



Composition of the Anuran Community in a Forest Management Area in Southeastern Amazonia
 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
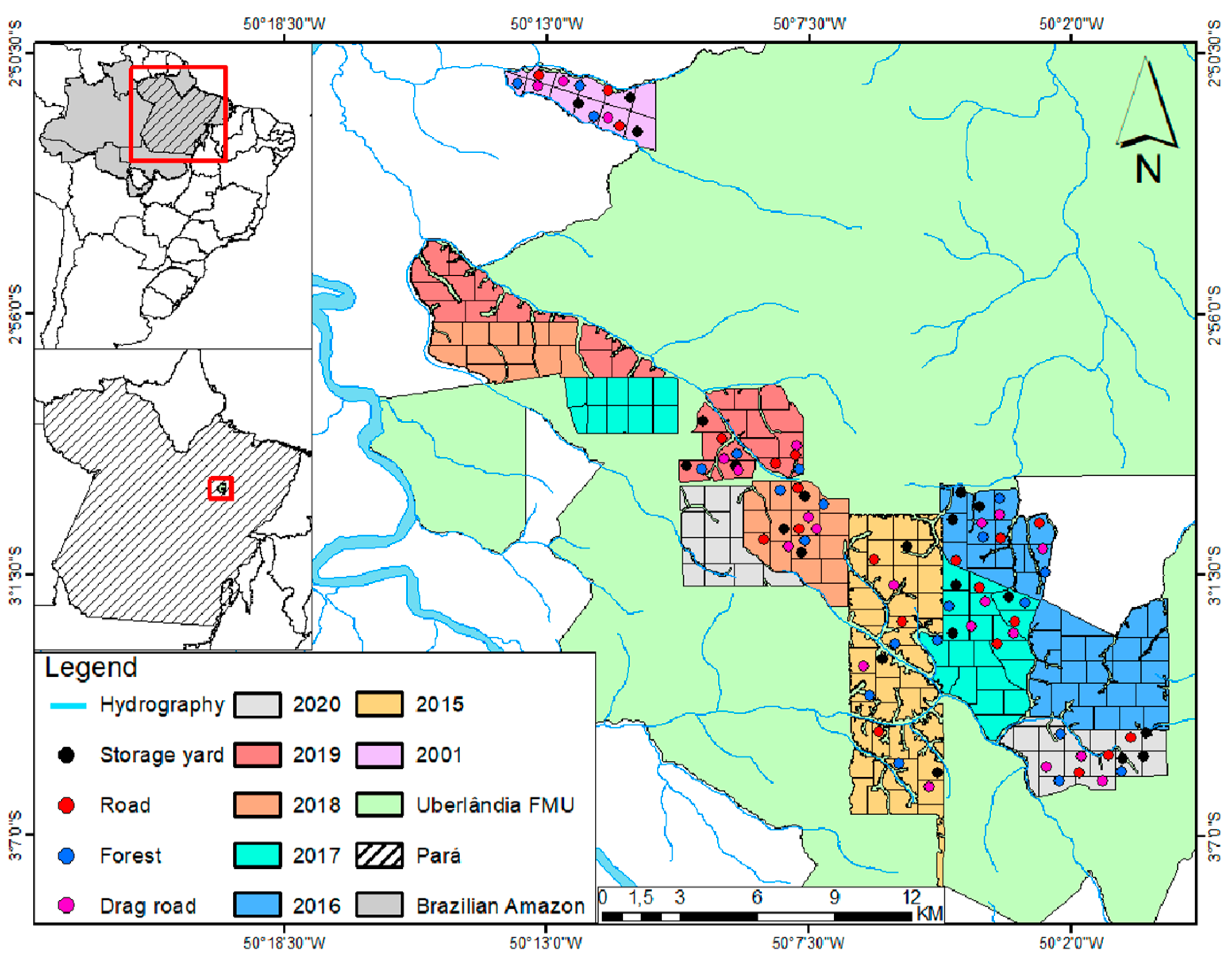
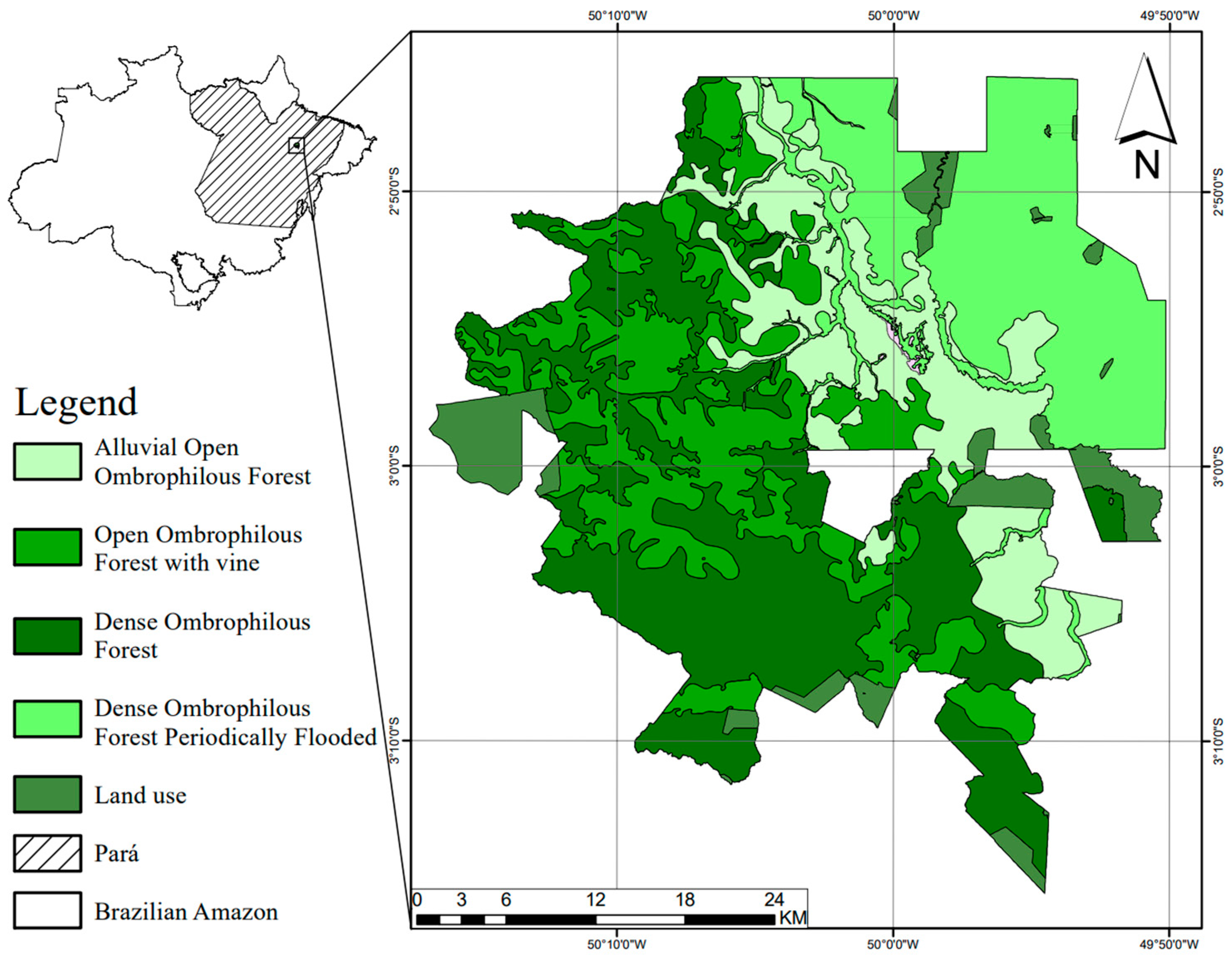
2.1. Study Area
2.2. Sampling
2.3. Sample Preparation
2.4. Data Analysis
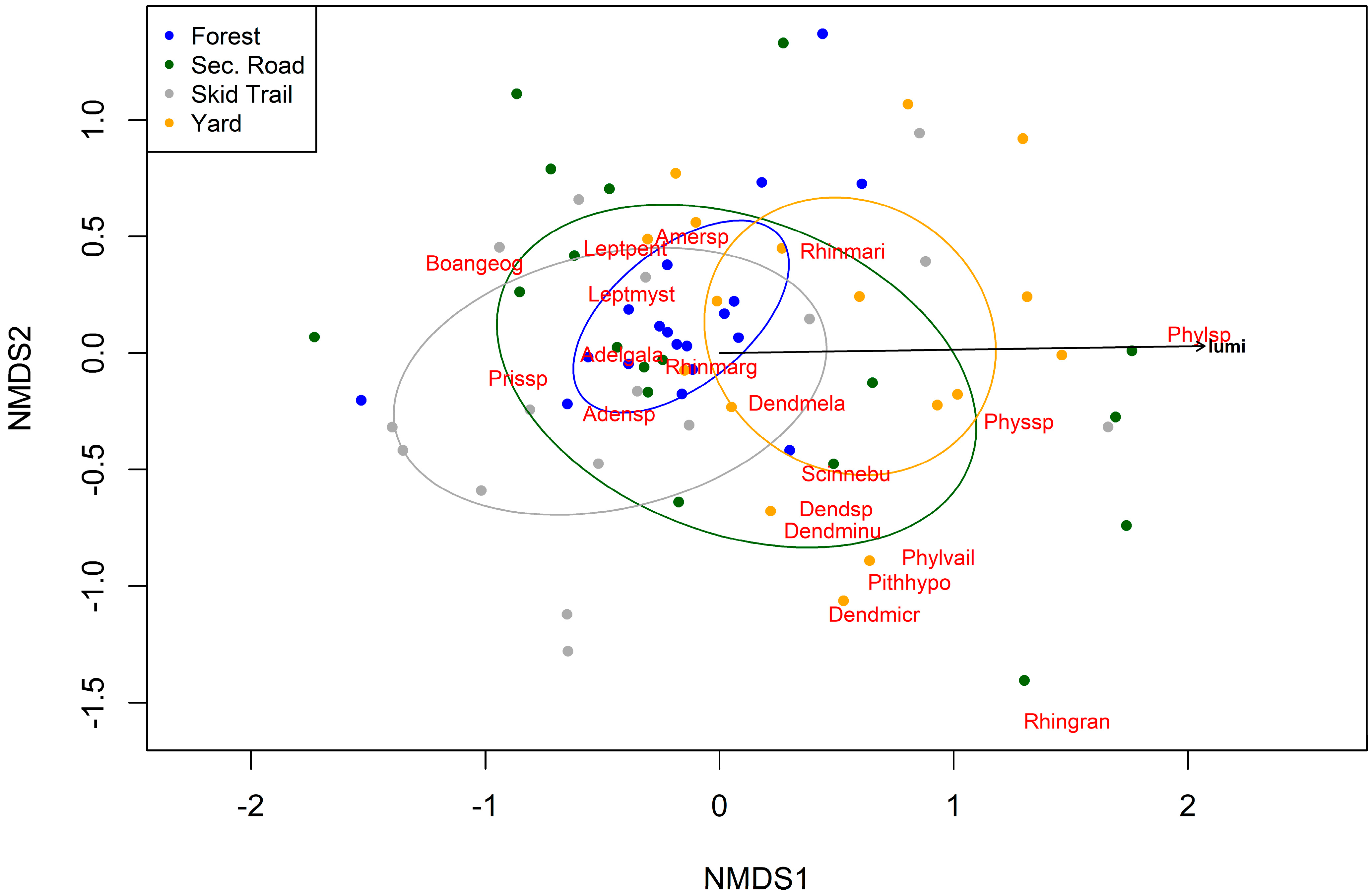
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maxwell, S.L.; Fuller, R.A.; Brooks, T.M.; Watson, J.E. Biodiversity: The ravages of guns, nets and bulldozers. Nat. News 2016, 536, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, V.D.; Patrick, D.A.; Hunter, M.L., Jr.; Calhoun, A.J. The role of forest harvesting and subsequent vegetative regrowth in determining patterns of amphibian habitat use. For. Ecol. Manag. 2012, 270, 16–174. [Google Scholar] [CrossRef]
- Leclère, D.; Obersteiner, M.; Barrett, M.; Butchart, S.H.; Chaudhary, A.; De Palma, A.; Young, L.; DeClerck, F.A.J.; Di Marco, M.; Doelman, J.C.; et al. Bending the curve of terrestrial biodiversity needs an integrated strategy. Nature 2020, 585, 551–556. [Google Scholar] [CrossRef] [PubMed]
- The International Tropical Timber Organization. State of Art Report on Improved Forest Harvesting and Reduced Impact Logging in Asia Pacific Region. 2020. Available online: https://www.itto.int/files/itto_project_db_input/2727/Technical/ppd%2019-99-1%20rev1(F)%20e.pdf (accessed on 18 October 2019).
- Food and Agriculture Organization of the United Nations. Reduced Impact Logging in Tropical Forests. 2004. Available online: http://www.fao.org/tempref/docrep/fao/008/j4290e/J4290e.pdf (accessed on 17 October 2019).
