

Spatial Distribution Pattern and Influencing Factors of Above-Ground Biomass and Species Diversity of Grassland in the Altay Forest Area

Abstract
:1. Introduction
2. Materials and Methods
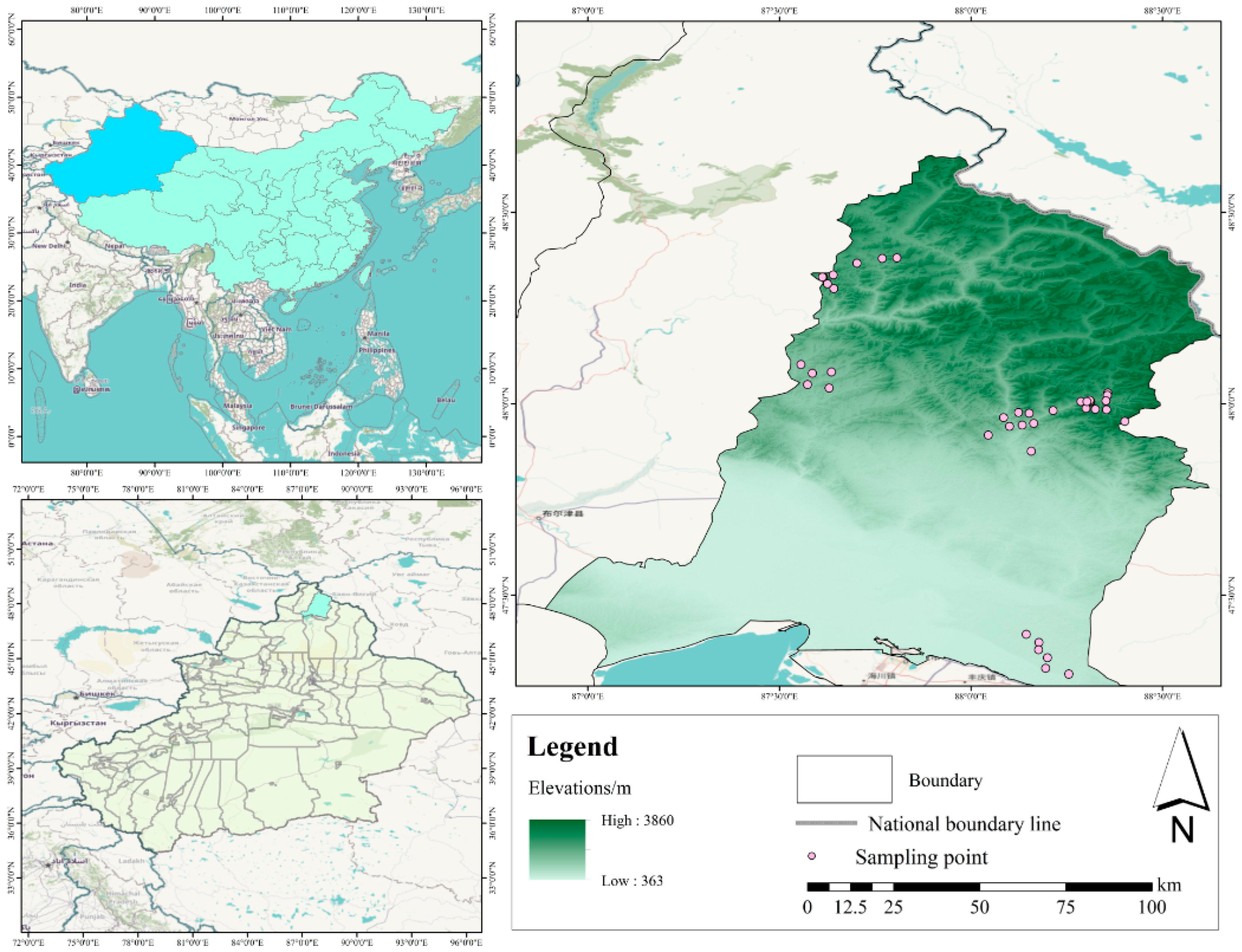
2.1. Study Area
2.2. Design of Experiments
2.3. Calculation of Species Diversity
2.4. Data Calculation
3. Results
3.1. Spatial Pattern and Influencing Factors of Grassland Above-Ground Biomass in Altay Forest Region
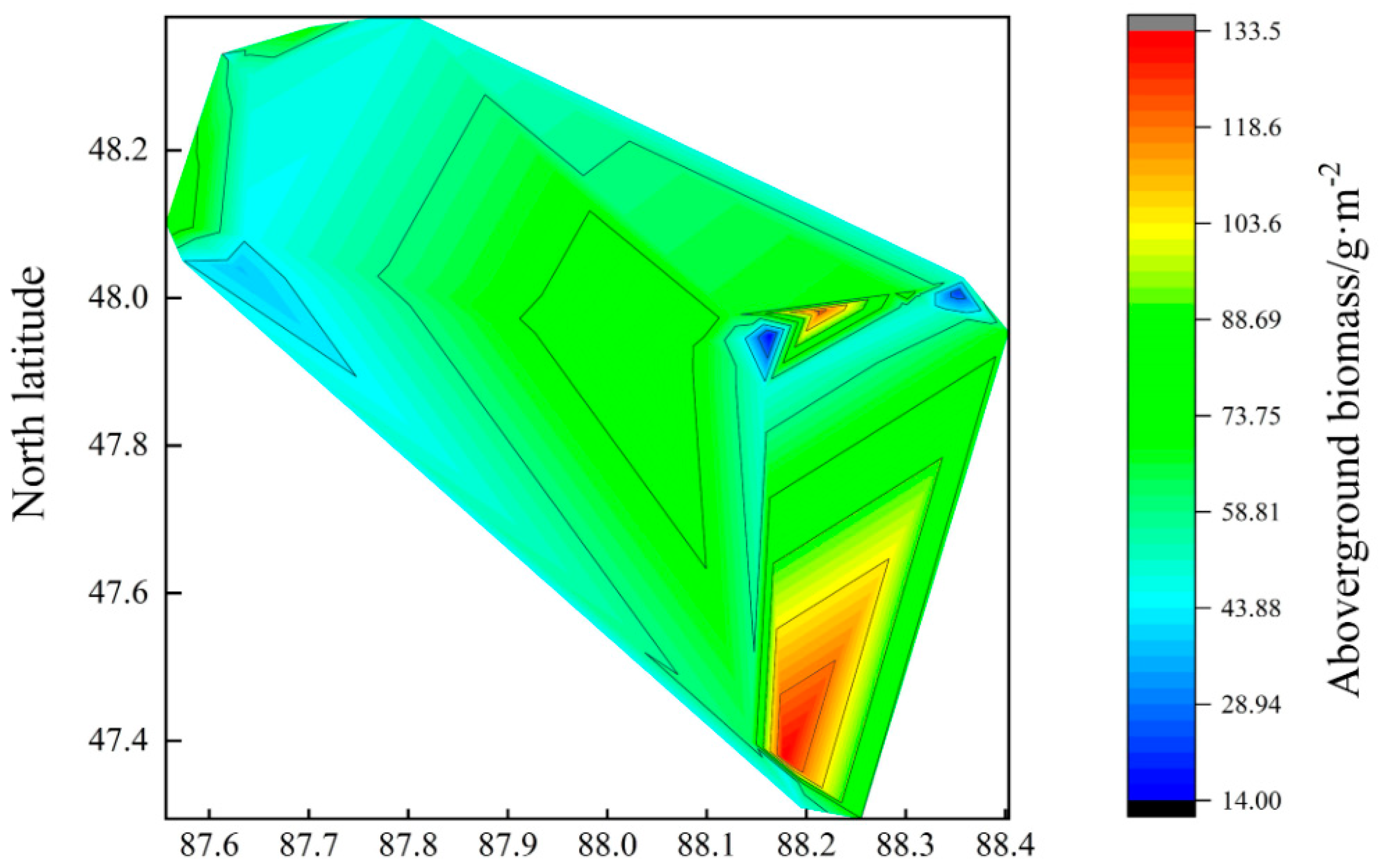
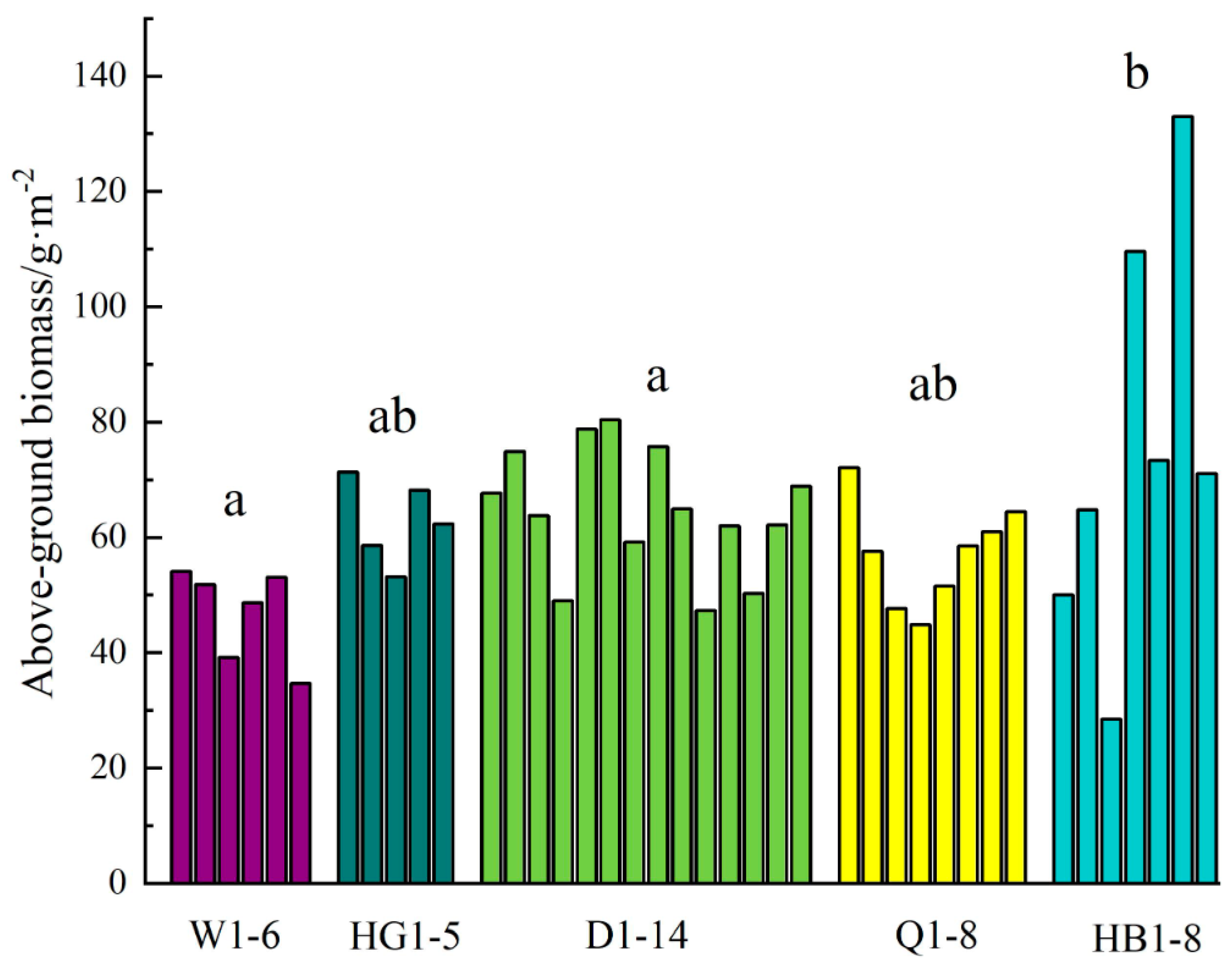
3.1.1. Spatial Pattern of Above-Ground Biomass
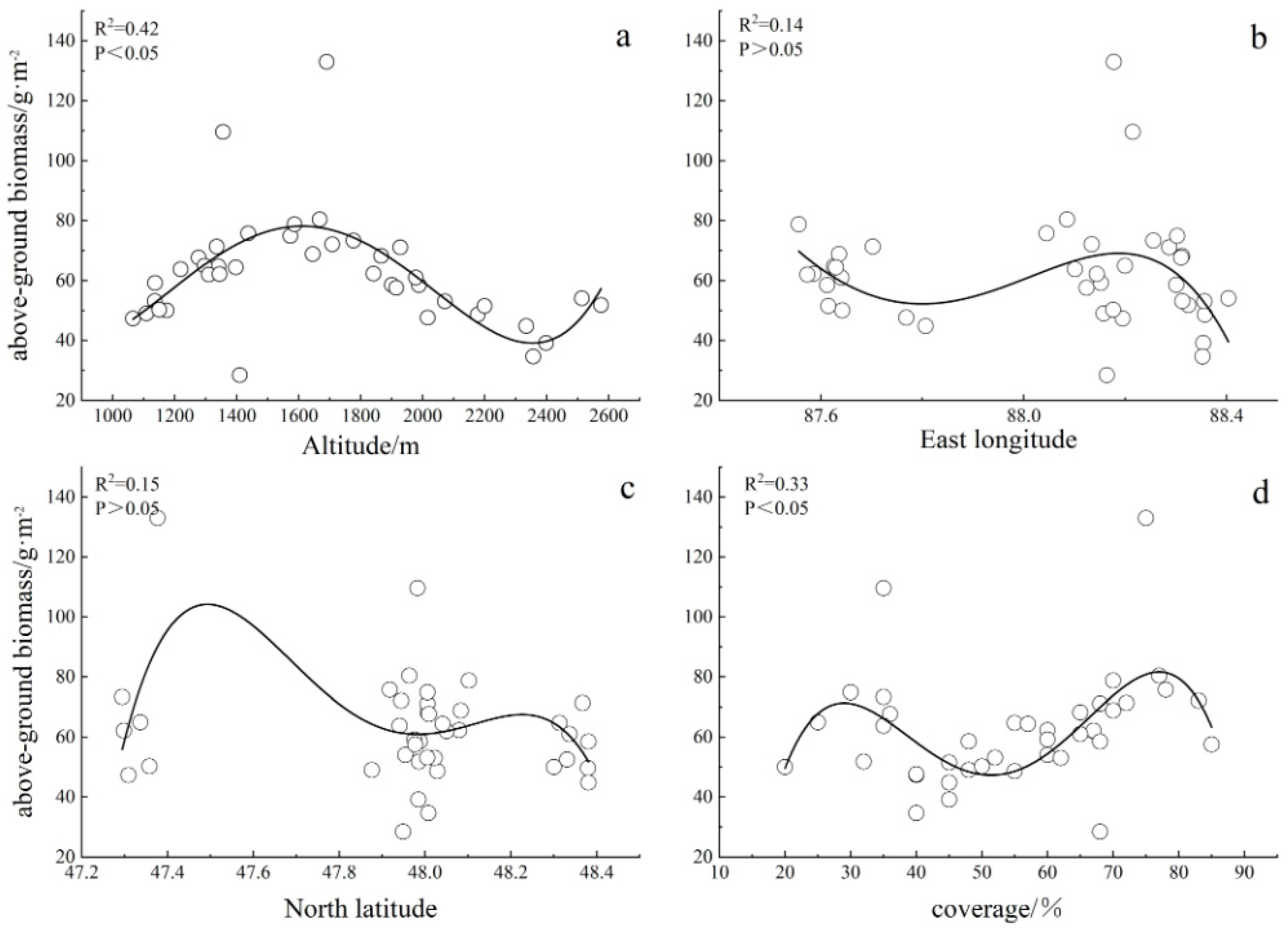
3.1.2. Influencing Factors of Spatial Pattern of Above-Ground Biomass
3.2. Spatial Pattern of Species Diversity and the Factors That Influence It
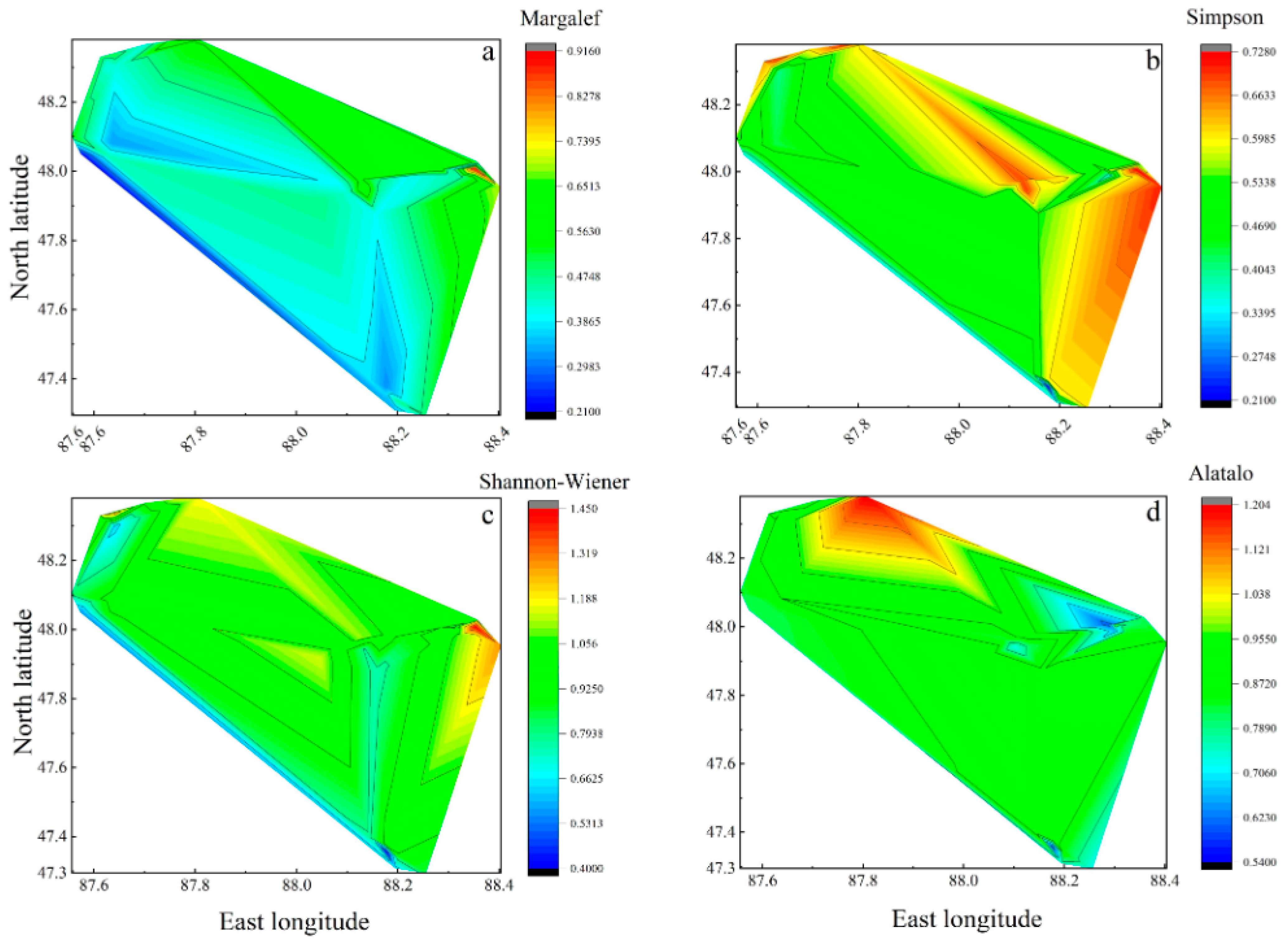
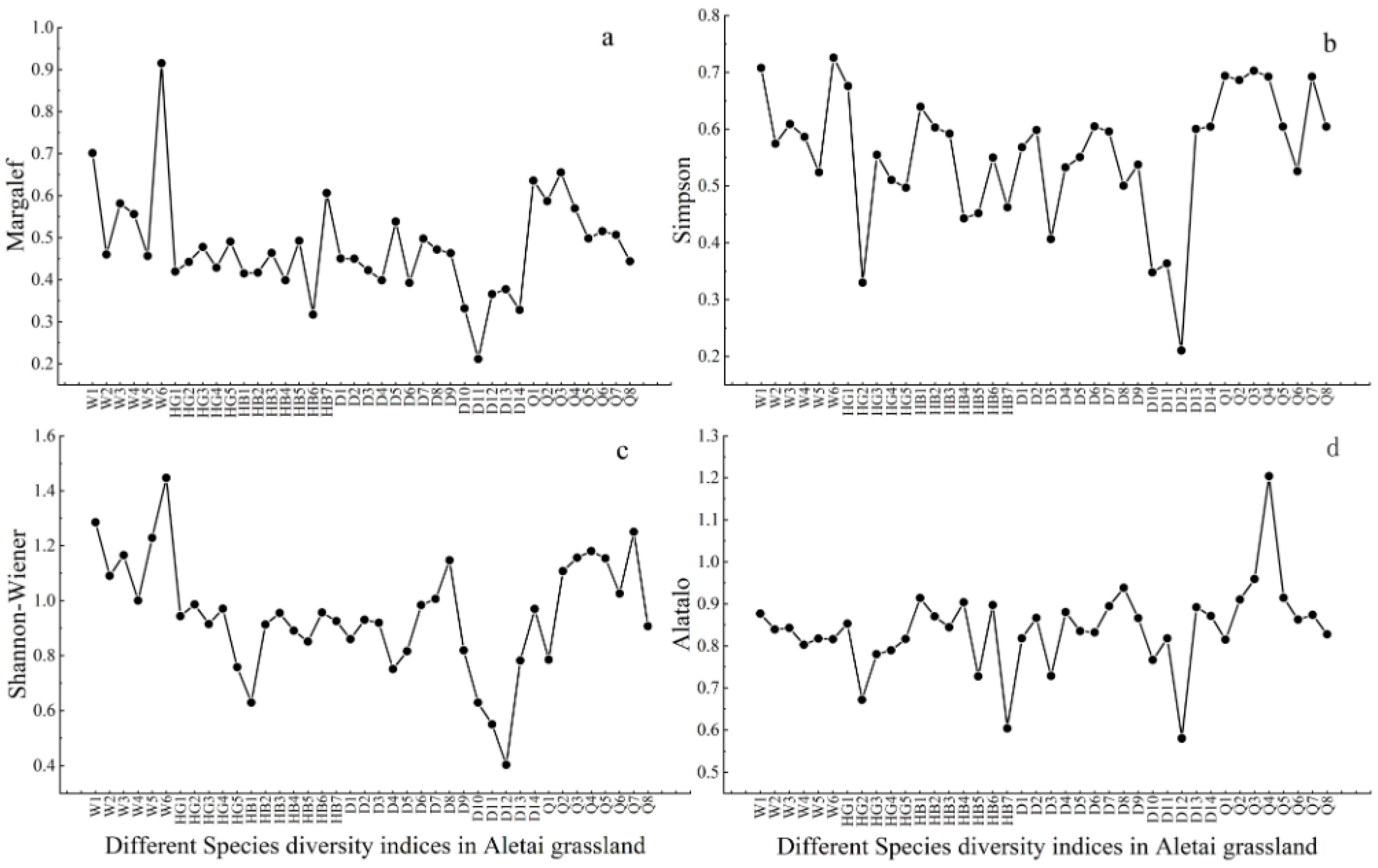
3.2.1. Spatial Pattern of Species Diversity
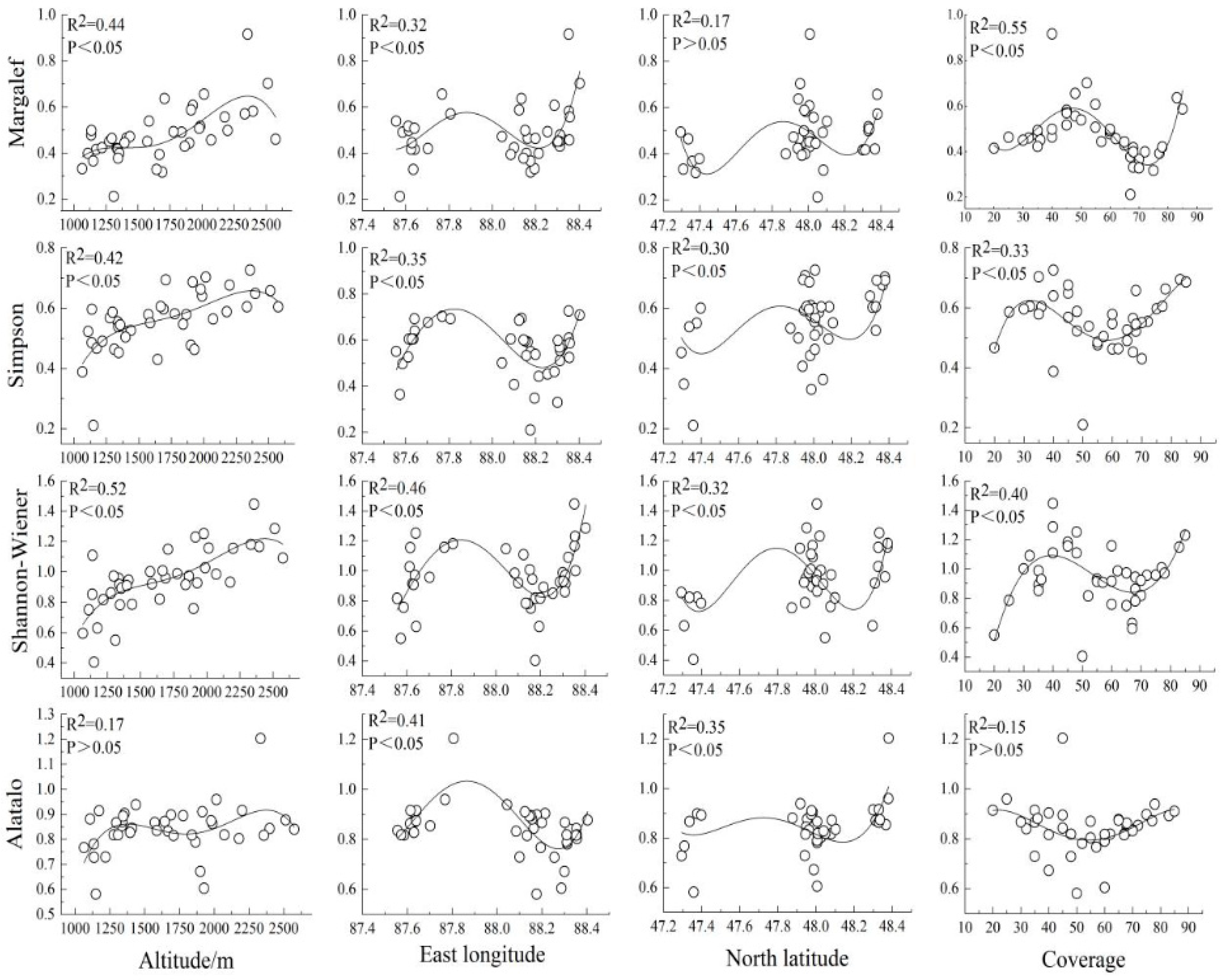
3.2.2. Influencing Factors of Spatial Pattern of Species Diversity
4. Discussion
4.1. Spatial Pattern of Above-Ground Biomass and Its Influencing Factors
4.2. Spatial Pattern of Species Diversity and Its Influencing Factors
5. Conclusions
- (1)
- The above-ground biomass of the grassland in the Altay forest area decreased gradually from northeast to southwest, and the above-ground biomass was significantly different in different plots. The average above-ground biomass was 46.93 g/m2 in the Wuxilike plot, and the highest above-ground biomass was 75.75 g/m2 in the altitude plots;
- (2)
- The above-ground biomass of grassland in the Altay forest region first increased and then decreased with the increase in altitude, showing a single peak, and the biomass was the largest at an altitude of 1400–1600 m. It showed a significant “bimodal” relationship with coverage;
- (3)
- The species diversity index of the grassland in the Altay forest area was generally high and characterized by high diversity in the northeast and low diversity in the southwest. Except for the Alatalo index, the lowest value mostly appeared in the southwest. The overall average species diversity index of the Dadonggou plot was the lowest, and the overall species diversity index of the Qiaoati plot was higher;
- (4)
