1. Introduction
Sclerophyllous low scrub formations are common habitats at low and middle altitudes in the eastern Mediterranean and Anatolian areas. They are sub-habitats of the F7.3 European Red List of Habitats [
1]. They usually occupy dry sites with shallow limestone (calcareous) soils. According to the EU’s description of the habitats, they are located in the coastal thermo-, meso-, and supra-Mediterranean zones of the Aegean islands, in mainland Greece and the Ionian islands, and in coastal Anatolia and Crete (up to 1200 m a.s.l.) [
2]. These habitats, which are also referred to as ‘phrygana’ in Greece and garrigue in other countries [
3], are traditionally used for grazing by livestock. They consist mainly of low, thorny, dimorphic shrub species [
4]. The aforementioned species have developed an adaptation mechanism to dry thermal conditions in the Mediterranean basin by replacing winter leaves with much smaller summer ones in order to conserve water [
5,
6]. The most characteristic dominated species are
Sarcopoterium spinosum, Cistus creticus,
C. salviifolius,
Erica manipuliflora,
Genista acanthoclada, Phlomis fruticosa, Corydothymu scapitatus, and
Euphorbia acanthothamnus [
7].
Sclerophyllous scrub formations are very diverse ecosystems. They comprise some of the most species-rich plant communities in the Mediterranean basin and provide many goods and services to humans. Regarding regulating and supporting services [
8], they contribute to the regulation of soil quality and protection, carbon sequestration, and provide a habitat for wildlife. Concerning provision services, they provide medicinal and aromatic plants, chemical extracts, and food as the majority of dominated scrubs and many of the understory herbs have medicinal and aromatic properties, e.g., honey from wild herbs, as well as forage for livestock and wildlife [
9].
These ecosystems in the Aegean islands have been traditionally used for grazing by livestock. Despite the fact that the main use of these ecosystems is grazing, the dominated phryganic species are unpalatable and/or less desirable for grazing. This means that the main source of forage is the herbaceous species under or among the shrubs. Thus, an increase in the density of the phryganic species will result in a decrease in the availability and quality of the forage. In addition to the reduction of forage availability, this thickening would also result in a decrease in species richness and floristic diversity. Shepherds in these areas know this and use fire as a tool to increase the forage quantity and quality [
7]. Additionally, they are among the major fire-prone biomes in the world [
10]. Fire in this biome is an essential ecological process and beneficial for the ecosystem’s function [
11]. In this respect, grazing and fire are key factors that have interacted with and shaped the structure and function of plant communities in the phryganic ecosystems in the Aegean islands.
However, grazing has drastically decreased in the past few decades in these areas, mainly due to changes in land uses and the increase in tourism. This has led to changes in landscapes and the environment, changes in vegetation composition and structure, decreases in forage quantity and quality, increases in woody vegetation, and the loss of biodiversity, endangering the provision of key ecosystem services [
12,
13]. Furthermore, the increase in woody vegetation may contribute to an increase in the risk and/or the frequency of wildfires. Fire-prone biomes have a characteristic historical range of variability in frequency, severity, and patchiness of fires [
14]. Any change in this historical range due to human intervention can alter the ecosystem’s response to fire [
15].
The effect of fire on ecosystem services has been studied mainly in forest ecosystems and for services related to soil stability and fertility [
15]. The general view is that fire is a natural disaster and has a negative effect on ecosystem services. On the other hand, these ecosystems can remain stable under grazing and burning [
16]. Particularly for post-fire grazing management, the question that arises is: when can it be applied to burned areas? The implementation of grazing management in burned areas directly depends on the establishment and structure of post-fire vegetation. Livestock grazing in burned areas before vegetation is well established could lead to ecosystem degradation. On the other hand, the thickening of vegetation before livestock grazing could make the ecosystem vulnerable to a new fire. However, research about the effect of fire on ecosystems adapted to fire, such as the sclerophyllous low scrub formations, is limited. The sclerophyllous low scrub formation of Aegean islands is assessed at the Least Concern status [
2] based on Indicators of quality of the European Red List of Habitats [
1]. This is mainly because of the extensive distribution of this habitat in the Eastern Mediterranean, which has not decreased in recent years. However, these ecosystems are highly affected by human activities such as grazing, fire, and cultivation abandonment. Therefore, further research focusing on the quality characteristics of these ecosystems is needed in order to detect the role of human activities in plant communities and how they affect the provision of their services. In this respect, the aim of the present study was to detect the effects of wildfire on species composition, floristic diversity, forage quality, and rangeland health indices related to ecosystem stability and function in three thermo-Mediterranean vegetation types (1) low formations of
Sarcopoterium spinosum, (2) low formations of
Cistus creticus and (3) low formations of
Cistus creticus in abandoned terraces. This information could be a tool for managers to implement grazing plans in the burned areas of these ecosystems.
4. Discussion
It is well substantiated that wildfires are a common phenomenon in the Mediterranean rangeland ecosystems, and they alter the structure and dynamics of plant communities [
41,
42]. Many plant species have adapted to fire using two basic mechanisms: (a) by resprouting from alive plants after the fire and (b) by recruiting from seeds [
43,
44]. As a result, the vegetation in these ecosystems has the ability to recover a few years after the fire [
45].
