
The Role of Almond-Leaved Pear Pyrus spinosa Forssk. in Mediterranean Pasturelands Carbon Storage and Woodlands Restoration
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
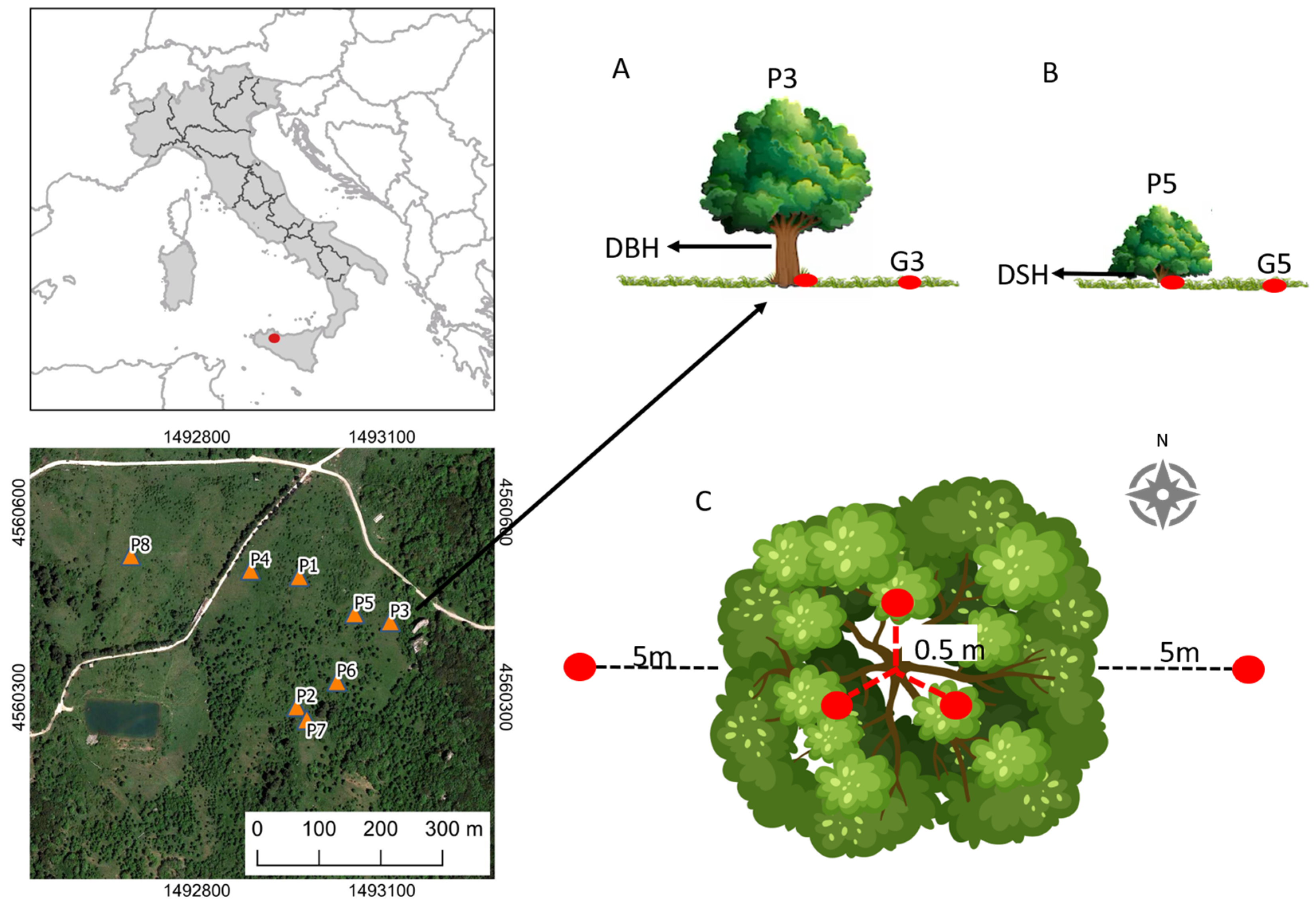
2.1. Study Area
2.2. Experimental Design
2.3. Plant Traits and Aboveground Biomass
2.4. Soil Sampling and Parameters
2.5. Statistical Analysis
3. Results
3.1. Pear vs. Pastureland
3.2. Intra and Inter Pear Individual Variation
3.3. Comparison with Other Studies
4. Discussion
4.1. Pear Effect on Carbon Storage
4.2. Influence of Age and Tree Structure
4.3. Comparison with Literature Data on SOC in Mediterranean Pasturelands
4.4. Management Implications
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Brogniez, D.; Ballabio, C.; Stevens, A.; Jones, R.J.A.; Montanarella, L.; van Wesemael, B. A map of the topsoil organic carbon content of Europe generated by a generalized additive model. Eur. J. Soil Sci. 2015, 66, 121–134. [Google Scholar] [CrossRef]
- Ferreira, C.S.S.; Seifollahi-Aghmiuni, S.; Destouni, G.; Ghajarnia, N.; Kalantari, Z. Soil degradation in the European Mediterranean region: Processes, status and consequences. Sci. Total Environ. 2022, 805, 150106. [Google Scholar] [CrossRef] [PubMed]
- Salvati, L.; Kosmas, C.; Kairis, O.; Karavitis, C.; Acikalin, S.; Belgacem, A.; Solé-Benet, A.; Chaker, M.; Fassouli, V.; Gokceoglu, C.; et al. Unveiling soil degradation and desertification risk in the Mediterranean basin: A data mining analysis of the relationships between biophysical and socioeconomic factors in agro-forest landscapes. J. Environ. Plan. Manag. 2015, 58, 1789–1803. [Google Scholar] [CrossRef]
- Catania, V.; Bueno, R.S.; Alduina, R.; Grilli, E.; La Mantia, T.; Castaldi, S.; Quatrini, P. Soil microbial biomass and bacterial diversity in southern European regions vulnerable to desertification. Ecol. Indic. 2022, 145, 109725. [Google Scholar] [CrossRef]