- Except for the Alatalo index, which had no significant correlation with altitude and coverage, the species diversity indexes showed significant correlations with altitude. The distribution pattern of longitude and coverage was an “S” type. There is a significant “W” distribution trend with latitude. Altitude, latitude, and longitude were the main environmental factors affecting the spatial pattern of above-ground biomass and species diversity in the Altay grassland.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, H.Z.; Fan, J.W.; Liu, S. A Comprehensive Analysis of Difference in Carbon Stock Estimation in the Grasslands of China. Acta Agrestia Sin. 2017, 25, 905–913. [Google Scholar]
- Scurlock, J.M.O.; Johnson, K.; Olson, R.J. Estimating net primary productivity from grassland biomass dynamics measurements. Glob. Chang. Biol. 2002, 8, 736–753. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.H.; Fang, J.Y.; Ma, W.H.; Guo, D.L.; Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Glob. Ecol. Biogeogr. 2010, 19, 268–277. [Google Scholar] [CrossRef]
- Wu, H.B.; Shui, H.W.; Hu, G.Z.; Wang, X.X.; Gan, Z.Z.B.; Yan, J.; He, S.C.; Xie, W.D.; Gao, Q.Z. Species diversity and biomass distribution patterns of alpine grassland along an elevation gradient in the Northern Tibetan Plateau. Ecol. Environ. Sci. 2019, 28, 1071–1079. [Google Scholar]
- Jia, W.X.; Liu, Y.R.; Zhang, Y.S.; Cao, W.X. Species diversity and biomass of grassland steppe in Qilian Mountains and their relationships with climate factors. Arid Zone Res. 2015, 32, 1167–1172. [Google Scholar]
- Zhang, Y.; Zhao, J.L.; Xin, X.P.; Wang, M.; Pan, F.J.; Yan, R.R.; Li, L.H. Effects of stocking rate on the interannual patterns of ecosystem biomass and soil nitrogen mineralization in a meadow steppe of northeast China. Plant Soil 2021, 473, 9–31. [Google Scholar] [CrossRef]
- He, M.Y.; Wang, Y.X.; Peng, Z.C.; Chang, S.H.; Saman, B.; Liu, Y.J.; Hou, F.J. The spatial pattern of aboveground biomass and species richness in the grassland of Qilian Mountain grassland. Pratacult. Sci. 2020, 37, 2012–2021. [Google Scholar]
- Dai, L.C.; Ke, X.; Zhang, F.W.; Du, Y.G.; Li, Y.K.; Guo, X.W.; Li, Q.; Lin, N.L.; Ma, J.J.; Cao, G.M. Allocation patterns of above- and belowground biomass and its response to meteorological factors on an alpine in Qinghai-Tibet Plateau. Acta Ecol. Sin. 2019, 39, 486–493. [Google Scholar]