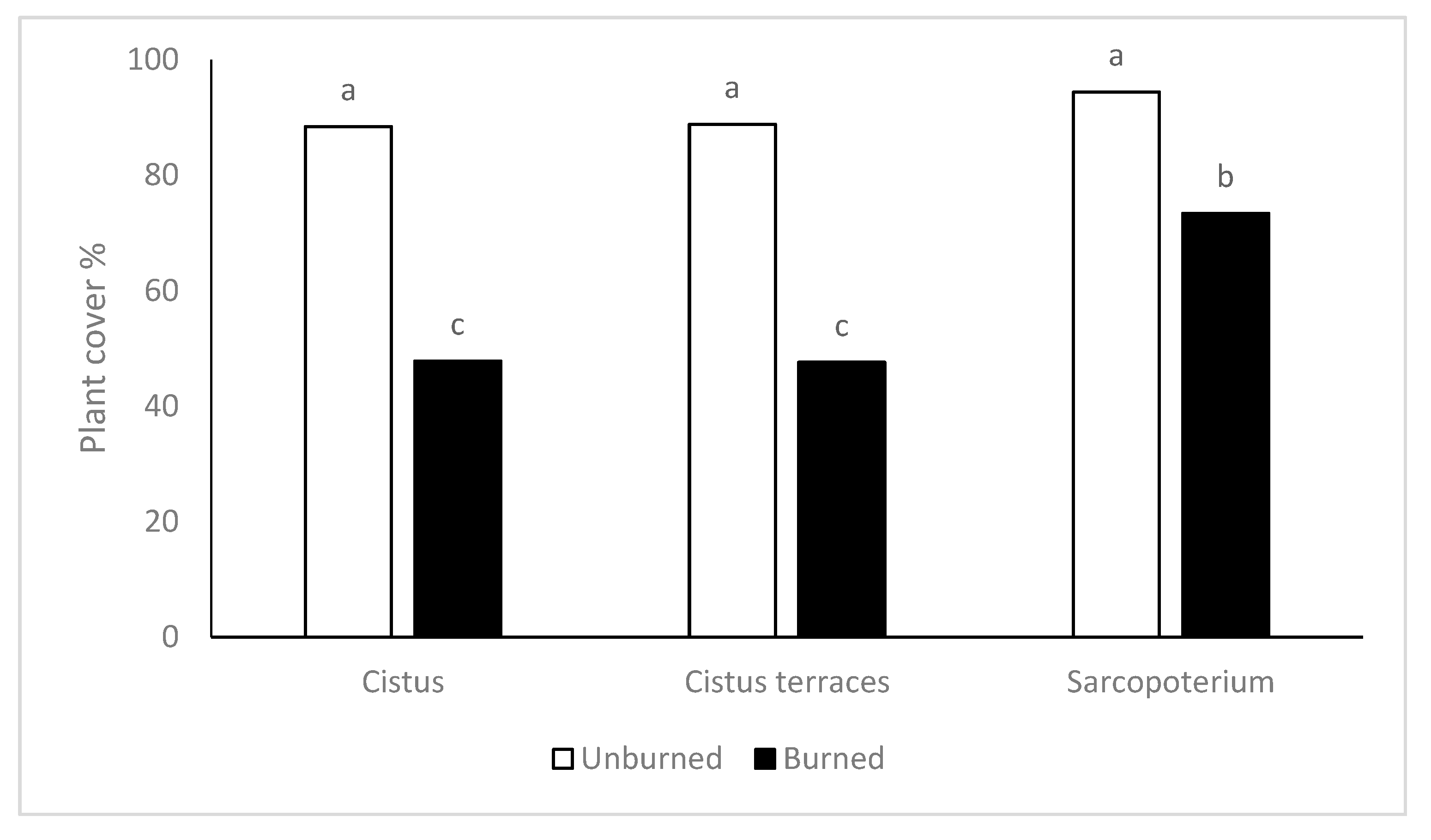
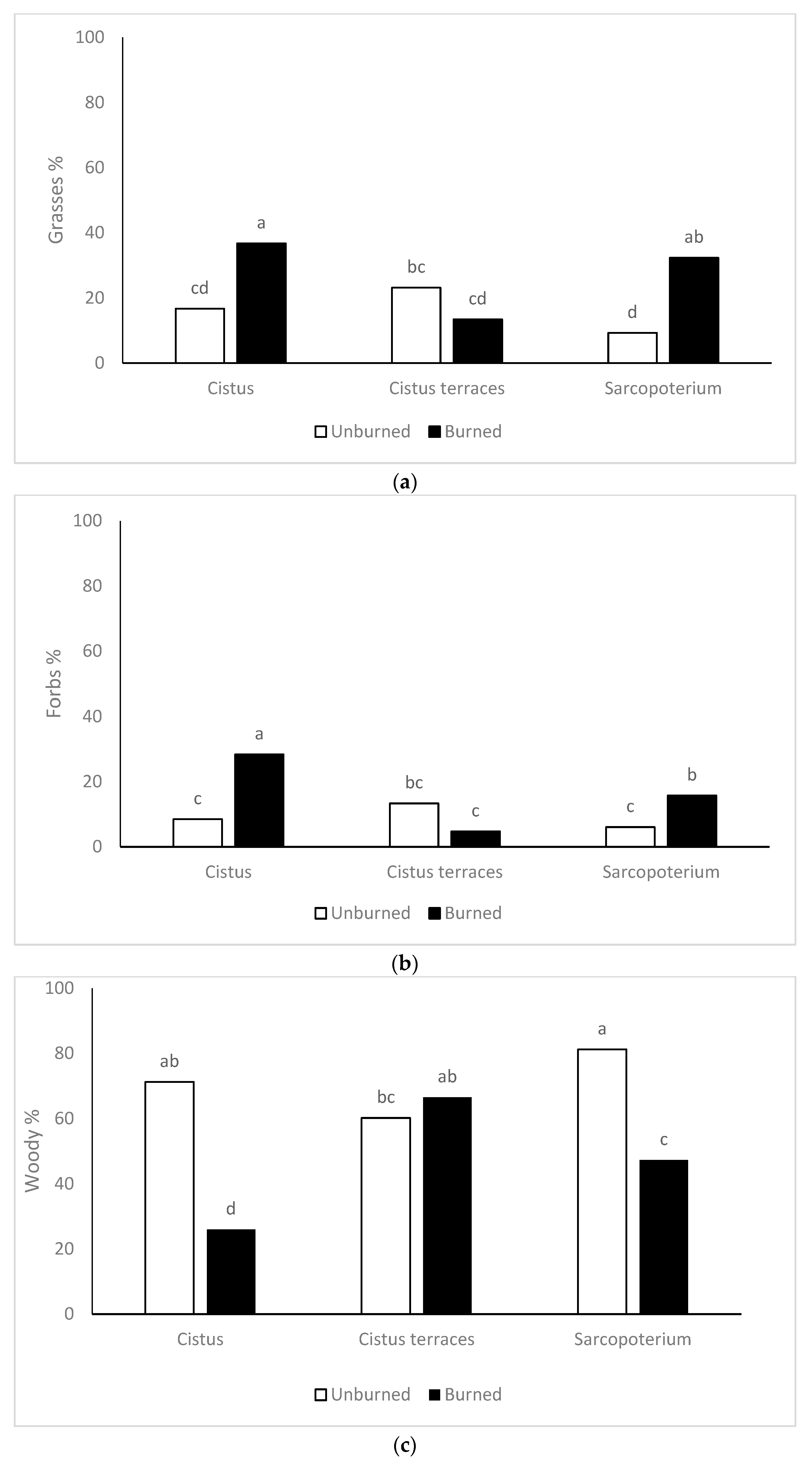
According to the results of the present study, there were no differences in plant cover among the vegetation types of the unburned sites. On the other hand, fire reduced plant cover in all the vegetation types, though in different degrees. Plant cover in the burned formations of
S. spinosum was higher than those recorded in the
C. creticus formations and the terraces with
C. creticus. According to Kazanis and Arianoutsou [
46], plant cover post-fire is affected by the woody species cover. Both woody species are post-fire pioneer plants [
47,
48].
Cistus species are force seeders [
49,
50].
C. ladanifer has been reported as a species that dominates in burnt areas [
49]. It can recover faster even than resprouted species [
51], besides the fact that vegetative resprouting has advantages over seed germination in burned environments [
52].
S. spinosum can recover by both resprouting and seed germination [
