Contrasting the Effect of Forest Landscape Condition to the Resilience of Species Diversity in a Human Modified Landscape: Implications for the Conservation of Tree Species


Abstract
:1. Introduction
2. Materials and Methods
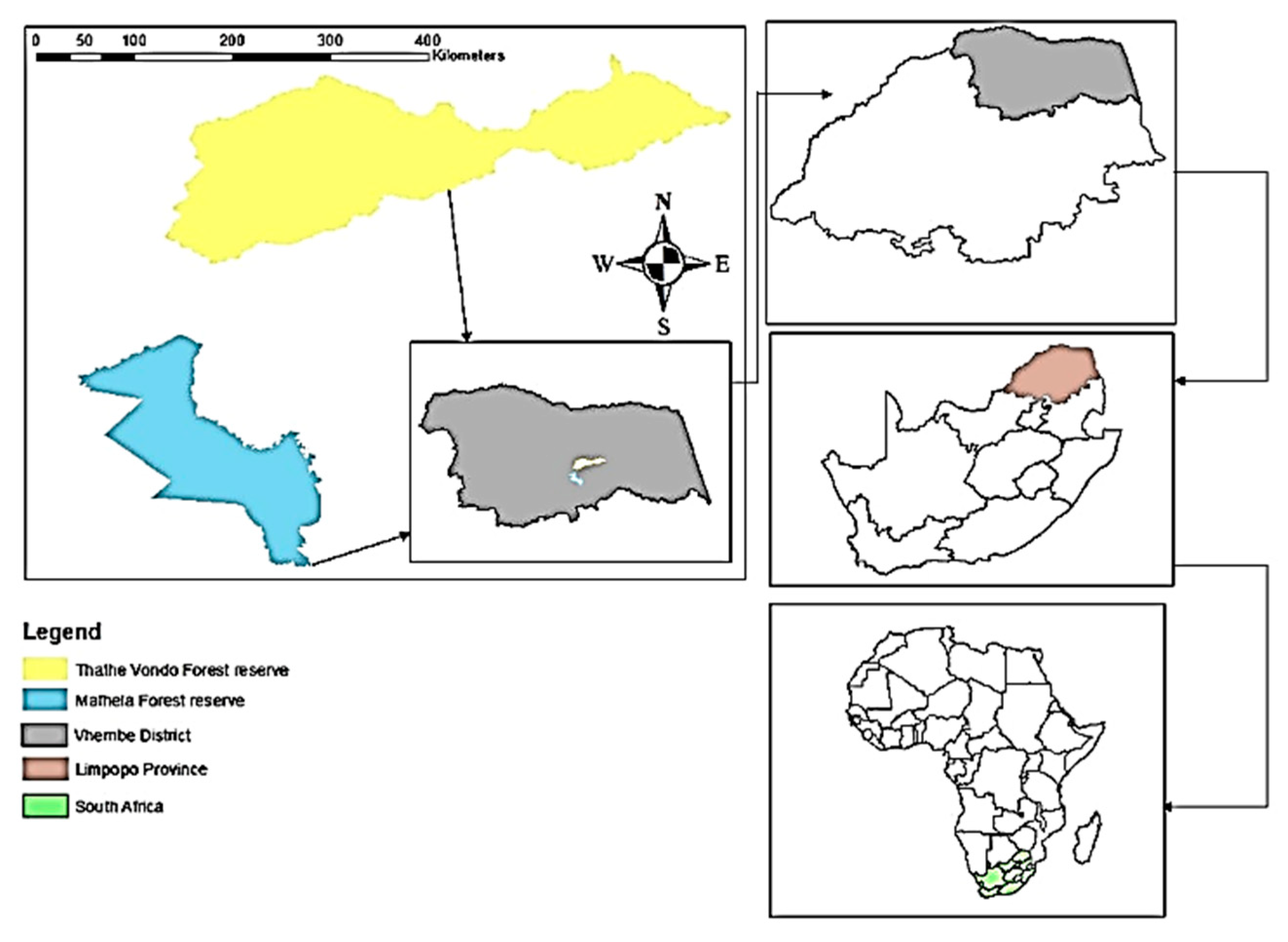
2.1. Study Area
- (a)
- Human modified landscape (HMFL): consisting of three tree-based traditional land use regimes under the custody of traditional authorities and the local community. These are:
- (b)
- Trees Along Streams and Rivers (TATR): local community members are not allowed to harvest live trees, but occasionally access the place for livestock grazing, watering, and shading (intermediately disturbed);
- (c)
- Common Resource Use Zones (CRUZ): this is an open access area for the harvesting of wild food, construction materials, livestock browsing and grazing, traditional medicines and others (highly disturbed);
- (d)
- Culturally Protected Forest Areas (CPA): these include sacred/holy forests that are protected by royal families for cultural values and only accessible to them (minimally disturbed).
- (e)
- State indigenous forests (SIF): these are fragmented forest patches, with minimal to no human disturbance and are legally protected by government conservation agencies.