- Piao, S.L.; Fang, J.Y.; He, Y.S.; Xiao, Y. Grassland vegetation biomass and its spatial distribution pattern in China. J. Plant Ecol. 2004, 28, 491–498. [Google Scholar]
- Zhang, Y.; Hou, L.L.; Yan, R.R.; Xin, X.P. Effects of grazing on plant community characteristics and Nutrient quality of Herbage in a Meadow Steppe. Sci. Agric. Sin. 2020, 53, 2550–2561. [Google Scholar]
- Wu, K.; Xu, W.X.; Yang, W.K. Impact of Rainfall Changes on Plant Community in Junggar Desert. Res. Soil Water Conserv. 2019, 26, 100–106. [Google Scholar]
- Lanta, V.; Leps, J. Effects of species and functional group richness on production in two fertility environments: An experiment with communities of perennial plants. Acta Oecol. 2007, 32, 93–103. [Google Scholar] [CrossRef]
- Dai, M.M.; Yang, K.; Li, S.; Wang, X.D.; Song, Z.B. Fish diversity and resource status in the Nanguang River, a tributary of upper Yangtze River. Biodivers. Sci. 2019, 27, 1081–1089. [Google Scholar]
- Guo, J.X.; Ye, M.; Yin, X.K.; Zhang, K.L. Biomass and Diversity of Grassland Community in the Two-river Source Pastoral Areas of Altai Mountains. Acta Bot. Boreal.-Occident. Sin. 2022, 42, 1936–1944. [Google Scholar]
- Chen, J.J.; Chang, S.H.; Wang, Z.F.; Peng, Z.C.; Cheng, Y.X.; Chen, X.J.; Hou, F.J. The relationship between plant species diversity and above-ground biomass in a typical steppe in Abaga Banner. Pratacult. Sci. 2018, 35, 2068–2078. [Google Scholar]
- Wang, Y.F.; Yu, S.X.; Wang, J. Biomass-dependent susceptibility to drought in experimental grassland communities. Ecol. Lett. 2007, 10, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Deborah, S.; Klaus, V.H.; Kleinebecker, T.; Boeddinghaus, R.S.; Hinderling, J.; Kandeler, E.; Marhan, S.; Nowak, S.; Sonnemann, I.; Wurst, S.; et al. Recovery of Ecosystem functions after experimental disturbance in 73 grasslands differing in land use intensity, plant species richness and community composition. J. Ecol. 2019, 107, 2635–2649. [Google Scholar]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Calderira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in European grasslands. Science 1999, 1286, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, R.J. Meta-analyses and mega-mistakes: Calling time on meta-analysis of the species richness-productivity relationship. Ecology 2010, 91, 2522–2533. [Google Scholar] [CrossRef]