- Macpherson, A.J.; Carter, D.R.; Schulze, M.D.; Vidal, E.; Lentini, M.W. The sustainability of timber production from Eastern Amazonian forests. Land Use Policy 2012, 29, 339–350. [Google Scholar] [CrossRef]
- Dodd, C.K.; Smith, L.L. Habitat destruction and alteration. Historical trends and future prospects for Amphibians. In Amphibian Conservation; Semlitsch, R.D., Ed.; Smithsonian Institution Press: Washington, DC, USA, 2003; pp. 94–112. [Google Scholar]
- Burivalova, Z.; Şekercioğlu, Ç.H.; Koh, L.P. Thresholds of logging intensity to maintain tropical forest biodiversity. Curr. Biol. 2014, 24, 1893–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuomisto, H.; Ruokolainen, K.; Kalliola, R.; Linna, A.; Danjoy, W.; Rodriguez, Z. Dissecting amazonian biodiversity. Science 1997, 26, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.A. Amphibians as models for studying environmental change. ILAR J. 2007, 48, 270–277. [Google Scholar] [CrossRef]
- Bitar, Y.O.; Juen, L.; Pinheiro, L.C.; Santos-Costa, M.C.D. Anuran beta diversity in a mosaic anthropogenic landscape in transitional Amazon. Bioone 2015, 49, 75–82. [Google Scholar] [CrossRef]
- Vaira, M. Report of a breeding aggregation extirpation of an endemic marsupial frog, Gastrotheca christiani, in Argentina. Froglog 2003, 60, 3. [Google Scholar]
- Kpan, T.F.; Ernst, R.; Roedel, M.O. Follow the forest: Slow resilience of West African rainforest frog assemblages after selective logging. For. Ecol. Manag. 2021, 497, 119489. [Google Scholar] [CrossRef]