- Morán-Ordóñez, A.; Ameztegui, A.; De Cáceres, M.; de-Miguel, S.; Lefèvre, F.; Brotons, L.; Coll, L. Future trade-offs and synergies among ecosystem services in Mediterranean forests under global change scenarios. Ecosyst. Serv. 2020, 45, 101174. [Google Scholar] [CrossRef]
- FAO; Plan Bleu. State of Mediterranean Forests 2018; Food and Agriculture Organization of the United Nations: Rome, Italy; Plan Bleu: Marseille, France, 2018. [Google Scholar]
- Badalamenti, E.; Battipaglia, G.; Gristina, L.; Novara, A.; Rühl, J.; Sala, G.; Sapienza, L.; Valentini, R.; La Mantia, T. Carbon stock increases up to old growth forest along a secondary succession in Mediterranean island ecosystems. PLoS ONE 2019, 14, e0220194. [Google Scholar] [CrossRef]
- Bell, S.M.; Terrer, C.; Barriocanal, C.; Jackson, R.B.; Rosell-Melé, A. Soil organic carbon accumulation rates on Mediterranean abandoned agricultural lands. Sci. Total Environ. 2021, 759, 143535. [Google Scholar] [CrossRef]
- Lasanta, T.; Sánchez-Navarrete, P.; Medrano-Moreno, L.M.; Khorchani, M.; Nadal-Romero, E. Soil quality and soil organic carbon storage in abandoned agricultural lands: Effects of revegetation processes in a Mediterranean mid-mountain area. Land Degrad. Dev. 2020, 31, 2830–2845. [Google Scholar] [CrossRef]
- Plieninger, T.; Hartel, T.; Martín-López, B.; Beaufoy, G.; Bergmeier, E.; Kirby, K.; Montero, M.J.; Moreno, G.; Oteros-Rozas, E.; Van Uytvanck, J. Wood-pastures of Europe: Geographic coverage, social–ecological values, conservation management, and policy implications. Biol. Conserv. 2015, 190, 70–79. [Google Scholar] [CrossRef]
- Eurostat. LUCAS—Land Use and Land Cover Survey. 2021. Available online: https://ec.europa.eu/eurostat/databrowser/view/LAN_LCV_OVW__custom_7683636/default/bar?lang=en (accessed on 3 September 2022).