- Li, X.L.; Gao, J.; Zhang, J.; Hai, Q.S. Natural and anthropogenic influences on the spatiotemporal change of degraded meadows in southern Qinghai province, West China: 1976–2015. Appl. Geogr. 2018, 97, 176. [Google Scholar] [CrossRef]
- Zhang, X.J.; Ma, K.X.; Liu, X.B.; Yang, F.L.; Lin, H.L. Spatiotemporal patterns of windbreak and sand fixation before and after grazing prohibition in Ningxia. Pratacult. Sci. 2023, 40, 638–653. [Google Scholar]
- Ma, Y.Y.; Li, Q.; Feng, P.; Yang, Z.Z. Diversity of ground-dwelling spider community in different restoring times of post-fire forest, Cangshan Mountain, Yunnan Province. Acta Ecol. Sin. 2013, 33, 0964–0974. [Google Scholar]
- Zhang, H.Y.; Ou, X.H. Using insect for indicator to monitor and assess forest Ecosystem Health. World For. Res. 2006, 19, 22–25. [Google Scholar]
- Liu, M.Q.; Hu, F.; Li, H.X.; Chen, X.Y.; He, Y.Q. Soil Arthropod Communities Under Different Artificial Woodland Restored on Degraded Red Soil. Acta Ecol. Sin. 2002, 22, 54–61. [Google Scholar]
- Zhang, J.T.; Zhao, F.X.; Chen, J.H.; Li, Y.; Sun, J. Correlations of species diversity and biomass with environmental factors in alpine grasslands across the Qingzang Plateau. Pratacult. Sci. 2023, 40, 616–626. [Google Scholar]
- Yao, X.X.; Zhou, R.; Li, C.H.; Dou, J.S.N. Effects of solpe aspect on vegetation distribution pattern and forage quality characteristics of alpine grassland in the Qinghai-Tibet plateau. Acta Agrestia Sin. 2021, 29, 2792–2799. [Google Scholar]
- Duan, M.J.; Gao, Q.Z.; Guo, Y.Q.; Wan, Y.F.; Li, Y.; Gan, Z.Z.B.; Dan, J.L.B.; Wei, L.T.; Xi, R.Z.M. Species diversity distribution pattern of alpine grassland communities along an altitudinal gradient in the Northern Tibet. Pratacult. Sci. 2011, 28, 1845–1850. [Google Scholar]
- Bai, Y.F.; Li, L.H.; Wang, Q.B.; Zhang, L.X.; Zhang, Y.; Chen, Z.Z. Changes in plant species diversity and productivity along gradients of precipitation and elevation in Xilin River basin, Inner Mongolia. Chin. J. Plant Ecol. 2000, 24, 667–673. [Google Scholar]
- Guo, Y.R.; Zhao, H.L.; Zhao, X.Y.; Zuo, X.A.; Yun, J.Y. Study on the relationship between species diversity and productivity of sandy grassland communities in Horqin sandland. Zrid Zone Res. 2007, 24, 198–203. [Google Scholar]
