

Effects of Habitat Heterogeneity and Topographic Variation on Insect Pest Risks in Alpine Regions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Pest Species Data
2.2. Environmental Data
2.3. Species Distribution Modeling
2.4. Delineating the Risk Areas
2.5. Synthesis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Chang. Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef]
- Pellissier, L.; Rasmann, S. The functional decoupling of processes in alpine ecosystems under climate change. Curr. Opin. Insect Sci. 2018, 29, 126–132. [Google Scholar] [CrossRef]
- Grimm, N.B.; Chapin, F.S., III; Bierwagen, B.; Gonzalez, P.; Groffman, P.M.; Luo, Y.; Melton, F.; Nadelhoffer, K.; Pairis, A.; Raymond, P.A.; et al. The impacts of climate change on ecosystem structure and function. Front. Ecol. Environ. 2013, 11, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Logan, J.A.; Régnière, J.; Powell, J.A. Assessing the impacts of global warming on forest pest dynamics. Front. Ecol. Environ. 2003, 1, 130–137. [Google Scholar] [CrossRef]
- Netherer, S.; Schopf, A. Potential effects of climate change on insect herbivores in European forests—General aspects and the pine processionary moth as specific example. For. Ecol. Manag. 2010, 259, 831–838. [Google Scholar] [CrossRef]
- Comiti, F.; Mao, L.; Penna, D.; Dell’Agnese, A.; Engel, M.; Rathburn, S.; Cavalli, M. Glacier melt runoff controls bedload transport in Alpine catchments. Earth Planet. Sci. Lett. 2019, 520, 77–86. [Google Scholar] [CrossRef]
- Potter, T.S.; Bowman, W.D. Testing invasion filters for the alpine: The roles of temperature, nitrogen deposition and soil. Biol. Invasions 2020, 22, 1889–1901. [Google Scholar] [CrossRef]
- Nicolosi, G.; Mammola, S.; Verbrugge, L.; Isaia, M. Aliens in caves: The global dimension of biological invasions in subterranean ecosystems. Biol. Rev. 2023, 98, 849–867. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, B.; Neumann, P.; Schweiger, O. Global warming promotes biological invasion of a honey bee pest. Glob. Chang. Biol. 2019, 25, 3642–3655. [Google Scholar] [CrossRef] [Green Version]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Soliman, T.; Mourits, M.C.M.; Lansink, A.O.; Van der Werf, W. Economic impact assessment in pest risk analysis. Crop. Prot. 2010, 29, 517–524. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Economic impact of exotic insect pests in Brazilian agriculture. J. Appl. Entomol. 2013, 137, 1–15. [Google Scholar] [CrossRef]
- Lu, Y.; Jenkins, A.; Ferrier, R.C.; Bailey, M.; Gordon, I.J.; Song, S.; Huang, J.; Jia, S.; Zhang, F.; Liu, X.; et al. Addressing China’s grand challenge of achieving food security while ensuring environmental sustainability. Sci. Adv. 2015, 1, e1400039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biber-Freudenberger, L.; Ziemacki, J.; Tonnang, H.E.; Borgemeister, C. Future risks of pest species under changing climatic conditions. PLoS ONE 2016, 11, e0153237. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, O.; Lischke, H.; Wermelinger, B. Climate change alters elevational phenology patterns of the European spruce bark beetle (Ips typographus). Glob. Chang. Biol. 2019, 25, 4048–4063. [Google Scholar] [CrossRef]