- Sitters, J.; Wubs, E.R.J.; Bakker, E.S.; Crowther, T.W.; Adler, P.B.; Bagchi, S.; Bakker, J.D.; Biederman, L.; Borer, E.T.; Cleland, E.E.; et al. Nutrient availability controls the impact of mammalian herbivores on soil carbon and nitrogen pools in grasslands. Glob. Chang. Biol. 2020, 26, 2060–2071. [Google Scholar] [CrossRef]
- Pellis, G.; Chiti, T.; Rey, A.; Curiel Yuste, J.; Trotta, C.; Papale, D. The ecosystem carbon sink implications of mountain forest expansion into abandoned grazing land: The role of subsoil and climatic factors. Sci. Total Environ. 2019, 672, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Pulido, M.; Schnabel, S.; Lavado Contador, J.F.; Lozano-Parra, J.; Gómez-Gutiérrez, Á.; Brevik, E.C.; Cerdà, A. Reduction of the frequency of herbaceous roots as an effect of soil compaction induced by heavy grazing in rangelands of SW Spain. CATENA 2017, 158, 381–389. [Google Scholar] [CrossRef]
- Shah, A.N.; Tanveer, M.; Shahzad, B.; Yang, G.; Fahad, S.; Ali, S.; Bukhari, M.A.; Tung, S.A.; Hafeez, A.; Souliyanonh, B. Soil compaction effects on soil health and cropproductivity: An overview. Environ. Sci. Pollut. Res. 2017, 24, 10056–10067. [Google Scholar] [CrossRef] [PubMed]
- Rolo, V.; López-Díaz, M.L.; Moreno, G. Shrubs affect soil nutrients availability with contrasting consequences for pasture understory and tree overstory production and nutrient status in Mediterranean grazed open woodlands. Nutr. Cycl. Agroecosystems 2012, 93, 89–102. [Google Scholar] [CrossRef]
- Seddaiu, G.; Bagella, S.; Pulina, A.; Cappai, C.; Salis, L.; Rossetti, I.; Lai, R.; Roggero, P.P. Mediterranean cork oak wooded grasslands: Synergies and trade-offs between plant diversity, pasture production and soil carbon. Agrof. Syst. 2018, 92, 893–908. [Google Scholar] [CrossRef]
- La Mantia, T.; Gristina, L.; Rivaldo, E.; Pasta, S.; Novara, A.; Rühl, J. The effects of post-pasture woody plant colonization on soil and aboveground litter carbon and nitrogen along a bioclimatic transect. Iforest Biogeosci. For. 2013, 6, 238–246. [Google Scholar] [CrossRef]
- European Commission. Nature Restoration Law—For People, Climate, and Planet; Publications Office of the European Union: Brussels, Belgium, 2023. [Google Scholar]
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef]
- Wieczorkowski, J.D.; Lehmann, C.E.R. Encroachment diminishes herbaceous plant diversity in grassy ecosystems worldwide. Glob. Chang. Biol. 2022, 28, 5532–5546. [Google Scholar] [CrossRef]
- Oggioni, S.D.; Ochoa-Hueso, R.; Peco, B. Livestock grazing abandonment reduces soil microbial activity and carbon storage in a Mediterranean Dehesa. Appl. Soil Ecol. 2020, 153, 103588. [Google Scholar] [CrossRef]
- Bueno, R.S.; García, D.; Galetti, M.; La Mantia, T. Past cover modulates the intense and spatially structured natural regeneration of woody vegetation in a pastureland. Plant Ecol. 2020, 221, 205–218. [Google Scholar] [CrossRef]
- Oksuz, D.P.; Aguiar, C.A.S.; Tápia, S.; Llop, E.; Lopes, P.; Serrano, A.R.M.; Leal, A.I.; Branquinho, C.; Correia, O.; Rainho, A.; et al. Increasing biodiversity in wood-pastures by protecting small shrubby patches. For. Ecol. Manag. 2020, 464, 118041. [Google Scholar] [CrossRef]