- Putz, F.E.; Zuidema, P.A.; Synnott, T.; Peña-Claros, M.; Pinard, M.A.; Sheil, D.; Zagt, R.; Vanclay, J.K.; Sist, P.; Gourlet-Fleury, S.; et al. Sustaining conservation values in selectively logged tropical forests: The attained and the attainable. Conserv. Lett. 2012, 5, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Berthinussen, A.; Richardson, O.C.; Altringham, J.D. Bat Conservation: Global Evidence for the Effects of Interventions; Conservation Evidence Series Synopses; University of Cambridge: Cambridge, UK, 2021. [Google Scholar]
- Camargo, J.L.C.; Kapos, V. Complex edge effects on soil moisture and microclimate in central Amazonian Forest. J. Trop. Ecol. 1995, 11, 205–221. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P. The Effects of Logging on Reptiles and Amphibians of Tropical Forests. In The Cutting Edge; Columbia University Press: Cambridge, UK, 2001; pp. 239–260. [Google Scholar]
- Fredericksen, N.J.; Fredericksen, T.S.; Flores, B.; Rumiz, D. Wildlife use of different-sized logging gaps in a tropical dry forest. Trop. Ecol. 1999, 40, 167–175. [Google Scholar]
- Fredericksen, N.J.; Fredericksen, T.S. Terrestrial wildlife responses to logging and fire in a Bolivian tropical humid forest. Biodivers. Conserv. 2002, 11, 27–38. [Google Scholar] [CrossRef]
- Popescu, V.D.; Hunter, M.L., Jr. Clear-cutting affects habitat connectivity for a forest amphibian by decreasing permeability to juvenile movements. Ecol. Appl. 2011, 21, 1283–1295. [Google Scholar] [CrossRef] [Green Version]
- Tilghman, J.M.; Ramee, S.W.; Marsh, D.M. Meta-analysis of the effects of canopy removal on terrestrial salamander populations in North America. Biol. Conserv. 2012, 152, 1–9. [Google Scholar] [CrossRef]