- Hlásny, T.; Zimová, S.; Bentz, B. Scientific response to intensifying bark beetle outbreaks in Europe and North America. For. Ecol. Manag. 2021, 499, 119599. [Google Scholar] [CrossRef]
- Wermelinger, B.; Rigling, A.; Schneider Mathis, D.; Kenis, M.; Gossner, M.M. Climate change effects on trophic interactions of bark beetles in inner alpine Scots pine forests. Forests 2021, 12, 136. [Google Scholar] [CrossRef]
- Bellone, D.; Klapwijk, M.J.; Bjorkman, C. Habitat heterogeneity affects predation of European pine sawfly cocoons. Ecol. Evol. 2017, 7, 11011–11020. [Google Scholar] [CrossRef]
- Barbaro, L.; Rusch, A.; Muiruri, E.W.; Gravellier, B.; Thiery, D.; Castagneyrol, B. Avian pest control in vineyards is driven by interactions between bird functional diversity and landscape heterogeneity. J. Appl. Ecol. 2017, 54, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, M.; De Marco, P. Leaf Beetle (Chrysomelidae: Coleoptera) Assemblages in a Mosaic of Natural and Altered Areas in the Brazilian Cerrado. Neotrop. Entomol. 2015, 44, 242–255. [Google Scholar] [CrossRef]
- Haan, N.L.; Zhang, Y.; Landis, D.A. Predicting landscape configuration effects on agricultural pest suppression. Trends Ecol. Evol. 2020, 35, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaumelle, L.; Auriol, A.; Grasset, M.; Pavy, A.; Thiéry, D.; Rusch, A. Benefits of increased cover crop diversity for predators and biological pest control depend on the landscape context. Ecol. Solut. Evid. 2021, 2, e12086. [Google Scholar] [CrossRef]
- Forman, R.T.; Godron, M. Patches and structural components for a landscape ecology. BioScience 1981, 31, 733–740. [Google Scholar]
- Turner, M.G. Landscape ecology: The effect of pattern on process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Cardina, J.; Johnson, G.A.; Sparrow, D.H. The nature and consequence of weed spatial distribution. Weed Sci. 1997, 45, 364–373. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ranson, K.J.; Fedotova, E.V. Spatial pattern of Siberian silkmoth outbreak and taiga mortality. Scand. J. For. Res. 2007, 22, 531–536. [Google Scholar] [CrossRef]
- Probert, A.F.; Ward, D.F.; Beggs, J.R.; Lin, S.L.; Stanley, M.C. Conceptual risk framework: Integrating ecological risk of introduced species with recipient ecosystems. Bioscience 2020, 70, 71–79. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J. Determining key monitoring areas for the 10 most important weed species under a changing climate. Sci. Total Environ. 2019, 683, 568–577. [Google Scholar] [CrossRef]
- Wang, C.J.; Wang, R.; Yu, C.M.; Dang, X.P.; Sun, W.G.; Li, Q.F.; Wang, X.T.; Wan, J.Z. Risk assessment of insect pest expansion in alpine ecosystems under climate change. Pest Manag. Sci. 2021, 77, 3165–3178. [Google Scholar] [CrossRef]
- Wang, P.; Deng, X.Z.; Jiang, S.J. Diffused impact of grassland degradation over space: A case study in Qinghai province. Phys. Chem. Earth 2017, 101, 166–171. [Google Scholar] [CrossRef]