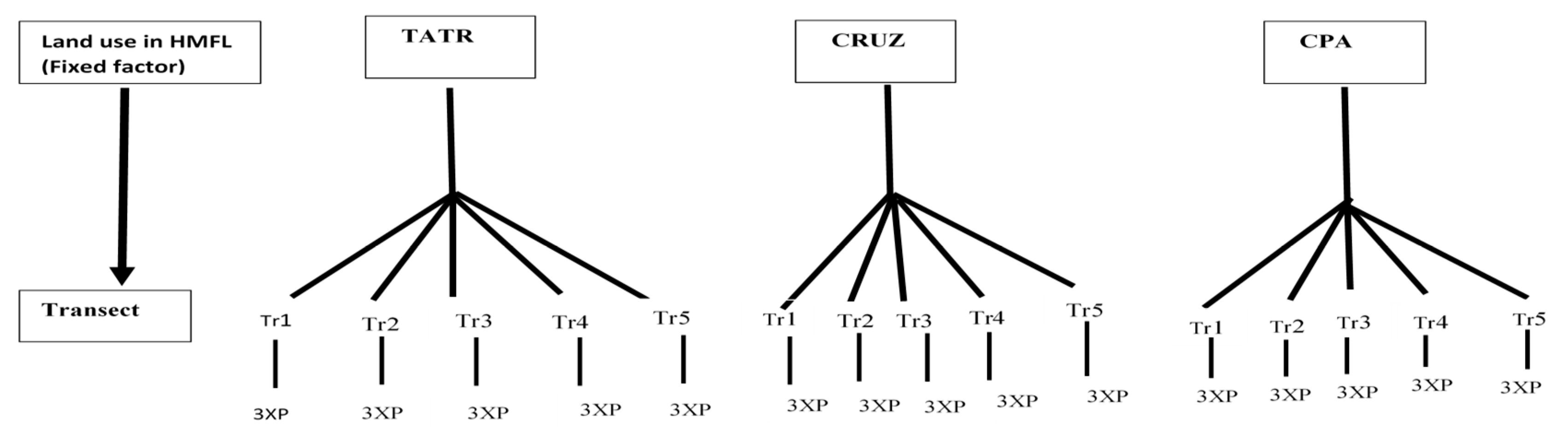
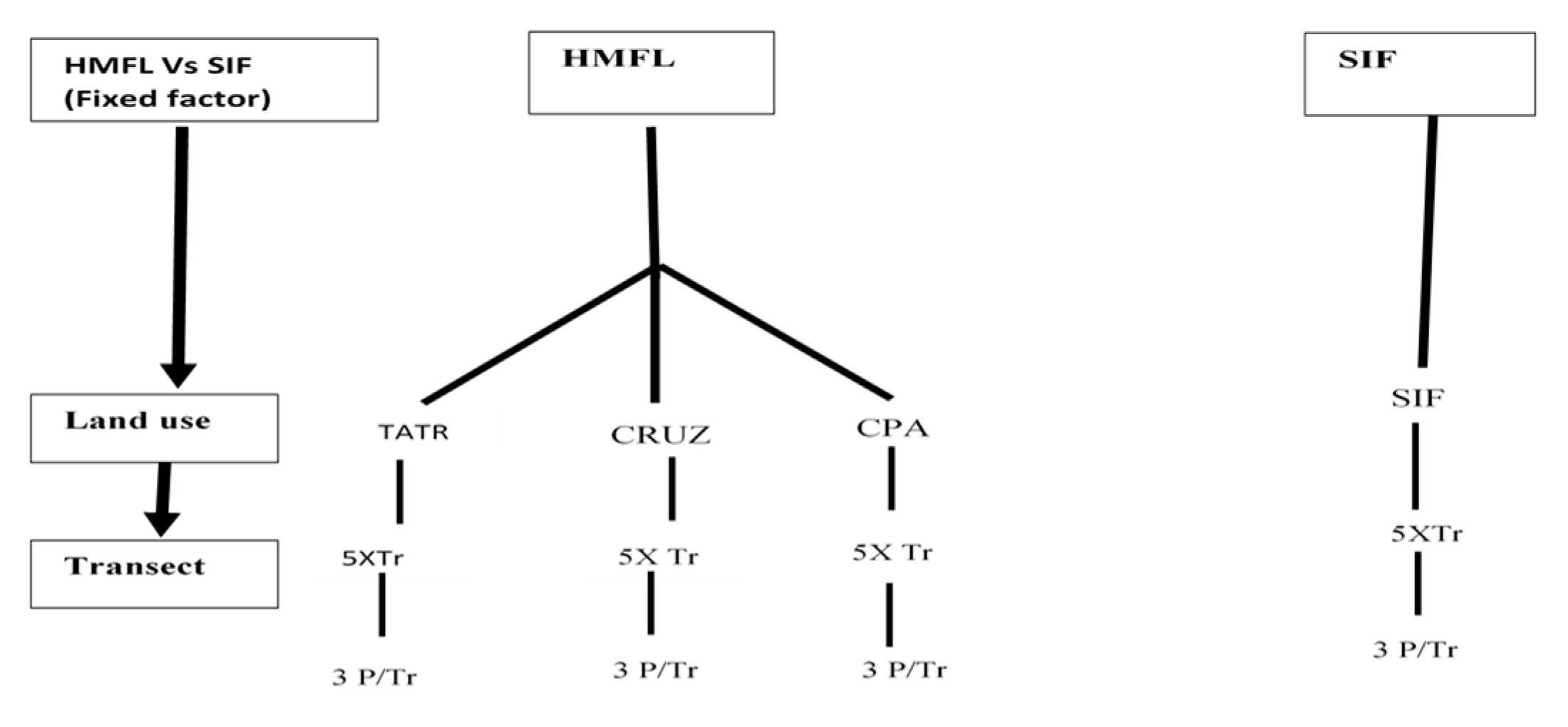
2.2. Sampling Design
2.3. Data Collection
2.3.1. Measurement of Tree Species Assemblage
2.3.2. Change Drivers of Overall β-Diversity of FRs
2.4. Statistical Analysis
2.4.1. The Effect of Land Use Regimes on the Difference of Mean β-Diversity in HMFL
2.4.2. Species Richness Difference (Δ ) along the Land Use Gradient
2.4.3. Change in Species Composition (Identity Replacement) along Land Use Gradient
2.4.4. A Local Contribution of the Human Modified Forest Landscape to Overall β-Diversity of Forest Reserve
2.5. The Influence of Land Use and Other Environmental Drivers in Overall β-Diversity of Human Modified Forest Reserve
3. Result
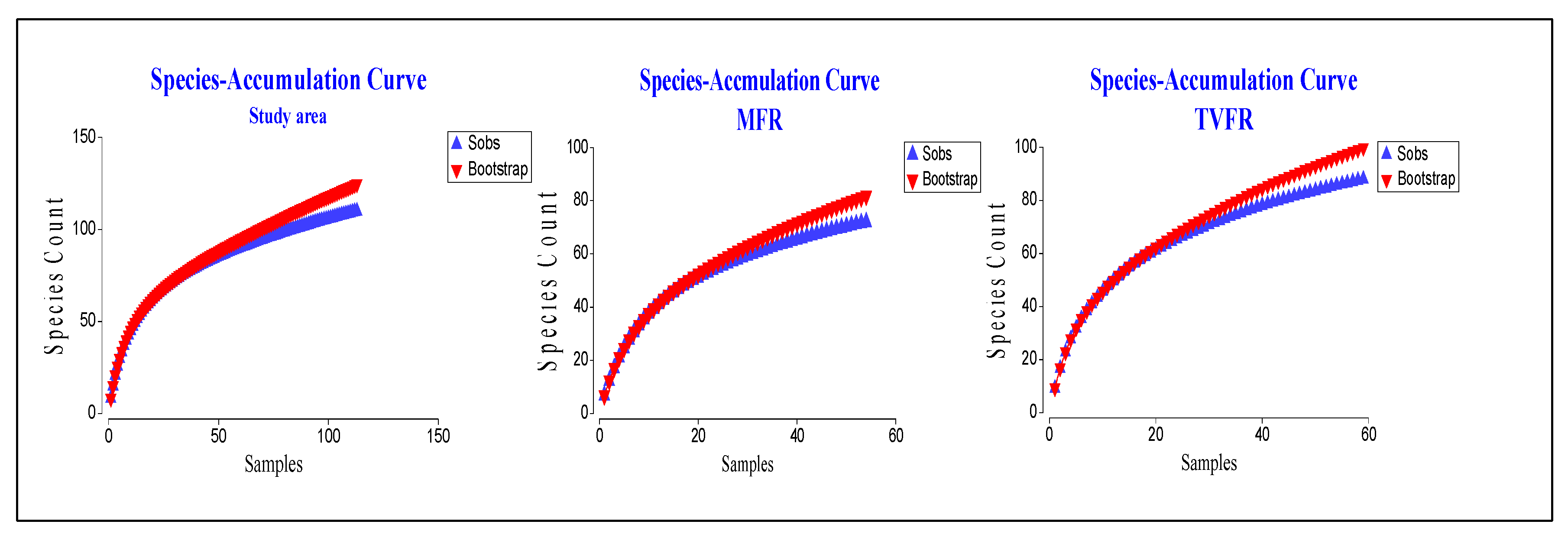
3.1. Description of Forest Complexity Condition
The Effect of Land Use Gradient on of Mean β-Diversity of HMFL
3.2. Species Richness along Land Use Intensity Gradient
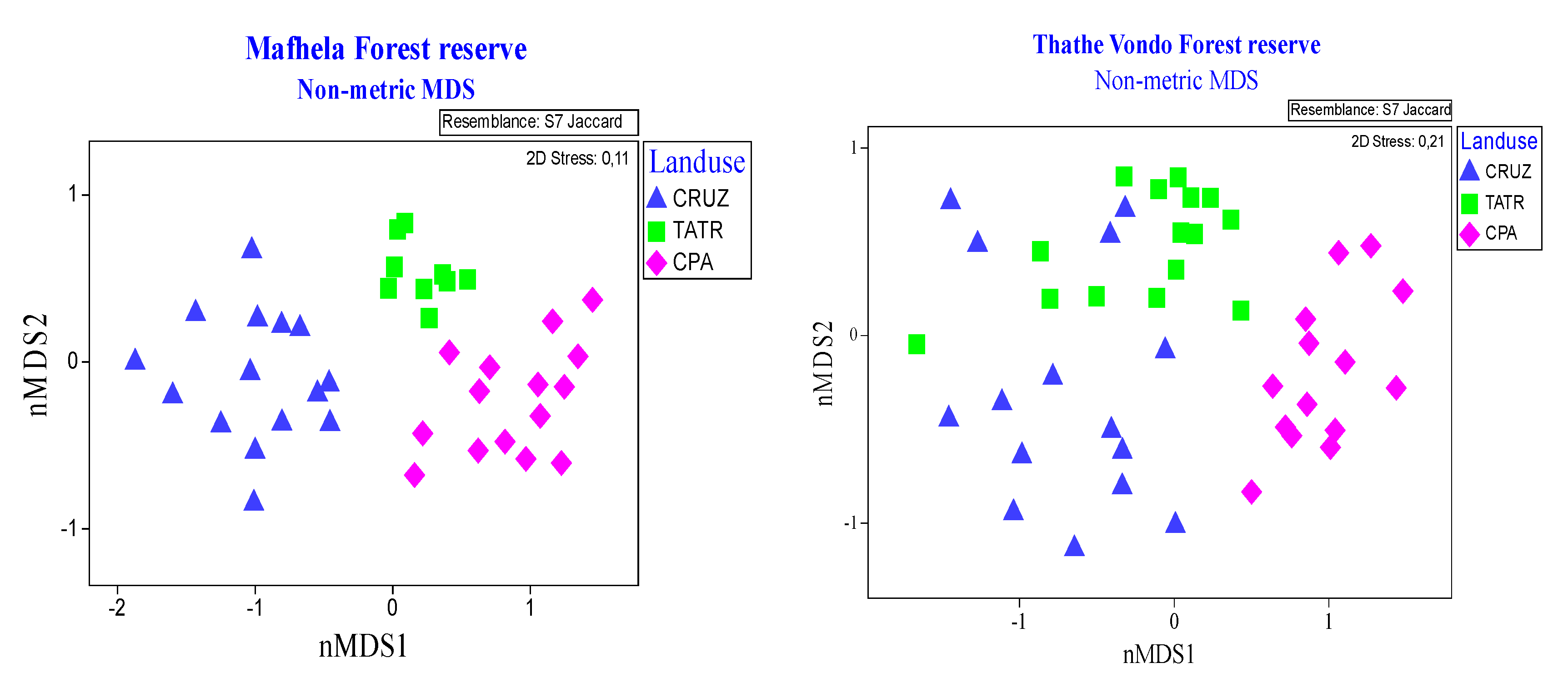
3.3. Change in Species Composition (Identity Replacement) along Land Use Gradients
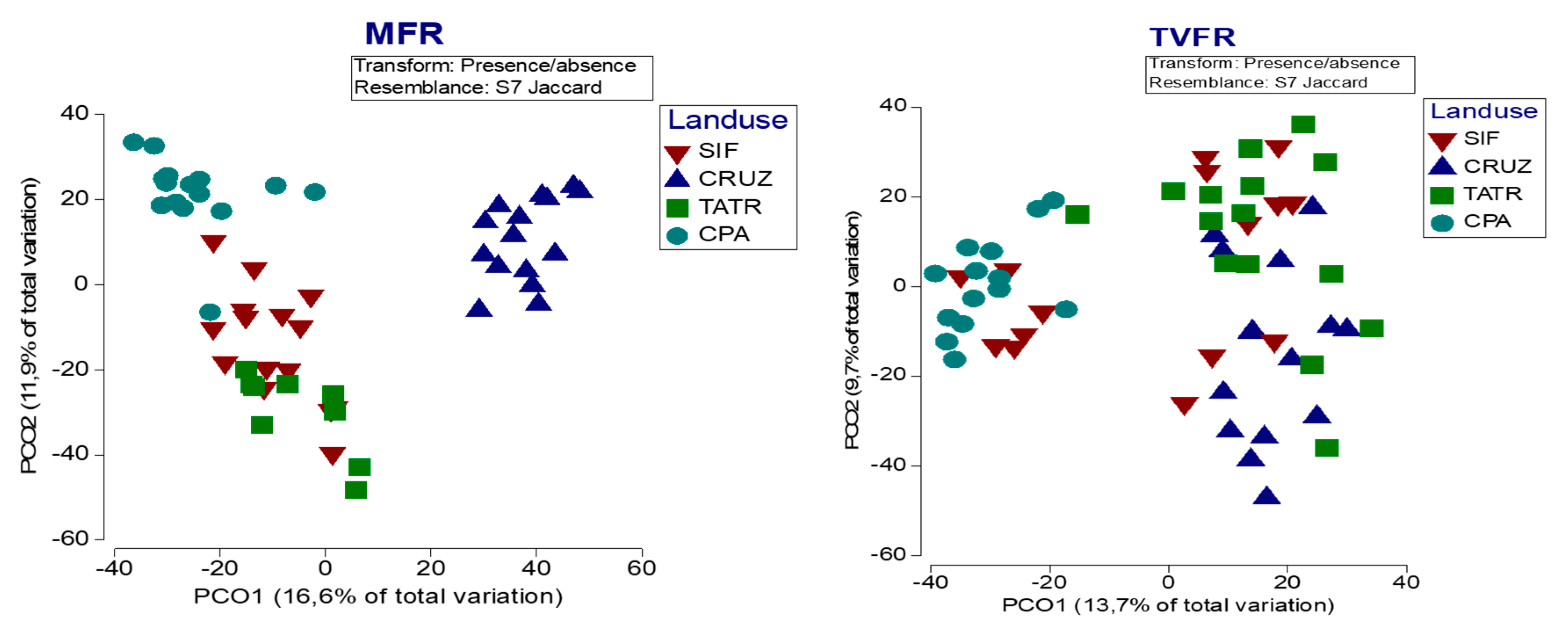
3.4. The Local Contribution of the Human Modified Forest Landscape to Overall β-Diversity of Forest Reserve
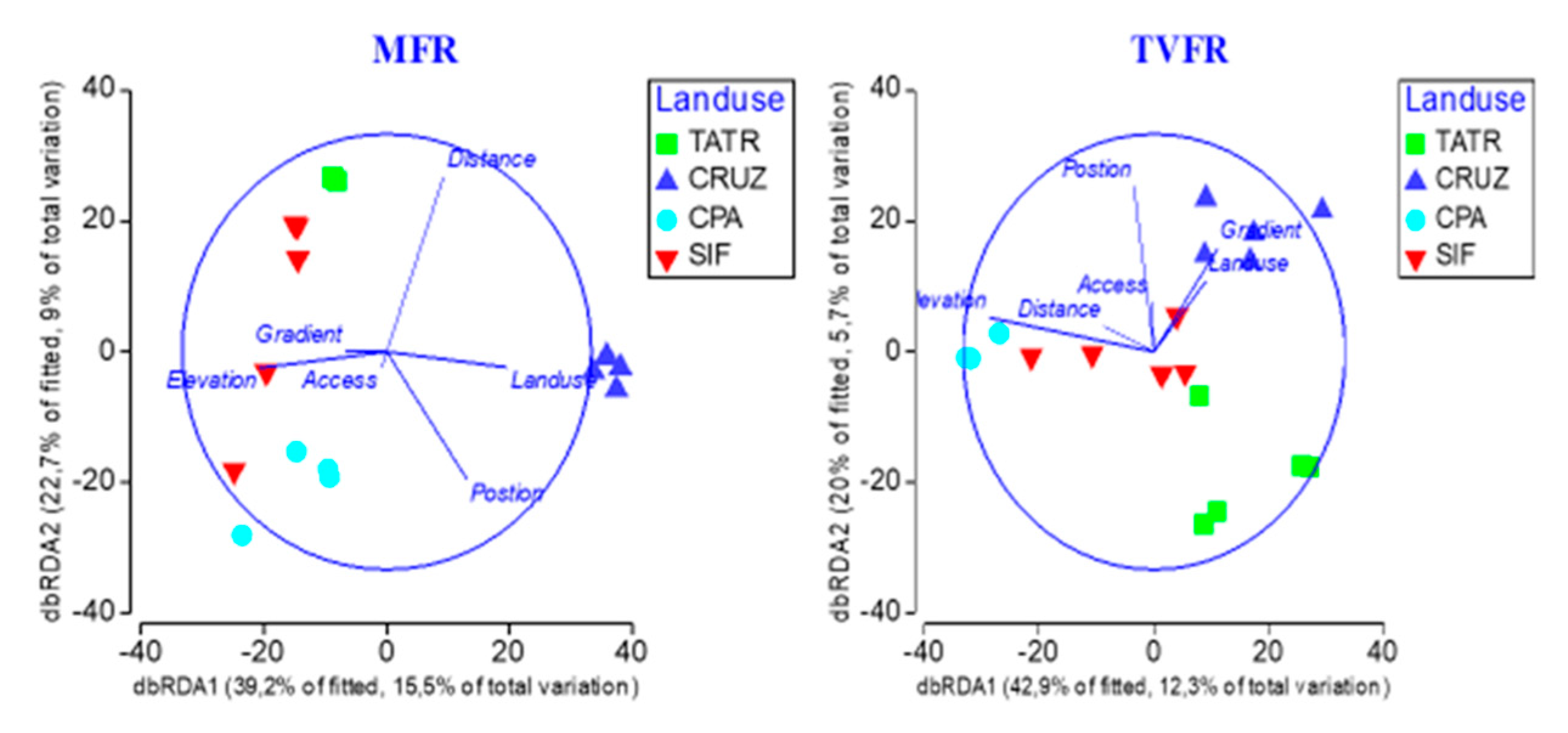
3.5. Change Drivers Influencing Overall β-Diversity of Forest Reserves
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harvey, E.; Gounand, I.; Ward, C.L.; Altermatt, F. Bridging ecology and conservation: From ecological networks to ecosystem function. J. Appl. Ecol. 2017, 54, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.E.; Dudley, N.; Segan, D.B.; Hockings, M. The performance and potential of protected areas. Nature 2014, 515, 67. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.P.; Arroyo-Rodríguez, V.; Fahrig, L.; Martínez-Ramos, M.; Tabarelli, M. In the hope for biodiversity-friendly tropical landscapes. Trends Ecol. Evol. 2013, 28, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar]
- Willig, M.R.; Presley, S.J. Biodiversity and disturbance. Encycl. Anthr. 2018, 3, 45–51. [Google Scholar]