- Yang, L.M.; Zhou, G.S.; Li, J.D. Relationship between productivity and plant species diversity of grassland communities in Songnen plain of northeast China. Chin. J. Plant Ecol. 2002, 26, 589–593. [Google Scholar]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Huang, J.H.; Lu, X.H.; Guo, Z.J. Service functions of the natural forest ecosystem in Burqin Forest Farm, Xinjiang. Arid Zone Res. 2014, 31, 866–873. [Google Scholar]
- Huang, J.H.; Lu, X.H.; Guo, Z.J.; Wang, J.P.; Zang, R.G. Assessment of natural forest ecosystem services in Buerjin County, Xinjiang. J. Beijing For. Univ. 2015, 37, 62–69. [Google Scholar]
- Yin, X.K.; Ye, M.; Guo, J.X.; Zhang, K.L.; Zhao, F.F. Relationship between species diversity and productivity of different grassland types in Buerjin forest area of Altai mountains. J. Soil Water Conserv. 2022, 36, 110–115. [Google Scholar]
- Xue, R.H.; Jiao, L.; Liu, X.P.; Chen, K. Evaluation of the stability of the radial growth of Larix sibirica at different altitudes in response to climate change in Altai Mountains, Xinjia. Chin. J. Ecol. 2021, 40, 1275–1284. [Google Scholar]
- Xu, Q.; Zhao, W.Y.; Wei, Y.; Ye, M.; Zhao, X.F. Forestry structure and spatial distribution pattern of different age tree species in forest area of eastern Altai Mountain. Arid Zone Res. 2022, 39, 1885–1895. [Google Scholar]
- Zhang, D.L.; Li, Y.B.; Yang, Y.P.; Lan, B. Synthesized Climate Change in the North Altay Mountains in the Past 2000 Years. Arid Zone Res. 2019, 36, 176–185. [Google Scholar]
- Jiang, S.X.; Zhang, T.W.; Yu, S.L.; Shang, H.M.; Zhang, R.B.; Qin, L.; Yuan, Y.J. Integrated Reconstruction of the Historical Climate Series for the Tianshan Mountains and the Altay Mountains. Desert Oasis Meteorol. 2022, 16, 102–111. [Google Scholar]
- Hu, J.; Yu, S.L.; Yuan, Y.J.; Shang, H.M.; Zhang, T.W.; Zhang, R.B. Characteristics of tree-ring width chronologies in the central Altai Mountains and climate response. Desert Oasis Meteorol. 2014, 8, 19–26. [Google Scholar]
- Xu, H.J.; Han, B.P. Main soil types and soil vertical distribution in Altay mountains Area, Xinjiang. Chin. J. Soil Sci. 2008, 39, 465–470. [Google Scholar]