- Lemckert, F. Impacts of selective logging on frogs in a forested area of northern New South Wales. Biol. Conserv. 1999, 89, 321–328. [Google Scholar] [CrossRef]
- Fredericksen, N.J.; Fredericksen, T.S. Impacts of selective logging on amphibians in a Bolivian tropical humid forest. For. Ecol. Manag. 2004, 191, 275–282. [Google Scholar] [CrossRef]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Sodhi, N.S.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef]
- Miranda, D.; Venancio, N.; De Alburquerque, S. Rapid survey of the herpetofauna in an area of forest management in eastern Acre, Brazil. Check List 2014, 10, 893–899. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.P.; Woodcock, P.; Edwards, F.A.; Larsen, T.H.; Hsu, W.W.; Benedick, S.; Wilcove, D.S. Reduced-impact logging and biodiversity conservation: A case study from Borneo. Ecol. Appl. 2012, 22, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adum, G.B.; Ofori-Boateng, C.; Oduro, W. Re-discovery of the Giant West African Squeaker, Arthroleptis krokosua Ernst, Agyei & Rödel, 2008 (Amphibia: Anura: Arthroleptidae) in two forests of south-western Ghana with observations on the species’ variability and habitat preferences. Zootaxa 2011, 2744, 34–38. [Google Scholar]
- Cushman, S.A. Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar] [CrossRef]