- Van Gemerden, B.S.; Olff, H.; Parren, M.P.; Bongers, F. The pristine rain forest? Remnants of historical human impacts on current tree species composition and diversity. J. Biogeogr. 2003, 30, 1381–1390. [Google Scholar] [CrossRef]
- Connell, J.H. Tropical rainforests and coral reefs as open non-equilibrium systems. In Population Dynamics; Blackwell: Oxford, UK, 1979; pp. 141–163. [Google Scholar]
- Fox, J.W. The intermediate disturbance hypothesis should be abandoned. Trends Ecol. Evol. 2013, 28, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.; Gardner, T.A.; Louzada, J.; Peres, C.A. Measuring the conservation value of tropical primary forests: The effect of occasional species on estimates of biodiversity uniqueness. PLoS ONE 2010, 5, e9609. [Google Scholar] [CrossRef]
- Noble, I.R.; Dirzo, R. Forests as human-dominated ecosystems. Science 1997, 277, 522–525. [Google Scholar] [CrossRef] [Green Version]
- Saura, S.; Bertzky, B.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 2018, 219, 53–67. [Google Scholar] [CrossRef]
- Alroy, J. Effects of habitat disturbance on tropical forest biodiversity. Proc. Natl. Acad. Sci. USA 2017, 114, 6056–6061. [Google Scholar] [CrossRef] [Green Version]
- Joppa, L.N.; Pfaff, A. High and far: Biases in the location of protected areas. PLoS ONE 2009, 4, e8273. [Google Scholar] [CrossRef] [PubMed]
- Lele, S.; Wilshusen, P.; Brockington, D.; Seidler, R.; Bawa, K. Beyond exclusion: Alternative approaches to biodiversity conservation in the developing tropics. Curr. Opin. Environ. Sustain. 2010, 2, 94–100. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Melo, F.P.; Martínez-Ramos, M.; Bongers, F.; Chazdon, R.L.; Meave, J.A.; Norden, N.; Santos, B.A.; Leal, I.R.; Tabarelli, M. Multiple successional pathways in human-modified tropical landscapes: New insights from forest succession, forest fragmentation and landscape ecology research. Biol. Rev. 2017, 92, 326–340. [Google Scholar] [CrossRef] [PubMed]