53]. The aggressive regrowth and competitive ability of this species resulted in a higher degree of plant cover in the formations of
S. spinosum.
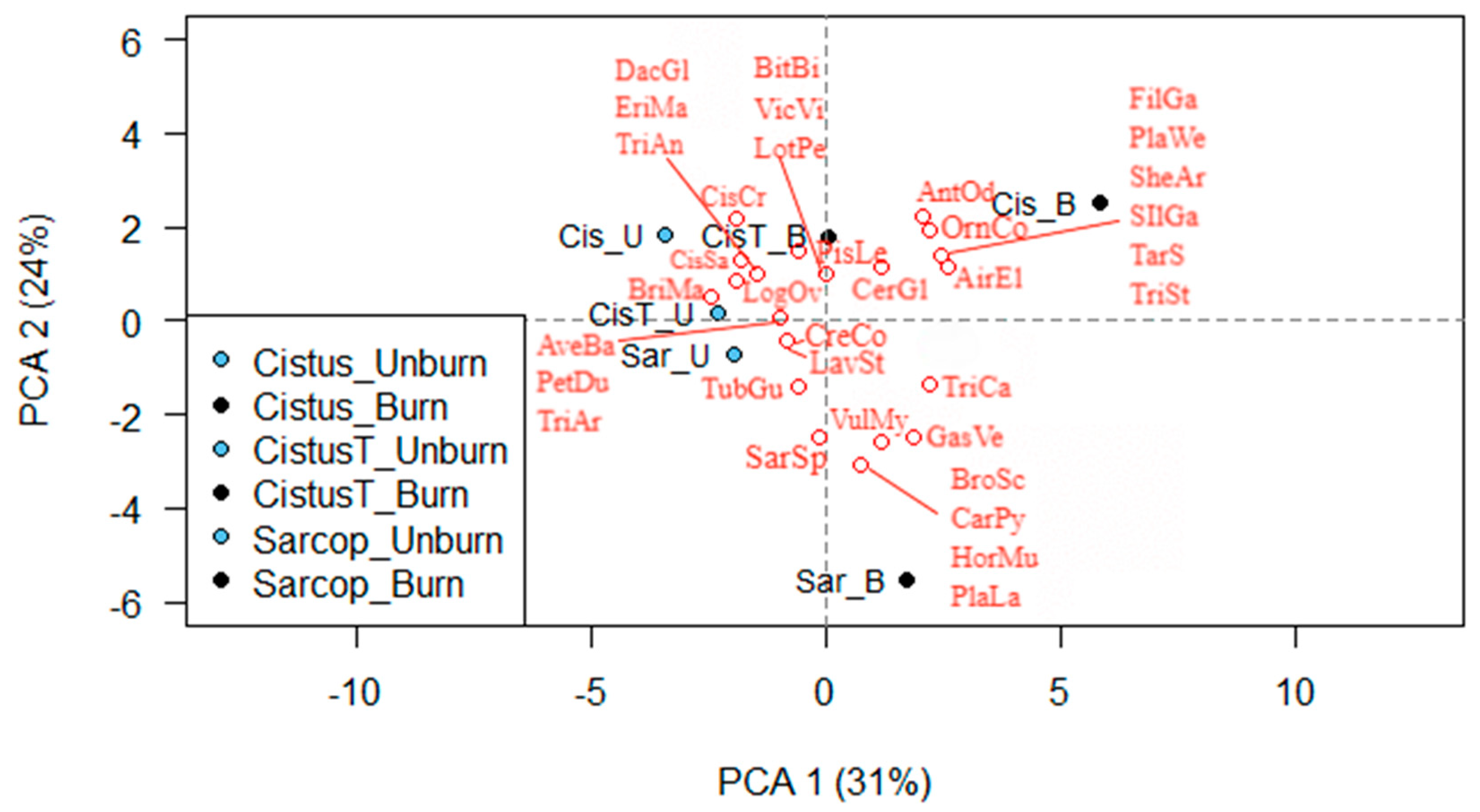
The floristic composition was differentiated in the burned sites in relation to the adjacent unburned ones but to a different degree across the three vegetation types. The pre-fire floristic composition was dominated by woody species (around 70%), followed by grasses, forbs, and legumes in all vegetation types. In the post-fire floristic composition, the abundance of woody species decreased, while the abundance of grasses, forbs, and legumes increased in the low formations of C. creticus and S. spinosum. Inversely, in the terraces of Cistus creticus, there were no differences in the abundance of woody species, grasses, and forbs between burned and unburned sites. This resulted in the highest differentiation in terms of the Jaccard similarity index being recorded between burned and unburned sites of C. creticus formation and the lowest in the terraces of C. creticus.