- Fløjgaard, C.; Normand, S.; Skov, F.; Svenning, J.C. Deconstructing the mammal species richness pattern in Europe–towards an understanding of the relative importance of climate, biogeographic history, habitat heterogeneity and humans. Glob. Ecol. Biogeogr. 2011, 20, 218–230. [Google Scholar] [CrossRef]
- Xu, Y.; Shen, Z.; Ying, L.; Zang, R.; Jiang, Y. Effects of current climate, paleo-climate, and habitat heterogeneity in determining biogeographical patterns of evergreen broad-leaved woody plants in China. J. Geogr. Sci. 2019, 29, 1142–1158. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Kolyaie, S.; Morton, D.C.; Zimmerman, J.K.; Uriarte, M. Effects of topography on tropical forest structure depend on climate context. J. Ecol. 2020, 108, 145–159. [Google Scholar] [CrossRef]
- Wang, C.J.; Wang, S.J.; Yu, C.M.; Wang, X.T.; Wang, R.; Wan, J.Z. Habitat heterogeneity and topographic variation as the drivers of insect pest distributions in alpine landscapes. Acta Ecol. Sin. 2023, 43, 596–603. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudı, K.M.; Chee, Y.E.; Yates, C.J. Astatistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Li, R.Q.; Xu, M.; Wong, M.H.G.; Qiu, S.; Li, X.H.; Ehrenfeld, D.; Li, D.M. Climate change threatens giant panda protection in the 21st century. Biol. Conserv. 2015, 182, 93–101. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Zimmermann, N.E.; Pouteau, R.; Yu, F.H. Current and future plant invasions in protected areas: Does clonality matter? Divers. Distrib. 2021, 27, 2465–2478. [Google Scholar] [CrossRef]
- Moilanen, A.; Franco, A.M.A.; Early, R.I.; Fox, R.; Wintle, B.; Thomas, C.D. Prioritizing multiple-use landscapes for conservation: Methods for large multispecies planning problems. Proc. R. Soc. B Biol. Sci. 2005, 272, 1885–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtomaki, J.; Moilanen, A. Methods and workflow for spatial conservation prioritization using Zonation. Environ. Model. Softw. 2013, 47, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on Earth. A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Vehviläinen, H.; Koricheva, J.; Ruohomäki, K. Tree species diversity influences herbivore abundance and damage: Meta-analysis of long-term forest experiments. Oecologia 2007, 152, 287–298. [Google Scholar] [CrossRef]
- Moreira, X.; Abdala-Roberts, L.; Rasmann, S.; Castagneyrol, B.; Mooney, K.A. Plant diversity effects on insect herbivores and their natural enemies: Current thinking, recent findings, and future directions. Curr. Opin. Insect Sci. 2016, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jactel, H.; Moreira, X.; Castagneyrol, B. Tree diversity and forest resistance to insect pests: Patterns, mechanisms, and prospects. Annu. Rev. Entomol. 2021, 66, 277–296. [Google Scholar] [CrossRef]
- Guyot, V.; Castagneyrol, B.; Vialatte, A.; Deconchat, M.; Selvi, F.; Bussotti, F.; Jactel, H. Tree diversity limits the impact of an invasive forest pest. PLoS ONE 2015, 10, e0136469. [Google Scholar] [CrossRef] [Green Version]