- Resasco, J.; Bruna, E.M.; Haddad, N.M.; Banks-Leite, C.; Margules, C.R. The contribution of theory and experiments to conservation in fragmented landscapes. Ecography 2017, 40, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Yeboah, D.; Chen, H.Y. Diversity–disturbance relationship in forest landscapes. Landsc. Ecol. 2016, 31, 981–987. [Google Scholar] [CrossRef]
- Hillebrand, H.; Blasius, B.; Borer, E.T.; Chase, J.M.; Downing, J.A.; Eriksson, B.K.; Filstrup, C.T.; Harpole, W.S.; Hodapp, D.; Larsen, S.; et al. Biodiversity change is uncoupled from species richness trends: Consequences for conservation and monitoring. J. Appl. Ecol. 2018, 55, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Jiao, F.; Li, Y.H.; Kallenbach, R.L. Anthropogenic disturbances are key to maintaining the biodiversity of grasslands. Sci. Rep. 2016, 6, 22132. [Google Scholar] [CrossRef] [Green Version]
- Assede, E.S.; Adomou, A.C.; Sinsin, B. Secondary succession and factors determining change in soil condition from fallow to savannah in the Sudanian Zone of Benin. Phytocoenologia 2012, 42, 181–189. [Google Scholar] [CrossRef]
- Laurance, W.F.; Nascimento, H.E.; Laurance, S.G.; Andrade, A.; Ewers, R.M.; Harms, K.E.; Luizao, R.C.; Ribeiro, J.E. Habitat fragmentation, variable edge effects, and the landscape-divergence hypothesis. PLoS ONE 2007, 2, e1017. [Google Scholar] [CrossRef]
- Derroire, G.; Balvanera, P.; Castellanos-Castro, C.; Decocq, G.; Kennard, D.K.; Lebrija-Trejos, E.; Leiva, J.A.; Odén, P.C.; Powers, J.S.; Rico-Gray, V.; et al. Resilience of tropical dry forests—A meta-analysis of changes in species diversity and composition during secondary succession. Oikos 2016, 125, 1386–1397. [Google Scholar] [CrossRef] [Green Version]
- Ghazoul, J.; Burivalova, Z.; Garcia-Ulloa, J.; King, L.A. Conceptualizing forest degradation. Trends Ecol. Evol. 2015, 30, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legendre, P.; Gauthier, O. Statistical methods for temporal and space–time analysis of community composition data. