; Prete, G.; Tommasi, L. Laterality Effects in the Spinning Dancer Illusion: The Viewing-from-above Bias Is Only Part of the Story. Br. J. Psychol. 2016, 107, 698–709. [Google Scholar] [CrossRef]
- Marzoli, D.; Lucafò, C.; Pagliara, A.; Cappuccio, R.; Brancucci, A.; Tommasi, L. Both Right- and Left-Handers Show a Bias to Attend Others’ Right Arm. Exp. Brain Res. 2015, 233, 415–424. [Google Scholar] [CrossRef]
- Marzoli, D.; Pagliara, A.; Prete, G.; Malatesta, G.; Lucafò, C.; Padulo, C.; Brancucci, A.; Tommasi, L. Lateralized Embodiment of Ambiguous Human Silhouettes: Data on Sex Differences. Data Brief 2019, 25, 104009. [Google Scholar] [CrossRef]
- Marzoli, D.; Lucafò, C.; Padulo, C.; Prete, G.; Giacinto, L.; Tommasi, L. Inversion Reveals Perceptual Asymmetries in the Configural Processing of Human Body. Front. Behav. Neurosci. 2017, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Marzoli, D.; Pagliara, A.; Prete, G.; Malatesta, G.; Lucafò, C.; Padulo, C.; Brancucci, A.; Tommasi, L. Hemispheric Asymmetries in the Processing of Body Sides: A Study with Ambiguous Human Silhouettes. Neurosci. Lett. 2017, 656, 114–119. [Google Scholar] [CrossRef]
- Marzoli, D.; Menditto, S.; Lucafò, C.; Tommasi, L. Imagining Others’ Handedness: Visual and Motor Processes in the Attribution of the Dominant Hand to an Imagined Agent. Exp. Brain Res. 2013, 229, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Marzoli, D.; Lucafò, C.; Rescigno, C.; Mussini, E.; Padulo, C.; Prete, G.; D’Anselmo, A.; Malatesta, G.; Tommasi, L. Sex-Specific Effects of Posture on the Attribution of Handedness to an Imagined Agent. Exp. Brain Res. 2017, 235, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Marzoli, D.; Mitaritonna, A.; Moretto, F.; Carluccio, P.; Tommasi, L. The Handedness of Imagined Bodies in Action and the Role of Perspective Taking. Brain Cogn. 2011, 75, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Marzoli, D.; Palumbo, R.; Di Domenico, A.; Penolazzi, B.; Garganese, P.; Tommasi, L. The Relation between Self-Reported Empathy and Motor Identification with Imagined Agents. PloS ONE 2011, 6, e14595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, C.L.; Stone, V.E.; Bozova, S.; Tanaka, J. The Body-Inversion Effect. Psychol. Sci. 2003, 14, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.L.; Stone, V.E.; Grubb, J.D.; McGoldrick, J.E. Turning Configural Processing Upside down: Part and Whole Body Postures. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Dane, S.; Erzurumluoglu, A. Sex and Handedness Differences in Eye-Hand Visual Reaction Times in Handball Players. Int. J. Neurosci. 2003, 113, 923–929. [Google Scholar] [CrossRef]
- Dane, S.; Gümüştekin, K. Correlation between Hand Preference and Distance of Focusing Points of Two Eyes in the Horizontal Plane. Int. J. Neurosci. 2002, 112, 1141–1147. [Google Scholar] [CrossRef]
- Judge, J.; Stirling, J. Fine Motor Skill Performance in Left- and Right-Handers: Evidence of an Advantage for Left-Handers. Laterality 2003, 8, 297–306. [Google Scholar] [CrossRef]
- Peterson, J.M.; Lansky, L.M. Left-Handedness among Architects: Some Facts and Speculation. Percept. Mot. Ski. 1974, 38, 547–550. [Google Scholar] [CrossRef]
- Annett, M. Left, Right, Hand and Brain: The Right Shift Theory; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 1985; ISBN 978-0-86377-018-0. [Google Scholar]
- Geschwind, N.; Galaburda, A.M. Cerebral Lateralization. Biological Mechanisms, Associations, and Pathology: II. A Hypothesis and a Program for Research. Arch. Neurol. 1985, 42, 521–552. [Google Scholar] [CrossRef] [PubMed]
- McLean, J.M.; Ciurczak, F.M. Bimanual Dexterity in Major League Baseball Players: A Statistical Study. N. Engl. J. Med. 1982, 307, 1278–1279. [Google Scholar] [CrossRef] [PubMed]