Fire generally contributes to the decline of woody species [
54,
55] in rangelands. In many cases, prescribed burning is used as a management tool to limit shrub encroachment in grasslands. The reduction of woody species leads to a limitation of competition, which favors the establishment of other functional groups [
56,
57]. The recovery of woody species after the fire depends on their regeneration capacity [
58]. As mentioned above, the regeneration capacity of both species (
C. creticus and
S. spinosum) is high and has contributed to their quick recovery. Notably, the percentage of
S. spinosum in the composition of the vegetation reached about 50% just one year after the fire. In this respect, Papanastasis [
59] reported the full recovery of
S. spinosum three years after a fire. Furthermore, it should be noted that the faster recovery of
Cistus creticus in the terraces compared to the other sites. The abundance of
Cistus creticus in terraces was similar before and after the fire. This can be attributed to the specific micro-environment of terraces (soil, light, temperature) that probably favors the germination of the seeds and the establishment of seedlings [
60]. Terraces are common in hilly areas and have been built in order to conserve soil and water as well as to increase the arable fields [
61]. Finally, the percentage of legumes increased in post-fire vegetation in all the vegetation types. Actually, the percentage of legumes in burned sites was three times more compared to the unburned ones. Probably, fire contributes to the cracking of their hard-coated seeds and accelerates their germination [
54,
62].
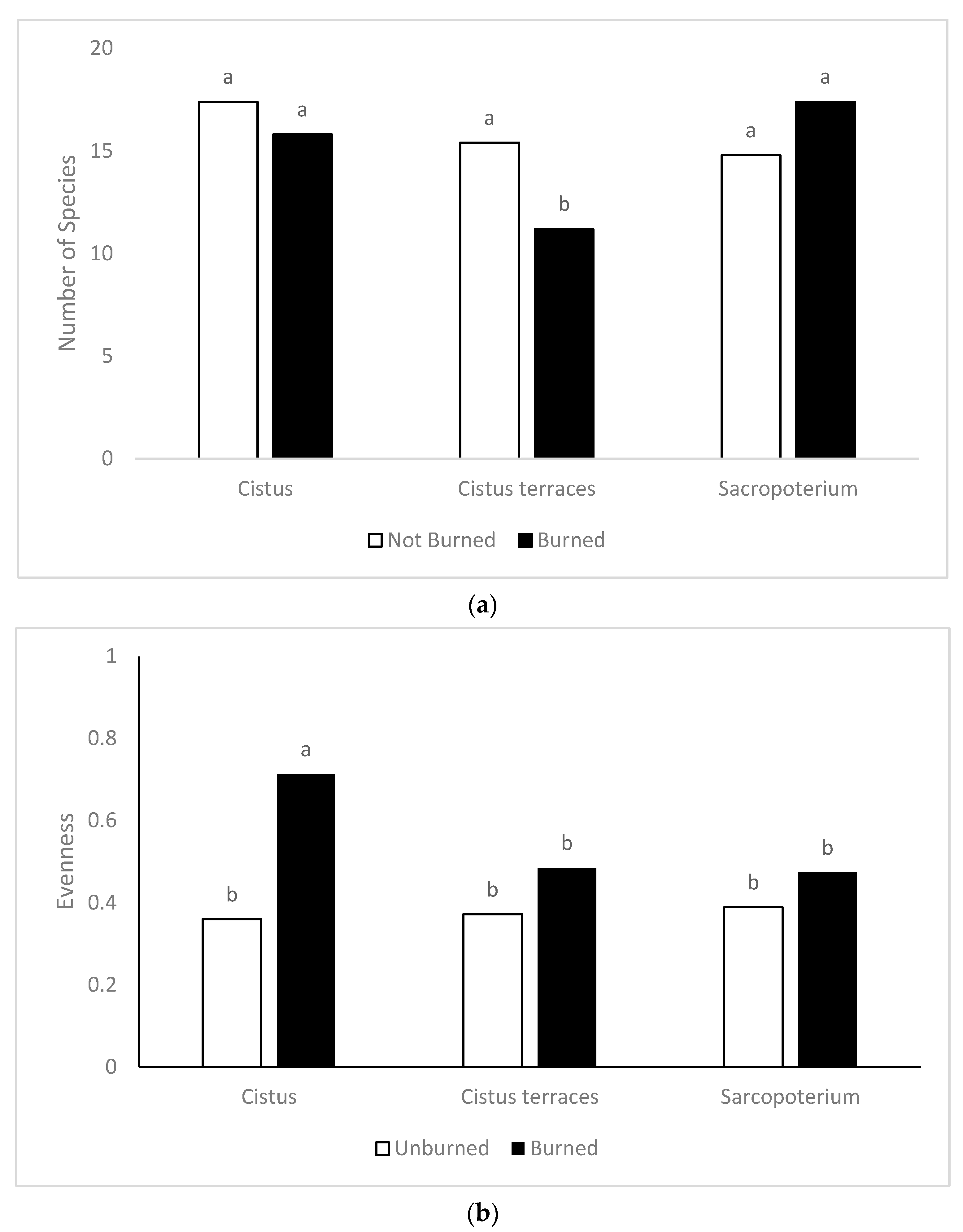
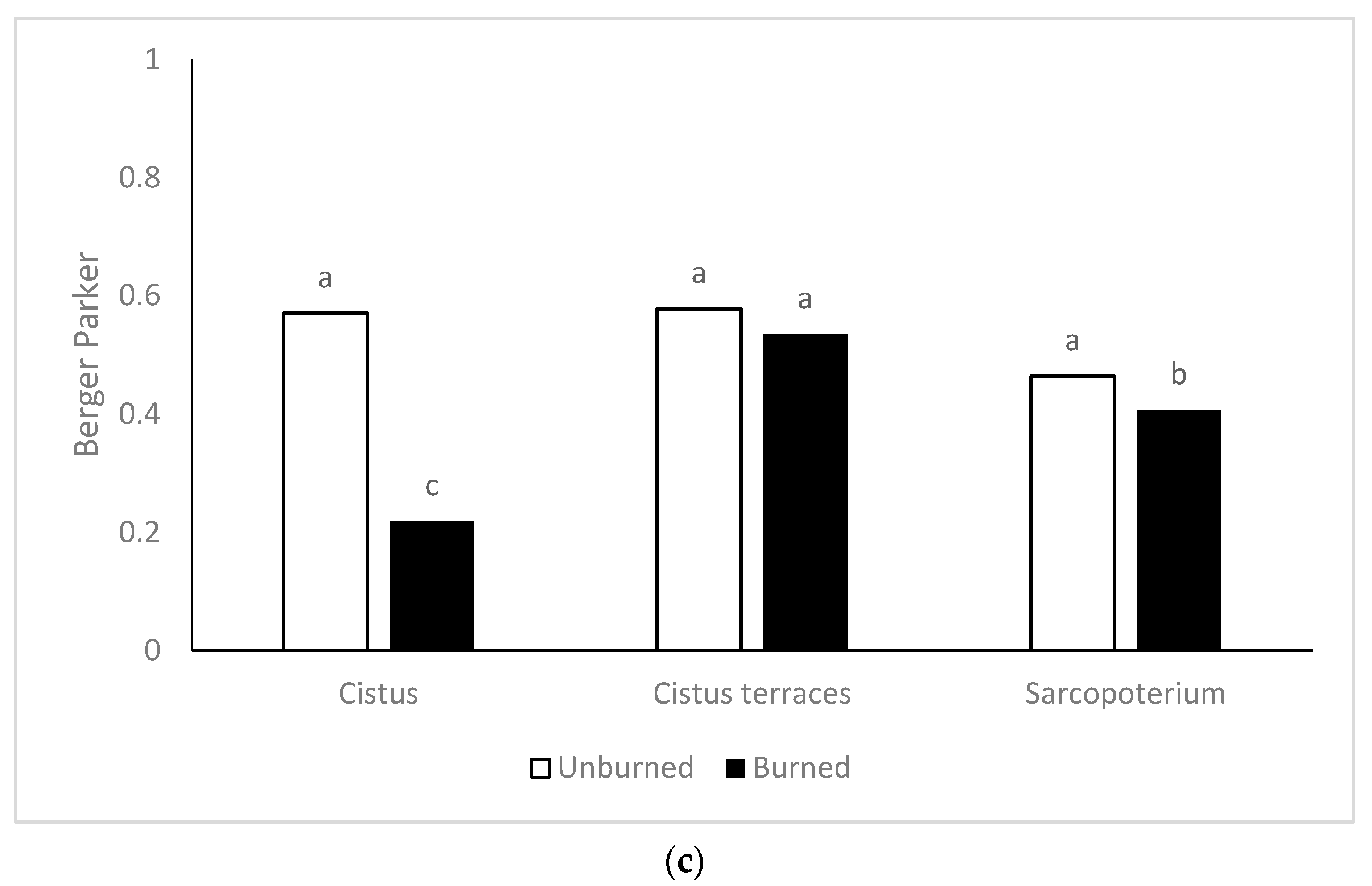
The floristic diversity was higher in the burned sites compared to the unburned ones. There are many reports about the positive effect of fire on floristic diversity in rangelands [
63,
64,
65]. In particular, for fire-prone ecosystems such as the phryganic, fire has been proposed as a major driver of their diversity [
66]. This positive effect mainly contributed to the decrease of the competitive woody species and the increase of the other functional groups.