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Holmes, I.; Vergara-Asenjo, G.; Miller, W.N.; Cunampio, M.; Cunampio, R.B.; Cunampio, M.B.; Potvin, C. A comparison of influences on the landscape of two social-ecological systems. Land Use Policy 2016, 57, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.T.; Xu, B.; Li, M. Vegetation patterns and species diversity along elevation and disturbance gradients in the Baihua Mountain Reserve, Beijing, China. Mt. Res. Dev. 2013, 33, 170–178. [Google Scholar] [CrossRef]
- Shova, T.; Hubacek, K. Drivers of illegal resource extraction: An analysis of Bardia National Park, Nepal. J. Environ. Manag. 2011, 92, 156–164. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batary, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol. Rev. 2013, 87, 661–685. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Pardini, R.; de Arruda Bueno, A.; Gardner, T.A.; Prado, P.I.; Metzger, J.P. Beyond the fragmentation threshold hypothesis: Regime shifts in biodiversity across fragmented landscapes. PLoS ONE 2010, 5, e13666. [Google Scholar] [CrossRef]
- Symes, C.T.; Olaf Wirminghaus, J.; Downs, C.T.; Louette, M. Species richness and seasonality of forest avifauna in three South African Afromontane forests. Ostrich 2002, 73, 106–113. [Google Scholar] [CrossRef]
- Araia, M.G.; Chirwa, P.W. Revealing the Predominance of Culture over the Ecological Abundance of Resources in Shaping Local People’s Forest and Tree Species Use-behavior: The Case of the Vhavenda People, South Africa. Sustainability 2019, 11, 3143. [Google Scholar] [CrossRef] [Green Version]
- Araia, M.G.; Chirwa, P.W. Nurturing forest resources in the Vhavenda community, South Africa: Factors influencing non-compliance behaviour of local people to state conservation rules. South. For. J. For. Sci. 2019, 81, 357–366. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, R.K. Permanova+ for Primer: Guide to Software and Statistical Methods; PRIMER-E-2015; Primer-E Limited: Plymouth, UK, 2008. [Google Scholar]