- Hölting, M.; Bovolo, C.I.; Ernst, R. Facing complexity in tropical conservation: How reduced impact logging and climatic extremes affect beta diversity in tropical amphibian assemblages. Biotropica 2016, 48, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Brüning, L.Z.; Krieger, M.; Meneses-Pelayo, E.; Eisenhauer, N.; Pinilla, M.P.R.; Reu, B.; Ernst, R. Land-use heterogeneity by small-scale agriculture promotes amphibian diversity in montane agroforestry systems of northeast Colombia. Agric. Ecosyst. Environ. 2018, 264, 15–23. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Moraes Gonçalves, J.L.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Zeitschrift. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Silva, J.N.M.; De Carvalho, J.O.P.; Lopes, J. Um sistema silvicultural policíclico para produção sustentada de madeira na Amazônia Brasileira. In Simpósio de Silvicultura na Amazônia Oriental: Contribuições do Projeto Embrapa/DFID; Embrapa Amazônia Oriental: Belém, PA, Brazil, 1999. [Google Scholar]
- Santos, I.V.; Balieiro, S.; Mattos, R.; Ramos, Y.; Santos, R.N.; Quadro, A.G. Relatório Técnico de Identificação, Manejo e Monitoramento de Florestas de Alto Valor de Conservação (HCVS) da Unidade de Manejo Florestal; UMF Uberlândia: Portel, PA, Brazil, 2016. [Google Scholar]
- Bitar, Y.O.; Tuomisto, H.; Pinheiro, L.C.; Juen, L.; Santos-Costa, M.C. Species turnover in Amazonian frogs: Low predictability and large differences among forests. Biotropica 2017, 49, 695–705. [Google Scholar] [CrossRef]
- Ministério do Meio Ambiente. Norma de Execução IBAMA n°1 de 24/04/2007. Brazilian Oficial Register 30 April 2007, No. 82; Brazilian Government: Brasília, Brazil, 2007. Available online: https://www.normasbrasil.com.br/norma/norma-de-execucao-12007_90937.html (accessed on 2 July 2021).