- Ma, K.P. Measuring methods of biodiversity i. Measuring methods of α diversity (part one). Chin. Biodivers. 1994, 2, 229–239. [Google Scholar]
- Ma, K.P. Measuring methods of biodiversity i. Measuring methods of α diversity (part two). Chin. Biodivers. 1994, 2, 231–239. [Google Scholar]
- Zhao, X.J.; Liang, Z.D.; Shao, L.J.; Zhao, X.F. Analysis and Evaluation on Nonlinear Regression Function of SPSS Software. Stat. Decis. 2021, 23, 20–22. [Google Scholar]
- Luo, Y.; He, Y.B.; Li, D.Z.; Wang, Y.H.; Yi, T.S.; Wang, H. A Comparison of Classifications of Families of Chinese Vascular Plants among Flora Republicae Popularis Sinicae, Flora of China and the New Classifications. Plant Divers. Resour. 2012, 34, 231–238. [Google Scholar] [CrossRef]
- Ma, W.L.; Shi, P.L.; Li, W.H.; He, Y.T.; Zhang, X.Z.; Shen, Z.X. The change of individual plant traits and biomass allocation in alpine meadow with elevation variation on the Qinghai-Tibetan Plateau. Sci. China Press 2010, 40, 533–543. [Google Scholar] [CrossRef]
- Luo, L.M.; Miao, Y.J.; Wu, J.S.; Pan, Y.; Tu, Y.L.; Yu, C.Q.; Zhao, Y.; Zhao, G.F.; Wu, J.X. Variation characteristics of species diversity of mountain shrub grassland in Lhasa River Valley with increasing altitude. Acta Pratacult. Sin. 2014, 23, 320–423. [Google Scholar]
- Ma, J.J.; Liu, Y.H.; Sheng, J.D.; Li, N.; Wu, H.Q.; Jia, H.T.; Sun, Z.J.; Cheng, J.H. Changes of relationships between dominant species and their relative biomass along elevational gradients in Xinjiang grasslands. Acta Pratacult. Sin. 2021, 30, 25–35. [Google Scholar]
- SU, J.J.; Liu, Y.P.; Liu, L.Y.; Wu, T.Z.; Wang, M. Quantitative classification and ordination analysis of typical plant communities in Altay Prefecture of Xinjiang. Acta Pratacult. Sin. 2023, 32, 50–67. [Google Scholar]
- He, B.; Wang, G.Y.; Su, H.C.; Shen, Y.P. Response of extreme hydrological events to climate change in the regions of Altay Mountains, Xinjiang. J. Glaciol. Geocryol. 2012, 34, 927–933. [Google Scholar]
- Liu, Y.Y.; Hu, Y.K.; Wang, X.; Gong, Y.M. Vertical differentiation characteristics of species diversity and biomass of alpine grassland in the middle of the southern slope of Tianshan Mountain. Chin. J. Ecol. 2013, 32, 311–318. [Google Scholar]
- Ehud, M. From patterns to function in living systems: Dryland ecosystems as a case study. Annu. Rev. Condens. Matter Phys. 2018, 9, 79–103. [Google Scholar]