- Sheil, D.; Puri, R.; Wan, M.; Basuki, I.; van Heist, M.; Liswanti, N. Recognizing local people’s priorities for tropical forest biodiversity. AMBIO A J. Hum. Environ. 2006, 35, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheil, D.; Puri, R.K.; Basuki, I.; van Heist, M.; Wan, M.; Liswanti, N.; Sardjono, M.A.; Samsoedin, I.; Sidiyasa, K.; Permana, E.; et al. Exploring Biological Diversity, Environment, and Local People’s Perspectives in Forest Landscapes: Methods for a Multidisciplinary Landscape Assessment; CIFOR SMK Grafika Desa Putera: Jakarta, Indonsia, 2002. [Google Scholar]
- Gillison, A.N. A Field Manual for Rapid Vegetation Classification and Survey for General Purposes; Centre for International Forestry Research: Bogor, Indonsia, 2006. [Google Scholar]
- Gillison, A.N.; Liswanti, N.; Rachman, I.A. Rapid Ecological Assessment of Kerinci Seblat National Park Buffer Zone; Centre for International Forestry Research: Bogor, Indonsia, 1996. [Google Scholar]
- Hairiah, K.; Sitompul, S.M.; van Noordwijk, M.; Palm, C. Methods for Sampling Carbon Stocks above and below Ground; ICRAF: Nairobi, Kenya, 2001; pp. 1–23. [Google Scholar]
- Young, A. Tropical Soils and Soil Survey; Cambridge University Press: Cambridge, UK, 1980; Volume 9. [Google Scholar]
- Clarke, K.R.; Gorley, R.N.; Anderson, M.A. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E-2015; Primer-E Limited: Plymouth, UK, 2015. [Google Scholar]
- Foggo, A.; Attrill, M.J.; Frost, M.T.; Rowden, A.A. Estimating marine species richness: An evaluation of six extrapolative techniques. Mar. Ecol. Prog. Ser. 2003, 248, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Avolio, M.L.; Pierre, K.J.L.; Houseman, G.R.; Koerner, S.E.; Grman, E.; Isbell, F.; Johnson, D.S.; Wilcox, K.R. A framework for quantifying the magnitude and variability of community responses to global change drivers. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Coetzee, B.W.; Gaston, K.J.; Chown, S.L. Local scale comparisons of biodiversity as a test for global protected area ecological performance: A meta-analysis. PLoS ONE 2014, 9, e105824. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Mayor, S.J.; Cahill, J.F., Jr.; He, F.; Boutin, S. Scaling disturbance instead of richness to better understand anthropogenic impacts on biodiversity. PLoS ONE 2015, 10, e0125579. [Google Scholar] [CrossRef] [Green Version]