- Karsten, R.J.; Meilby, H.; Larsen, J.B. Regeneration and management of lesser known timber species in the Peruvian Amazon following disturbance by logging. For. Ecol. Manag. 2014, 327, 76–85. [Google Scholar] [CrossRef]
- Crump, M.L.; Scott, N.J., Jr. Visual encounter surveys. In Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians; Heyer, W.R., Donnelly, M.A., McDiarmid, R.W., Hayek, L.C., Foster, M.S., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1994. [Google Scholar]
- Gascon, C. Population-and community-level analyses of species occurrences of central Amazonian rainforest tadpoles. Ecology 1991, 72, 1731–1746. [Google Scholar] [CrossRef]
- Ernst, R.; Rödel, M.O. Anthropogenically induced changes of predictability in tropical anuran assemblages. Ecology 2005, 86, 3111–3118. [Google Scholar] [CrossRef]
- de Oliveira, E.A.; da Silva, L.A.; Silva, E.A.P.; Guimarães, K.L.A.; Penhacek, M.; Martínez, J.G.; Hernández-Ruz, E.J.; Rodrigues, L.R.R.; Santana, D.J. Four new species of Pristimantis Jiménez de la Espada, 1870 (Anura: Craugastoridae) in the eastern Amazon. PLoS ONE 2020, 15, e0229971. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.J.; Townsend, C.R.; Reynolds, R.P.; MacCulloch, R.D.; Lathrop, A. Amphibians and reptiles of Guyana, South America: Illustrated keys, annotated species accounts, and a biogeographic synopsis. Proc. Biol. Soc. Wash. 2013, 125, 317–578. [Google Scholar] [CrossRef]
- Hero, J.M. An illustrated key to tadpoles occurring in the Central Amazon rainforest, Manaus, Amazonas, Brasil. Amazoniana 1990, 11, 201–262. [Google Scholar]
- Dubeux, M.J.M.; Nascimento, F.A.C.D.; Lima, L.R.; Magalhães, F.D.M.; Silva, I.R.S.D.; Gonçalves, U.; Mott, T.; Almeida, J.P.F.; Correia, L.L.; Garda, A.A.; et al. Morphological characterization and taxonomic key of tadpoles (Amphibia: Anura) from the northern region of the Atlantic Forest. Biota Neotrop. 2020, 20, e20180718. [Google Scholar] [CrossRef]
- Pereyra, M.O.; Blotto, B.L.; Baldo, D.; Chaparro, J.C.; Ron, S.R.; Elias-Costa, A.J.; Faivovich, J.; Iglesias, P.P.; Venegas, P.J.; Thomé, M.T.C.; et al. Evolution in the genus Rhinella: A total evidence phylogenetic analysis of neotropical true toads (Anura: Bufonidae). Bull. Am. Mus. Nat Hist. 2021, 447, 1–156. [Google Scholar] [CrossRef]
- Fouquet, A.; Martinez, Q.; Zeidler, L.; Courtois, E.A.; Gaucher, P.; Blanc, M.; Kok, P.J.; Lima, J.D.; Souza, S.M.; Rodrigues, M.T.; et al. Cryptic diversity in the Hypsiboas semilineatus species group (Amphibia, Anura) with the description of a new species from the eastern Guiana Shield. Zootaxa 2016, 4084, 79–104. [Google Scholar] [CrossRef]
- Orrico, V.G.; Grant, T.; Faivovich, J.; Rivera-Correa, M.; Rada, M.A.; Lyra, M.L.; Haddad, C.F.; Cassini, C.S.; Valdujo, P.H.; Schargel, W.E.; et al. The phylogeny of Dendropsophini (Anura: Hylidae: Hylinae). Cladistics 2021, 37, 73–105. [Google Scholar] [CrossRef]
- Warton, D.I. Regularized sandwich estimators for analysis of high dimensional data using generalized estimating equations. Biometrics 2011, 67, 116–123. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R package for modelbased analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; McGraw-Hill Book Company: New York, NY, USA, 1994; p. 670. [Google Scholar]