- Wang, C.T.; Cao, G.M.; Wang, Q.L.; Jing, Z.C.; Ding, L.M.; Long, R.J. Changes of plant community species composition and biomass along environmental gradient in the alpine grassland of Qinghai-Tibet Plateau. Sci. China Press 2007, 37, 585–592. [Google Scholar]
- Guan, H.L.; Fan, J.W.; Li, Y.Z. The impact of different introduced artificial grassland species combinations on community biomass and species diversity in temperate steppe of the Qinghai-Tibetan Plateau. Acta Pratacult. Sin. 2019, 28, 192–201. [Google Scholar]
- Zhang, Y.J.; Zhu, J.T.; Shen, R.N.; Wang, L. Research progress on the effects of grazing on grassland ecosystem. Chin. J. Plant Ecol. 2020, 44, 553–564. [Google Scholar] [CrossRef]
- Zhou, W.S.; Zhang, C.T.; Zhou, Z.P.; Zhou, T. Correction of the relationship between altitude and latitude of vegetation distribution. Sci. Sin. Vitae 2021, 51, 334–345. (In Chinese) [Google Scholar] [CrossRef]
- Niu, Y.J.; Zhou, J.W.; Yang, S.W.; Wang, G.Z.; Liu, L.; Hua, L.M. Quantitative apportionment of slope aspect and altitude to soil moisture and temperature and plant distribution on alpine meadow. Chin. J. Appl. Ecol. 2017, 28, 1489–1497. [Google Scholar]
- Liu, Y.; Zhang, J.; Yang, W.Q. Responses of alpine biodiversity to climate change. Biodivers. Sci. 2009, 17, 88–96. [Google Scholar]
- Deng, D.Z.; He, L.; Yan, W.X.; Chen, D.C. Analysis of spatial distribution pattern of dominant populations of desertification grassland in northwest Sichuan. Acta Agrestia Sin. 2017, 25, 492–498. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Large Plot | Humans | Constructive Species | Main Companion Species |
|---|---|---|---|
| Wuxilike | Autumn pasture | Androsace umbellata (Lour.) Merr. | Fragaria vesca L. |
| Eleusine indica (L.) Gaertn. | Galium odoratum (L.) Scop. | ||
| Kobresia capillifolia (Decne.) C. B. Clarke | |||
| Dadonggou | Front range | Piper kadsura (Choisy) Ohwi | Eleusine indica (L.) Gaertn. |
| Taraxacum mongolicum Hand.-Mazz. | Polygonum aviculare L. | ||
| Vally | Summer pasture | Festuca ovina L. | Potentilla chinensis Ser. |
| Eleusine indica (L.) Gaertn. | Plantago asiatica L. | ||
| HB | Stock route | Kobresia capillifolia (Decne.) C. B. Clarke | Imperata cylindrica (L.) Beauv. |