- Ellis, E.C.; Antill, E.C.; Kreft, H. All is not loss: Plant biodiversity in the Anthropocene. PLoS ONE 2012, 7, e30535. [Google Scholar] [CrossRef] [Green Version]
- Li, S.P.; Cadotte, M.W.; Meiners, S.J.; Pu, Z.; Fukami, T.; Jiang, L. Convergence and divergence in a long-term old-field succession: The importance of spatial scale and species abundance. Ecol. Lett. 2016, 19, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Munyati, C.; Sinthumule, N.I. Cover Gradients and the Forest-Community Frontier: Indigenous Forests under Communal Management at Vondo and Xanthia, South Africa. J. Sustain. For. 2014, 33, 757–775. [Google Scholar] [CrossRef]
- Hansen, A.J.; DeFries, R. Ecological mechanisms linking protected areas to surrounding lands. Ecol. Appl. 2007, 17, 974–988. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Use Regimes | Number of Species in MFR | Sampling Effectiveness (%) | Number of Species in TVFR | Sampling Effectiveness (%) | ||
|---|---|---|---|---|---|---|
| Sob | Sboot * | Sob | Sboot * | |||
| TATR | 18 | 21 | 85.71 | 55 | 68 | 80.88 |
| CRUZ | 33 | 39 | 84.61 | 57 | 69 | 82.61 |
| CPA | 26 | 34 | 78.80 | 26 | 30 | 86.67 |
| SIF | 39 | 45 | 86.67 | 54 | 65 | 83.07 |
| Change Drivers | Thathe Vondo Forest Reserve (TVFR) | Mafhela Forest Reserve (MFR) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Land Use | Distance | Access | Position | Gradient | Land Use | Distance | Access | Position | Gradient | |
| Distance | −0.58 | −0.55 | ||||||||
| Access | 0.24 | −0.17 | 0.24 | 0.64 | ||||||
| Position | 0.30 | 0.48 | −0.06 | 0.17 | 0.13 | −0.08 | ||||
| Gradient | 0.19 | −0.57 | 0.24 | −0.75 | 0.04 | −0.43 | −0.12 | 0.66 | ||
| Elevation | −0.31 | 0.48 | −0.47 | 0.09 | −0.14 | −0.91 | 0.74 | 0.44 | −0.06 | −0.33 |
| HMFLs | Source | Df | SS | MS | Pseudo-F | P(Perm) | Unique Terms |
|---|---|---|---|---|---|---|---|