- Silva, E.P.; Mendes-Pinto, T.J.; Júnior, L.H.C.; Sales, M.E.P. Riqueza de espécies de anfíbios anuros em um fragmento florestal na área urbana de Manaus, Amazonas, Brasil. Rev. Biol. Farm. 2011, 5, 131–144. [Google Scholar]
- Filho, R.G.Q.; Mendes-Pinto, T.J.; Fragoso, A.C. Anurofauna da área do Centro Universitário Nilton Lins, Manaus, Amazonas, Brasil (Amphibia; Anura). Rev. Biol. Farm. 2010, 4, 13–19. [Google Scholar]
- Dunn, R.R. Recovery of faunal communities during tropical forest regeneration. Conserv. Biol. 2004, 18, 302–309. [Google Scholar] [CrossRef]
- Rozendaal, D.M.; Bongers, F.; Aide, T.M.; Alvarez-Dávila, E.; Ascarrunz, N.; Balvanera, P.; Poorter, L.; Becknell, J.M.; Bentos, T.V.; Brancalion, P.H.S.; et al. Biodiversity recovery of Neotropical secondary forests. Sci. Adv. 2019, 5, eaau3114. [Google Scholar] [CrossRef] [Green Version]
- Hernández, O.O.; Urbina, C.N.; Martínez, R.M. Recovery of amphibian and reptile assemblages during old-field succession of tropical rain forests. Biotropica 2015, 47, 377–388. [Google Scholar] [CrossRef]
- Asad, S.; Abrams, J.F.; Guharajan, R.; Lagan, P.; Kissing, J.; Sikui, J.; Wilting, A.; Rödel, M.-O. Amphibian responses to conventional and reduced impact logging. For. Ecol. Manag. 2021, 484, 118949. [Google Scholar] [CrossRef]
- Keenan, R.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Ernst, R.; Linsenmair, K.E.; Rödel, M.O. Diversity erosion beyond the species level: Dramatic loss of functional diversity after selective logging in two tropical amphibian communities. Biol. Conserv. 2006, 133, 143–155. [Google Scholar] [CrossRef]
- Haggerty, C.J.; Crisman, T.L.; Rohr, J.R. Effects of forestry-driven changes to groundcover and soil moisture on amphibian desiccation, dispersal, and survival. Ecol. Appl. 2019, 29, e01870. [Google Scholar] [CrossRef]
- Haddad, C.F.B.; Prado, C.P.A. Reproductive modes in frogs and their unexpected diversity in the Atlantic Forest of Brazil. BioScience 2005, 55, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Bitar, Y.O.; Pinheiro, L.P.; Abe, P.S.; Santos-Costa, M.C. Species composition and reproductive modes of anurans from a transitional Amazonian forest, Brazil. Zoologia 2012, 29, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Lindenmayer, D.B.; Franklin, J.F.; Lõhmus, A.; Baker, S.C.; Bauhus, J.; Beese, W.; Gustafsson, L.; Brodie, A.; Kiehl, B.; Kouki, J.; et al. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012, 5, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Olden, J.D.; Poff, N.L.; Douglas, M.R.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Olden, J.D.; Rooney, T.P. On defining and quantifying biotic homogenization. Glob. Ecol. Biogeogr. 2006, 15, 113–120. [Google Scholar] [CrossRef]
- Caldwell, J.P. Structure and behavior of Hyla geographica tadpole schools, with comments on classification of group behavior in tadpoles. Copeia 1989, 1989, 938–948. [Google Scholar] [CrossRef]