- Paracchini, M.L.; Petersen, J.E.; Hoogeveen, Y.; Bamps, C.; Burfield, I.; van Swaay, C. High Nature Value Farmland in Europe. An Estimate of the Distribution Patterns on the Basis of Land Cover and Biodiversity Data; EUR 23480 EN. JRC47063; OPOCE: Luxembourg, 2008. [Google Scholar]
- Vidaković, A.; Liber, Z.; Šatović, Z.; Idžojtić, M.; Volenec, I.; Zegnal, I.; Pintar, V.; Radunić, M.; Poljak, I. Phenotypic Diversity of Almond-Leaved Pear (Pyrus spinosa Forssk.) along Eastern Adriatic Coast. Forests 2021, 12, 1630. [Google Scholar] [CrossRef]
- Falcinelli, F.; Moraldi, M. Il pero mandorlino (Pyrus amygdaliformis Villars.). Sherwood For. Ed Alberi Oggi 2001, 68, 39–42. [Google Scholar]
- Bueno, R.S.; García, D.; Galetti, M.; La Mantia, T. Trophic and spatial complementarity on seed dispersal services by birds, wild mammals, and cattle in a Mediterranean woodland pasture. Glob. Ecol. Conserv. 2021, 31, e01880. [Google Scholar] [CrossRef]
- Badalamenti, E.; La Mantia, T.; La Mantia, G.; Cairone, A.; La Mela Veca, D. Living and Dead Aboveground Biomass in Mediterranean Forests: Evidence of Old-Growth Traits in a Quercus pubescens Willd. s.l. Stand. Forests 2017, 8, 187. [Google Scholar] [CrossRef]
- Badalamenti, E.; Pasta, S.; La Mantia, T.; La Mela Veca, D.S. Criteria to identify old-growth forests in the Mediterranean: A case study from Sicily based on literature review and some management proposals. Feddes Repert. 2018, 129, 25–37. [Google Scholar] [CrossRef]
- Gianguzzi, L.; La Mantia, A. Le serie di vegetazione della riserva “Bosco Ficuzza, Rocca Busambra, Bosco del Cappeliere e Gorgo del Drago” (Provincia di Palermo). Nat. Sic. 2004, 28, 265–326. [Google Scholar]
- Bazan, G.; Marino, P.; Guarino, R.; Domina, G.; Schicchi, R. Bioclimatology and Vegetation Series in Sicily: A Geostatistical Approach. Ann. Bot. Fenn. 2015, 52, 1–18. [Google Scholar] [CrossRef]
- Fierotti, G.; Dazzi, C.; Raimondi, S. Carta dei suoli della Sicilia (scala 1: 250.000). Regione Siciliana, Assessorato Territorio e Ambiente. Università degli studi di Palermo, Facoltà di Agraria. Istituto di Agronomia Generale, Cattedra di Pedologia. 1988. [Google Scholar]
- Bianchetto, E.; Buscemi, I.; Corona, P.; Giardina, G.; La Mantia, T.; Pasta, S. Fitting the Stocking Rate with Pastoral Resources to Manage and Preserve Mediterranean Forestlands: A Case Study. Sustainability 2015, 7, 7232–7244. [Google Scholar] [CrossRef]
- Gschwantner, T.; Schadauer, K.; Vidal, C.; Lanz, A.; Tomppo, E.; di Cosmo, L.; Robert, N.; Englert Duursma, D.; Lawrence, M. Common Tree Definitions for National Forest Inventories in Europe. Silva Fenn. 2009, 43, 303–321. [Google Scholar] [CrossRef]
- Boyce, R.L.; Ocasio, M. Pyrus calleryana allometric equations and stand structure in southwestern Ohio and northern Kentucky. Invasive Plant Sci. Manag. 2020, 13, 239–246. [Google Scholar] [CrossRef]
- Tabacchi, G.; Di Cosmo, L.; Gasparini, P.; Morelli, S. Stima del Volume e Della Fitomassa Delle Principali Specie Forestali Italiane. Equazioni di Previsione, Tavole del Volume e Tavole Della Fitomassa Arborea Epigea; Consiglio per la Ricerca e la sperimentazione in Agricoltura, Unità di Ricerca per il Monitoraggio e la Pianificazione Forestale: Trento, Italy, 2011. [Google Scholar]
- Meier, E. Identifying and Using Hundreds of Woods Worldwide. Wood Database. 2015. Available online: https://www.wood-database.com/wp-content/uploads/wood-book-sample.pdf (accessed on 7 December 2021).