- Nass, R.D.; Gazzaniga, M.S. Cerebral Lateralization and Specialization in Human Central Nervous System. In Comprehensive Physiology; American Cancer Society: Atlanta, GA, USA, 1987; pp. 701–761. ISBN 978-0-470-65071-4. [Google Scholar]
- Aggleton, J.P.; Wood, C.J. Is There a Left-Handed Advantage in “Ballistic” Sports? Int. J. Sport Psychol. 1990, 21, 46–57. [Google Scholar]
- Hemenway, D.A. Bimanual Dexterity in Baseball Players. N. Engl. J. Med. 1983, 309, 1587–1588. [Google Scholar] [PubMed]
- Wood, C.J.; Aggleton, J.P. Handedness in “fast Ball” Sports: Do Left-Handers Have an Innate Advantage? Br. J. Psychol. 1989, 80, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Coren, S. The Left-Hander Syndrome: The Causes and Consequences of Left-Handedness; Free Press: New York, NY, USA, 1992; ISBN 978-0-02-906682-9. [Google Scholar]
- Hagemann, N. The Advantage of Being Left-Handed in Interactive Sports. Atten. Percept. Psychophys. 2009, 71, 1641–1648. [Google Scholar] [CrossRef]
- Loffing, F.; Schorer, J.; Hagemann, N.; Baker, J. On the Advantage of Being Left-Handed in Volleyball: Further Evidence of the Specificity of Skilled Visual Perception. Atten. Percept. Psychophys. 2012, 74, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Loffing, F.; Hagemann, N.; Schorer, J.; Baker, J. Skilled Players’ and Novices’ Difficulty Anticipating Left- vs. Right-Handed Opponents’ Action Intentions Varies across Different Points in Time. Hum. Mov. Sci. 2015, 40, 410–421. [Google Scholar] [CrossRef]
- Schorer, J.; Loffing, F.; Hagemann, N.; Baker, J. Human Handedness in Interactive Situations: Negative Perceptual Frequency Effects Can Be Reversed! J. Sports Sci. 2012, 30, 507–513. [Google Scholar] [CrossRef]
- Jacobs, A.; Pinto, J.; Shiffrar, M. Experience, Context, and the Visual Perception of Human Movement. J. Exp. Psychol. Hum. Percept. Perform. 2004, 30, 822–835. [Google Scholar] [CrossRef] [Green Version]
- Troje, N.F.; McAdam, M. The Viewing-from-Above Bias and the Silhouette Illusion. i-Percept. 2010, 1, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertamini, M.; Bode, C.; Bruno, N. The Effect of Left-Right Reversal on Film: Watching Kurosawa Reversed. i-Percept. 2011, 2, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Simmons, J.P.; Nelson, L.D.; Simonsohn, U. A 21 Word Solution; Social Science Research Network: Rochester, NY, USA, 2012. [Google Scholar]
- Liu, C.-H.; Tzeng, O.J.L.; Hung, D.L.; Tseng, P.; Juan, C.-H. Investigation of Bistable Perception with the “Silhouette Spinner”: Sit Still, Spin the Dancer with Your Will. Vis. Res. 2012, 60, 34–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmaso, D.; Longoni, A.M. Problems in the Assessment of Hand Preference. Cortex J. Devoted Study Nerv. Syst. Behav. 1985, 21, 533–549. [Google Scholar] [CrossRef]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Chen, M.M.Z.; Karlinsky, A.; Welsh, T.N. Hand, but Not Foot, Cues Generate Increases in Salience at the Pointed-at Location. Acta Psychol. 2020, 210, 103165. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucafò, C.; Marzoli, D.; Zdybek, P.; Malatesta, G.; Smerilli, F.; Ferrara, C.; Tommasi, L. The Bias toward the Right Side of Others Is Stronger for Hands than for Feet. Symmetry 2021, 13, 146. https://doi.org/10.3390/sym13010146
Lucafò C, Marzoli D, Zdybek P, Malatesta G, Smerilli F, Ferrara C, Tommasi L. The Bias toward the Right Side of Others Is Stronger for Hands than for Feet. Symmetry. 2021; 13(1):146. https://doi.org/10.3390/sym13010146
Chicago/Turabian StyleLucafò, Chiara, Daniele Marzoli, Przemysław Zdybek, Gianluca Malatesta, Federico Smerilli, Cosimo Ferrara, and Luca Tommasi. 2021. "The Bias toward the Right Side of Others Is Stronger for Hands than for Feet" Symmetry 13, no. 1: 146. https://doi.org/10.3390/sym13010146