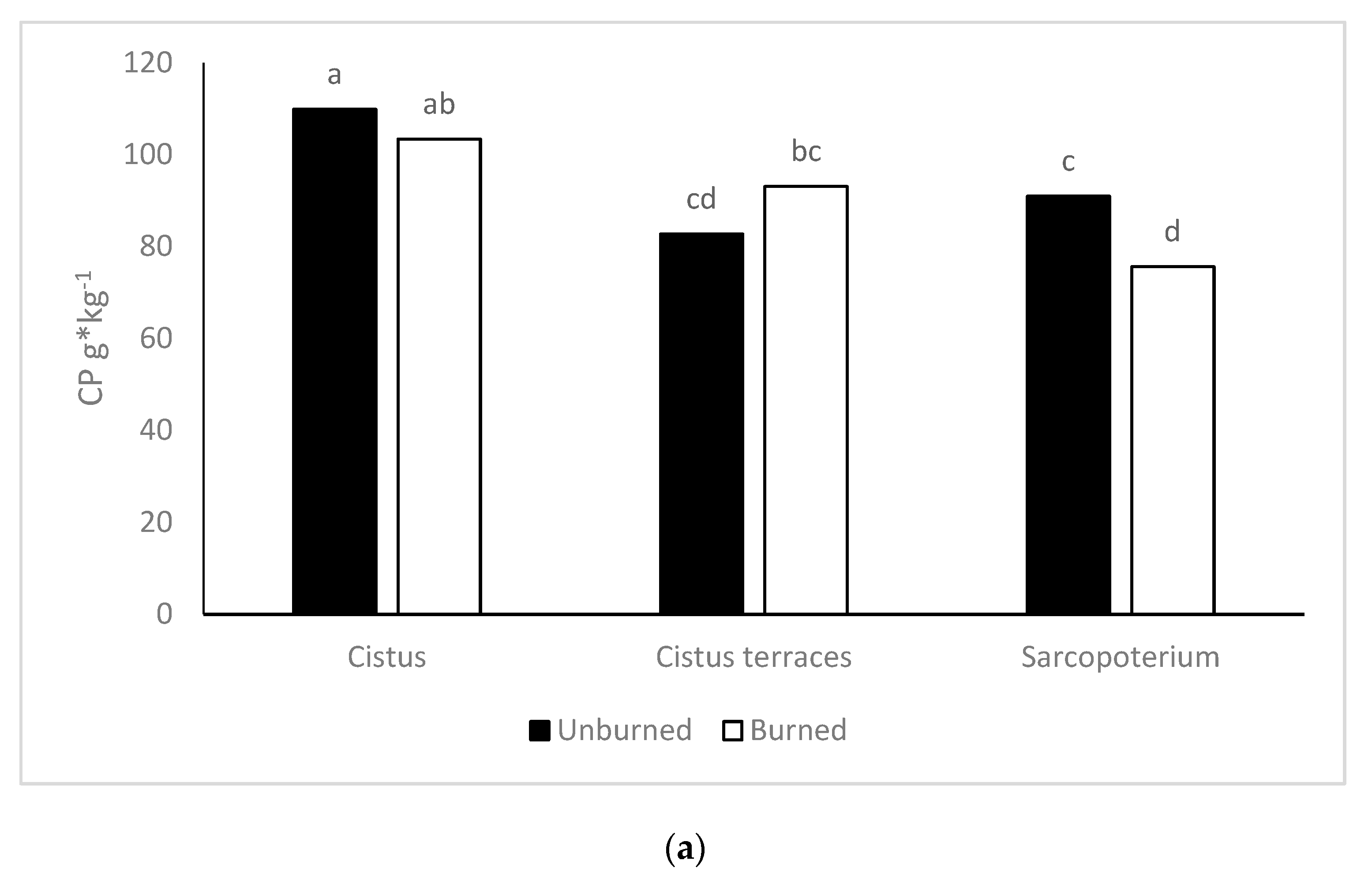
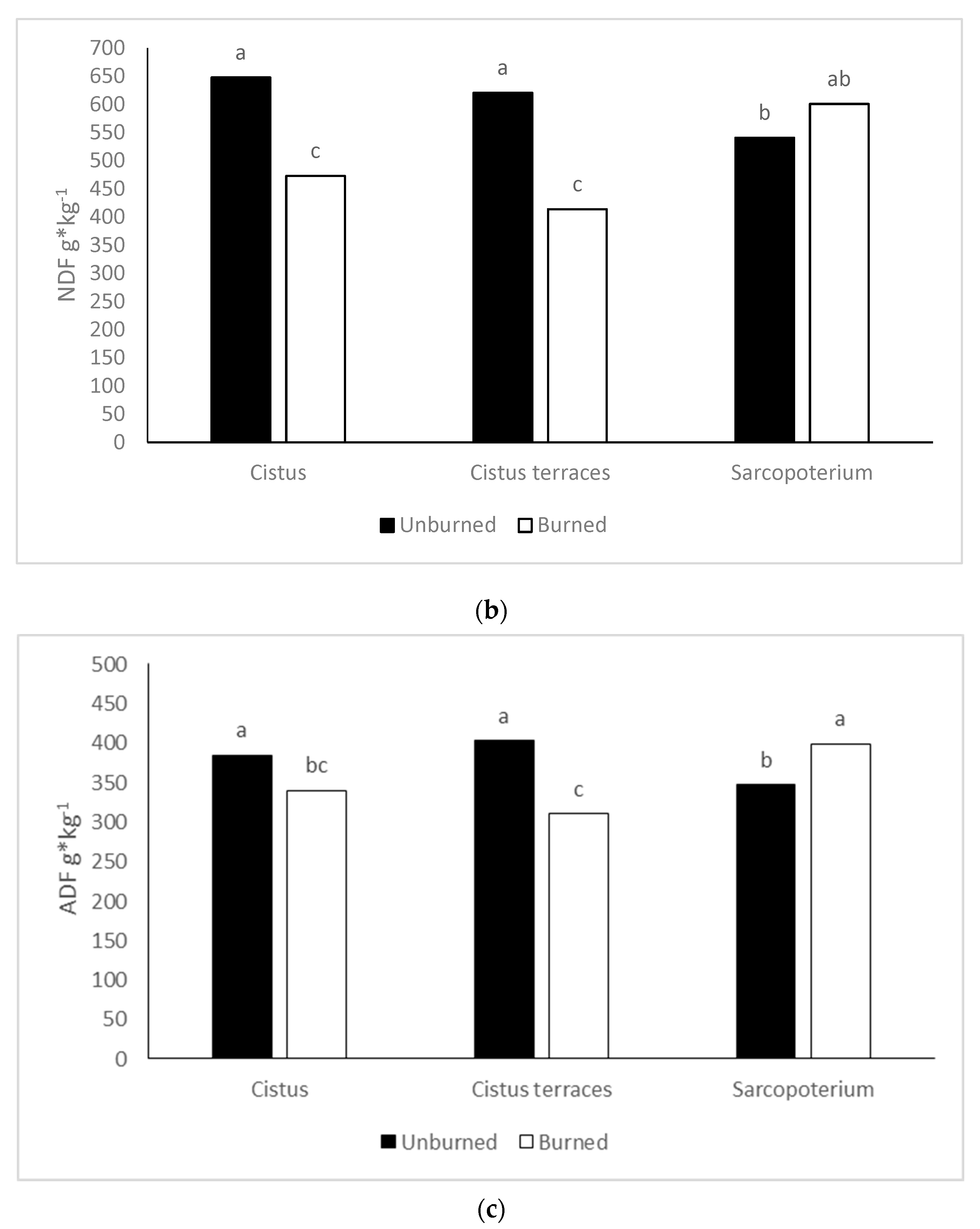
The CP content did not significantly differ between burned and unburned areas. This finding was unexpected as forage in burned areas has higher crude protein than forage in unburned ones [
67]. The absence of a response can be an indication that livestock did not graze in these areas in time to benefit from the initial greening up in the burned locations [
68]. Another potential explanation is that the effect of time (after one year) evened out the difference in CP content. According to Thapa [
69], in research carried out in grasslands in Nepal, there was no difference in the CP content of the post-fire regrowth forage after four months. Although there was a decrease in fiber content, ADL probably increased due to an increase in legumes in the burned areas. This is due to the possible higher content of condensed tannins, which interfere with ADL [
70].
On the other hand, the vegetation type (across fire) had different CP content for the
Cistus vegetation types. These variations may have developed because of their different growing environment. Temel and Tan [