- Vallan, D.; Andreone, F.; Raherisoa, V.H.; Dolch, R. Does selective wood exploitation affect amphibian diversity? The case of An’Ala, a tropical rainforest in eastern Madagascar. Oryx 2004, 38, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Hillers, A.; Veith, M.; RÖDEL, M.O. Effects of forest fragmentation and habitat degradation on West African leaf-litter frogs. Conserv. Biol. 2008, 22, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Sinsch, U. Postmetamorphic dispersal and recruitment of first breeders in a Bufo calamita metapopulation. Oecologia 1997, 112, 42–47. [Google Scholar] [CrossRef]
- Crump, M.L. Anuran reproductive modes: Evolving perspectives. J. Herpetol. 2015, 49, 12–16. [Google Scholar] [CrossRef]
- Goutte, S.; Sah, H.A.; Grafe, T.U. Environmental correlates of species richness and composition of riparian anuran communities in rainforests of north-western Borneo: A metacommunity perspective. Herpetol. J. 2017, 27, 25–32. [Google Scholar]
- Bernarde, P.S.; Macedo, L.C. Impacto do desmatamento e formação de pastagens sobre a anurofauna de serapilheira em Rondônia. Iheringia Ser. Zool. 2008, 98, 454–459. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.R.; Norman, J. The herpetofauna of forest litter plots from Cameroon, Africa. US Fish Wildl. Res. Rep. 1982, 13, 145–150. [Google Scholar]
- Heinen, J.T. Comparisons of the leaf litter herpetofauna in abandoned cacao plantations and primary rain forest in Costa Rica: Some implications for faunal restoration. Biotropica 1992, 27, 431–439. [Google Scholar] [CrossRef]
- Young, J.E.; Christian, K.A.; Donnellan, S.; Tracy, C.R.; Parry, D. Comparative analysis of cutaneous evaporative water loss in frogs demonstrates correlation with ecological habits. Physiol. Biochem. Zool. 2005, 78, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Res. | Df.diff | Dev Pr (>Dev) |
|---|---|---|---|
| (Intercept) | 62 | ||
| Environment type | 59 | 3102.6 | 0.001 *** |
| Variable | Res. | Df.diff | Dev Pr (>Dev) |
|---|---|---|---|
| (Intercept) | 62 | ||
| Environment | 59 | 3102.56 | 0.002 ** |
| Luminosity | 58 | 126.20 | 0.187 |
| Litterfall | 57 | 123.23 | 0.436 |
| Moisture | 56 | 117.29 | 0.771 |
| Temperature | 55 | 147.00 | 0.013 * |
| APU | 49 | 6137.59 | 0.006 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, F.O.d.N.; dos Santos, G.G.A.; Serra, A.B.; Miranda, C.L.; Araújo, G.d.S.; Ruz, E.J.H. Composition of the Anuran Community in a Forest Management Area in Southeastern Amazonia. Land 2023, 12, 1437. https://doi.org/10.3390/land12071437
Pereira FOdN, dos Santos GGA, Serra AB, Miranda CL, Araújo GdS, Ruz EJH. Composition of the Anuran Community in a Forest Management Area in Southeastern Amazonia. Land. 2023; 12(7):1437. https://doi.org/10.3390/land12071437
Chicago/Turabian StylePereira, Fabrício Otávio do Nascimento, Graciliano Galdino Alves dos Santos, Anderson Borges Serra, Cleuton Lima Miranda, Guilherme da Silva Araújo, and Emil José Hernández Ruz. 2023. "Composition of the Anuran Community in a Forest Management Area in Southeastern Amazonia" Land 12, no. 7: 1437. https://doi.org/10.3390/land12071437