- Guyot, V.; Castagneyrol, B.; Vialatte, A.; Deconchat, M.; Jactel, H. Tree diversity reduces pest damage in mature forests across Europe. Biol. Lett. 2016, 12, 20151037. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Fei, S.; Potter, K.M.; Liebhold, A.M.; Wen, J. Tree diversity regulates forest pest invasion. Proc. Natl. Acad. Sci. USA 2019, 116, 7382–7386. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.W.; Ji, G.X.; Liu, S.Y.; Li, B.; Lian, L.; He, W.H.; Zhang, P. Physiological adaptations of Elymus dahuricus to high altitude on the Qinghai-Tibetan Plateau. Acta Physiol. Plant 2019, 41, 115. [Google Scholar] [CrossRef]
- Stenberg, J.A. A conceptual framework for integrated pest management. Trends Plant Sci. 2017, 22, 759–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, D.P.; Tobias, J.A.; Sheil, D.; Meijaard, E.; Laurance, W.F. Maintaining ecosystem function and services in logged tropical forests. Trends Ecol. Evol. 2014, 29, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keesstra, S.; Nunes, J.; Novara, A.; Finger, D.; Avelar, D.; Kalantari, Z.; Cerdà, A. The superior effect of nature based solutions in land management for enhancing ecosystem services. Sci. Total Environ. 2018, 610, 997–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grecchi, R.C.; Gwyn, Q.H.J.; Bénié, G.B.; Formaggio, A.R.; Fahl, F.C. Land use and land cover changes in the Brazilian Cerrado: A multidisciplinary approach to assess the impacts of agricultural expansion. Appl. Geogr. 2014, 55, 300–312. [Google Scholar] [CrossRef]
- Lai, L.; Huang, X.; Yang, H.; Chuai, X.; Zhang, M.; Zhong, T.; Chen, Z.; Chen, Y.; Wang, X.; Thompson, J.R. Carbon emissions from land-use change and management in China between 1990 and 2010. Sci. Adv. 2016, 2, e1601063. [Google Scholar] [CrossRef] [Green Version]




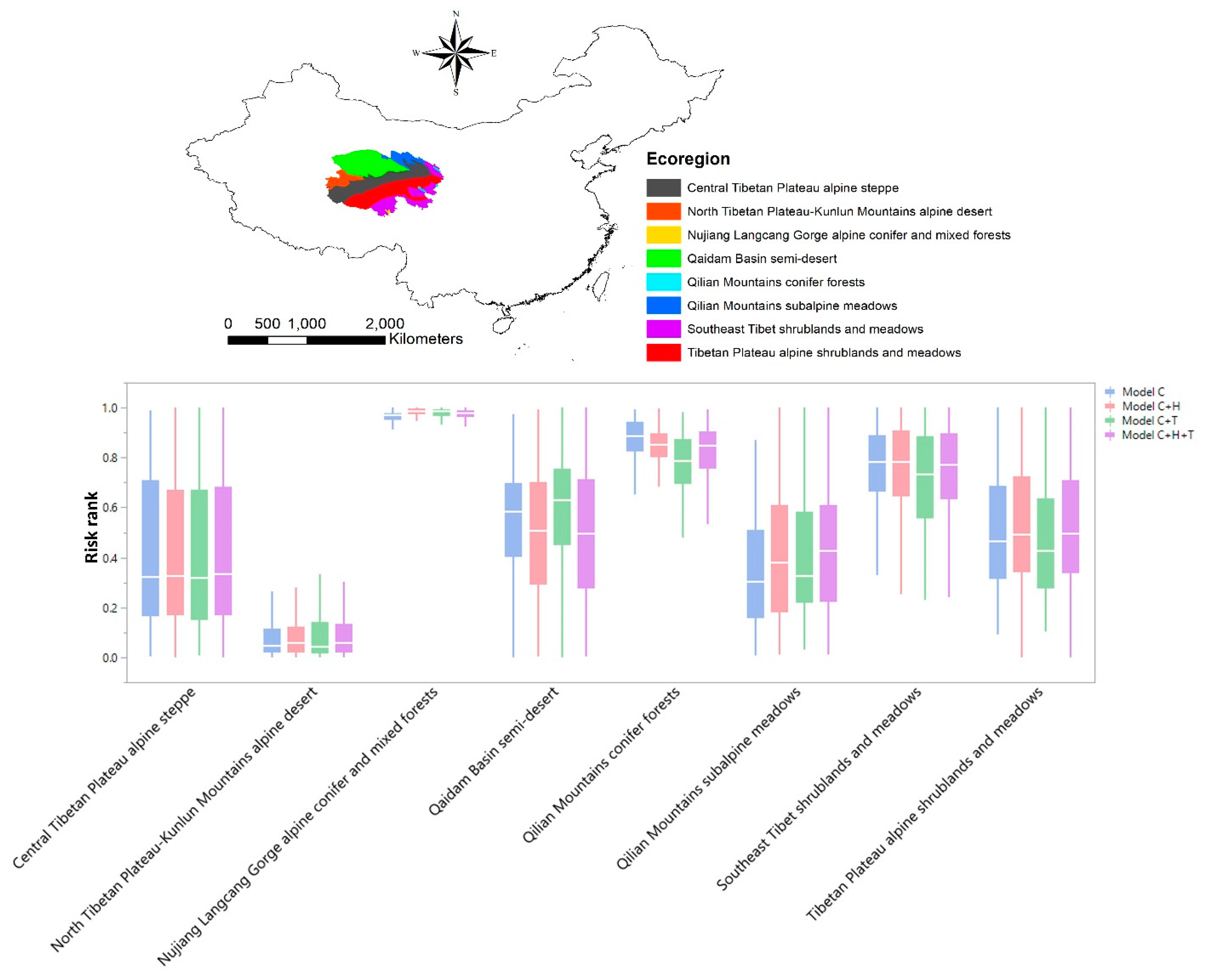{kind=link}
{kind=link}
{kind=link}
| Ecoregion | Code | Clim | Clim + H | Clim + T | Clim + H + T |
|---|---|---|---|---|---|
| Central Tibetan Plateau alpine steppe | CTPAS | 0.424 | 0.413 | 0.415 | 0.419 |
| North Tibetan Plateau–Kunlun Mountains alpine desert | NTPKMAD | 0.070 | 0.087 | 0.110 | 0.101 |
| Nujiang Langcang Gorge alpine conifer and mixed forests | NLGACMF | 0.962 | 0.982 | 0.977 | 0.973 |
| Qaidam Basin semi-desert | QBSD | 0.566 | 0.517 | 0.607 | 0.509 |
| Qilian Mountains conifer forests | QMCF | 0.878 | 0.839 | 0.777 | 0.822 |
| Qilian Mountains subalpine meadows | QMSM | 0.354 | 0.441 | 0.419 | 0.468 |
| Southeast Tibet shrublands and meadows | STSM | 0.751 | 0.767 | 0.716 | 0.758 |
| Tibetan Plateau alpine shrublands and meadows | TPASM | 0.516 | 0.530 | 0.479 | 0.527 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhang, F.-X.; Li, L.-P.; Wang, C.-J.; Wan, J.-Z. Effects of Habitat Heterogeneity and Topographic Variation on Insect Pest Risks in Alpine Regions. Land 2023, 12, 1314. https://doi.org/10.3390/land12071314
Wang L, Zhang F-X, Li L-P, Wang C-J, Wan J-Z. Effects of Habitat Heterogeneity and Topographic Variation on Insect Pest Risks in Alpine Regions. Land. 2023; 12(7):1314. https://doi.org/10.3390/land12071314
Chicago/Turabian StyleWang, Lei, Fei-Xue Zhang, Lan-Ping Li, Chun-Jing Wang, and Ji-Zhong Wan. 2023. "Effects of Habitat Heterogeneity and Topographic Variation on Insect Pest Risks in Alpine Regions" Land 12, no. 7: 1314. https://doi.org/10.3390/land12071314