| Stipa sareptana Becker | Gagea nakaiana | ||
| Qiaoati | Road repairs on the side | Festuca ovina L. | Sect. Ranunculoides Chen et C. M. Hu |
| Fragaria vesca L. | Geranium wilfordii Maxim. |
| Environmental Factors | Species Diversity Index | Fitting Equations |
|---|---|---|
| Altitude | Margalef | Y = −2.64 × 10−10x3 + 1.48 × 10−6x2 + −0.0025x + 1.73 |
| Simpson | Y = −4.36 × 10−13x4 − 3.17 × 10−9x3 − 8.54 × 10−6x2 + 0.01x − 3.97 | |
| Shannon–Wiener | Y = 1.01 × 10−10x3 + 6.1 × 10−7x2 + 0.002x − 0.34 | |
| Alatalo | Y = −1.04 × 10−12x4 − 7.68 × 10−9x3 + 2.08 × 10−5 × 10−5x2 + 0.02x − 9.65 | |
| Longitude | Margalef | Y = 12.68x4 − 4455.51x3 + 587,328.38x2 − 3.44x + 7.56 |
| Simpson | Y = 0.9x4 − 169.48x3 + 12,032.61x2 − 379,617.37x + 4,490,153.97 | |
| Shannon–Wiener | Y = 16.71x4 − 5866.40x3 + 772,541x2 − 4.52x + 9.92 | |
| Alatalo | Y = 9.72x4 − 3414.63x3 + 450,027.34x2 − 2.64x + 5.79 | |
| Latitude | Margalef | Y = 6.65x4 − 1271.80x3 + 91,262.97.19x2 − 2,910,565.38x + 3.48 |
| Simpson | Y = 6.67x4 − 2339.19x3 + 307,869.22x2 − 1.8x + 3.95 | |
| Shannon–Wiener | Y = 11.30x4 − 2160.71x3 + 154,949.32x2 − 4,938,453.13x + 5.9 | |
| Alatalo | Y = 1.68x4 − 319.94x3 + 22,788.57x2 − 721,358.58x + 8,562,119.2 | |
| Coverage | Margalef | Y = −5.83 × 10−7x4 + 1.12 × 10−4x3 + 0.0073x2 − 0.19x + 2.11 |
| Simpson | Y = −2.41 × 10−7x4 − 5.83 × 10−5x3 − 0.0049x2 + 0.16x − 1.31 | |
| Shannon–Wiener | Y = −2.38 × 10−5x3 − 0.0038x2 + 0.19x − 1.98 | |
| Alatalo | Y = −7.16 × 10−7x3 − 1.46 × 10−5x2 − 0.0082x + 1.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, X.; Ye, M.; He, Q.; Zhang, K. Spatial Distribution Pattern and Influencing Factors of Above-Ground Biomass and Species Diversity of Grassland in the Altay Forest Area. Land 2023, 12, 1370. https://doi.org/10.3390/land12071370
Pan X, Ye M, He Q, Zhang K. Spatial Distribution Pattern and Influencing Factors of Above-Ground Biomass and Species Diversity of Grassland in the Altay Forest Area. Land. 2023; 12(7):1370. https://doi.org/10.3390/land12071370
Chicago/Turabian StylePan, Xiaoting, Mao Ye, Qingzhi He, and Kaili Zhang. 2023. "Spatial Distribution Pattern and Influencing Factors of Above-Ground Biomass and Species Diversity of Grassland in the Altay Forest Area" Land 12, no. 7: 1370. https://doi.org/10.3390/land12071370