71] reported similar results for the CP content of
C. creticus, although they have estimated lower NDF and ADF content for the same species in comparison to this study. According to Gokkus [
72],
S. spinosum had lower CP content (5.37%) in spring compared to our result but almost double ADL content (15.98%). Generally, the two dominant species,
C. creticus and
S. spinosum, are not as preferable with low nutritive value. However, both have been affected by wildfire, which had a positive effect on the nutritive value in terms of fiber and CP content in the
C. creticus vegetation type but had an adverse effect on the
S. spinosum vegetation type [
73].
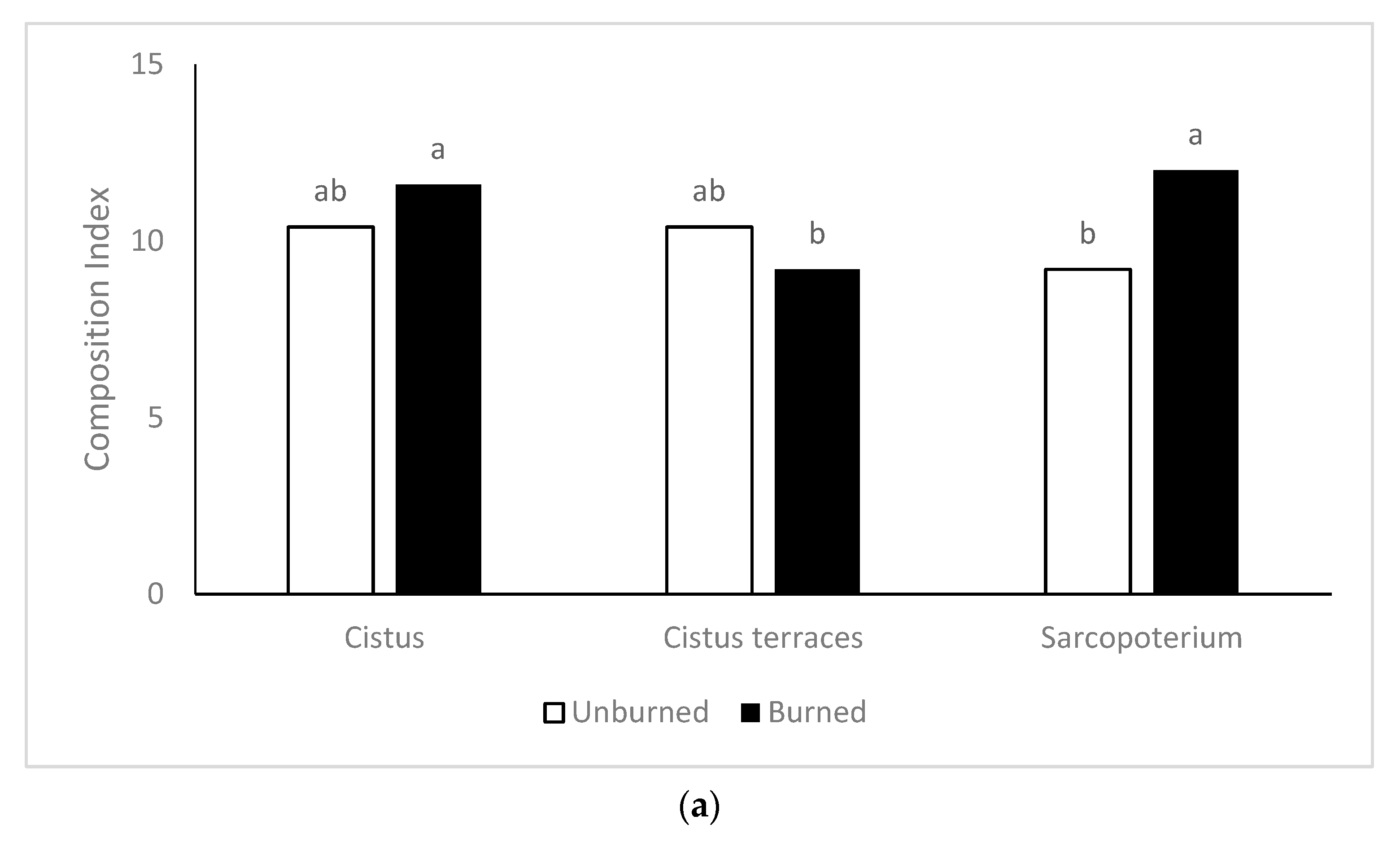
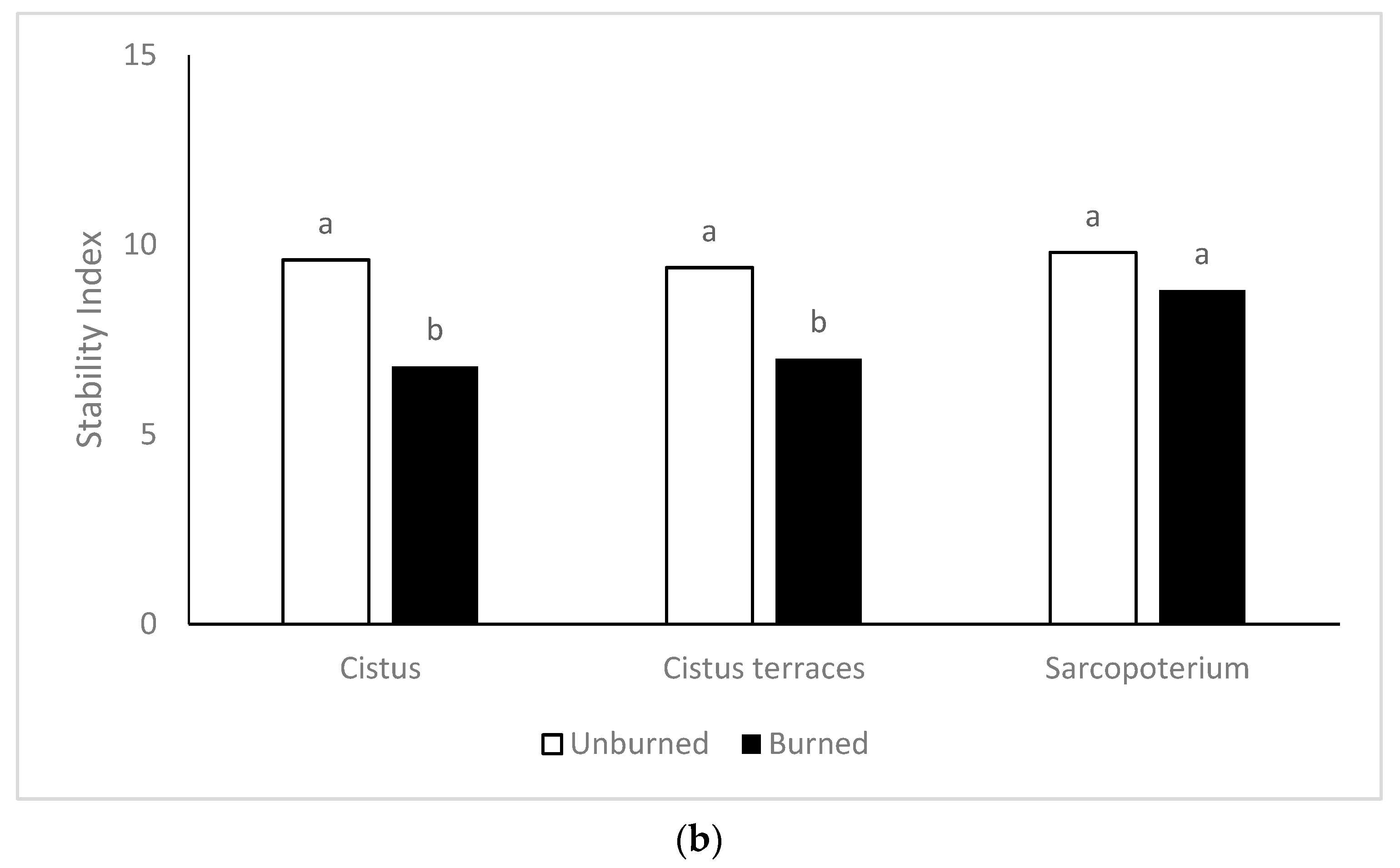
The landscape function and stability were negatively affected by burning. The increased soil erosion and the reduced plant cover recorded in the burned rangeland sites constitute the main reason for the reduced stability index, mainly in the
C. creticus formations and the terraces with
C. creticus. Increased risk for soil erosion in burned similar vegetation types has been reported [
74]. Although legumes’ percentage increased in the burned sites, the reduced herbage production and the increased soil erosion reduced the function index. In contrast, burning benefited the composition index, especially in the formations of
S. spinosum, because of the lower woody species cover and the increased floristic diversity. It has to be noted that formations of
S. spinosum were favored by burning more than the other vegetation types as the composition index was increased, while the stability index did not affect one year after the fire.
Sarcopoterium spinosum is an unpalatable species [
75] and is not consumed by livestock except early in spring, while
Cistus creticus is browsed by goats mainly in autumn [
76]. Therefore, prescribed burning has been used to combat dense
Sarcopoterium spinosum communities and improve rangeland vegetation [
45]. The results of the present study confirm that burning can improve the landscape in this vegetation type, as well as the idea that these ecosystems can remain stable when not dense [
16].

 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}