| MFR | La | 2 | 44,509 | 22,254 | 7.398 | 0.001 | 987 |
| Tr | 10 | 30,082 | 3008.20 | 1.537 | 0.001 | 996 | |
| Res | 26 | 50,865 | 1956.30 | ||||
| Total | 38 | 125,460 | |||||
| TVFR | La | 2 | 31,482 | 15,741 | 3.8127 | 0.001 | 998 |
| Tr | 12 | 49,717 | 4143.10 | 1.8422 | 0.001 | 994 | |
| Res | 29 | 65,222 | 2249 | ||||
| Total | 43 | 146,690 |
| Pairwise Land Use Regimes | HMFLs in MFR | HMFLs in TVFR | ||||
|---|---|---|---|---|---|---|
| T | p(Perm) | Av. DJ (%) | T | p(Perm) | Av. DJ (%) | |
| (TATR & CRUZ) | 2.53 | 0.016 | 61 | 1.59 | 0.012 | 38 |
| (TATR & CPA) | 2.77 | 0.022 | 55 | 2.22 | 0.008 | 51 |
| (CRUZ & CPA) | 2.84 | 0.008 | 61 | 2.05 | 0.006 | 50 |
| HMFLs | Source | Df | SS | MS | Psuedo-F | P (Perm) | Unique Perms |
|---|---|---|---|---|---|---|---|
| MFR | La | 2 | 3.740 | 1.870 | 5.466 | 0.029 | 965 |
| Tr | 10 | 3.421 | 0.342 | 0.970 | 0.529 | 999 | |
| Res | 26 | 9.715 | 0.374 | ||||
| Total | 38 | 16.876 | |||||
| TVFR | La | 2 | 19.333 | 9.667 | 15.454 | 0.001 | 998 |
| Tr | 12 | 7.508 | 0.626 | 1.046 | 0.455 | 999 | |
| Res | 29 | 17.350 | 0.598 | ||||
| Total | 43 | 44.822 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araia, M.G.; Chirwa, P.W.; Assédé, E.S.P. Contrasting the Effect of Forest Landscape Condition to the Resilience of Species Diversity in a Human Modified Landscape: Implications for the Conservation of Tree Species. Land 2020, 9, 4. https://doi.org/10.3390/land9010004
Araia MG, Chirwa PW, Assédé ESP. Contrasting the Effect of Forest Landscape Condition to the Resilience of Species Diversity in a Human Modified Landscape: Implications for the Conservation of Tree Species. Land. 2020; 9(1):4. https://doi.org/10.3390/land9010004
Chicago/Turabian StyleAraia, Mulugheta Ghebreslassie, Paxie Wanangwa Chirwa, and Eméline Sêssi Pélagie Assédé. 2020. "Contrasting the Effect of Forest Landscape Condition to the Resilience of Species Diversity in a Human Modified Landscape: Implications for the Conservation of Tree Species" Land 9, no. 1: 4. https://doi.org/10.3390/land9010004