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Silva, J.S.; Rego, F.C.; Martins-Loução, M.A. Belowground traits of mediterranean woody plants in a Portuguese shrubland. Ecol. Mediterr. 2002, 28, 5–13. [Google Scholar] [CrossRef]
- Marziliano, P.A.; Lafortezza, R.; Medicamento, U.; Lorusso, L.; Giannico, V.; Colangelo, G.; Sanesi, G. Estimating belowground biomass and root/shoot ratio of Phillyrea latifolia L. in the Mediterranean forest landscapes. Ann. For. Sci. 2015, 72, 585–593. [Google Scholar] [CrossRef]
- Olff, H.; Van Andel, J.; Bakker, J.P. Biomass and Shoot/Root Allocation of Five Species from a Grassland Succession Series at Different Combinations of Light and Nutrient Supply. Funct. Ecol. 1990, 4, 193–200. [Google Scholar] [CrossRef]
- De Long, J.R.; Jackson, B.G.; Wilkinson, A.; Pritchard, W.J.; Oakley, S.; Mason, K.E.; Stephan, J.G.; Ostle, N.J.; Johnson, D.; Baggs, E.M.; et al. Relationships between plant traits, soil properties and carbon fluxes differ between monocultures and mixed communities in temperate grassland. J. Ecol. 2019, 107, 1704–1719. [Google Scholar] [CrossRef] [PubMed]
- Passalacqua, N.G.; Aiello, S.; Bernardo, L.; Gargano, D. Monitoring biomass in two heterogeneous mountain pasture communities by image based 3D point cloud derived predictors. Ecol. Indic. 2021, 121, 107126. [Google Scholar] [CrossRef]
- Ma, S.; He, F.; Tian, D.; Zou, D.; Yan, Z.; Yang, Y.; Zhou, T.; Huang, K.; Shen, H.; Fang, J. Variations and determinants of carbon content in plants: A global synthesis. Biogeosciences 2018, 15, 693–702. [Google Scholar] [CrossRef]
- Baruah, T.C.; Barthakur, H.P. A Textbook of Soil Analysis; Vikas Publishing House PVT LTD: New Delhi, India, 1997. [Google Scholar]
- Blake, G.R.; Hartge, K.H. Bulk Density. In Methods of Soil Analysis; Klute, A., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1986; pp. 363–375. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.1.2; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Jendoubi, D.; Liniger, H.; Ifejika Speranza, C. Impacts of land use and topography on soil organic carbon in a Mediterranean landscape (north-western Tunisia). SOIL 2019, 5, 239–251. [Google Scholar] [CrossRef]
- Papini, R.; Valboa, G.; Favilli, F.; L’Abate, G. Influence of land use on organic carbon pool and chemical properties of Vertic Cambisols in central and southern Italy. Agric. Ecosyst. Environ. 2011, 140, 68–79. [Google Scholar] [CrossRef]
- Romanyà, J.; Rovira, P. An appraisal of soil organic C content in Mediterranean agricultural soils. Soil Use Manag. 2011, 27, 321–332. [Google Scholar] [CrossRef]
- Salis, M.; Sepe, L.; Francaviglia, R.; Fedrizzi, M.; Bazzoffi, P.; Claps, S.; Pitzalis, M.; Lilliu, M.; Ruda, P.; Sabia, E.; et al. Environmental effectiveness of GAEC cross-compliance Standard 4.1 (b, c) ‘Protection of permanent pasture land’ and economic evaluation of the competitiveness gap for farmers. Ital. J. Agron. 2015, 10, 714. [Google Scholar] [CrossRef]
- Seddaiu, G.; Porcu, G.; Ledda, L.; Roggero, P.P.; Agnelli, A.; Corti, G. Soil organic matter content and composition as influenced by soil management in a semi-arid Mediterranean agro-silvo-pastoral system. Agric. Ecosyst. Environ. 2013, 167, 1–11. [Google Scholar] [CrossRef]
- Pellis, G. Data for: Ecosystem carbon sink capacity of mountainous forests expansion over abandoned grazing land: The role of subsoil and climatic factors v1. Mendeley Data 2019. [Google Scholar] [CrossRef]
- Chiti, T.; Emanuele, B.; Guido, P.; Lucia, P.; Vincenza, C.M.; Riccardo, V. Soil organic carbon pool’s contribution to climate change mitigation on marginal land of a Mediterranean montane area in Italy. J. Environ. Manag. 2018, 218, 593–601. [Google Scholar] [CrossRef]
- Moreno, G.; Obrador, J.J.; García, A. Impact of evergreen oaks on soil fertility and crop production in intercropped dehesas. Agric. Ecosyst. Environ. 2007, 119, 270–280. [Google Scholar] [CrossRef]
- Rossetti, I.; Bagella, S.; Cappai, C.; Caria, M.C.; Lai, R.; Roggero, P.P.; Martins da Silva, P.; Sousa, J.P.; Querner, P.; Seddaiu, G. Isolated cork oak trees affect soil properties and biodiversity in a Mediterranean wooded grassland. Agric. Ecosyst. Environ. 2015, 202, 203–216. [Google Scholar] [CrossRef]
- Serrano, J.M.; Shahidian, S.; da Silva, J.M.; Carvalho, M. Monitoring of soil organic carbon over 10 years in a Mediterranean silvo-pastoral system: Potential evaluation for differential management. Precis. Agric. 2016, 17, 274–295. [Google Scholar] [CrossRef]
- Sharrow, S.H. Soil compaction by grazing livestock in silvopastures as evidenced by changes in soil physical properties. Agrof. Syst. 2007, 71, 215–223. [Google Scholar] [CrossRef]
- Golodets, C.; Kigel, J.; Sternberg, M. Recovery of plant species composition and ecosystem function after cessation of grazing in a Mediterranean grassland. Plant Soil 2010, 329, 365–378. [Google Scholar] [CrossRef]
- Navarro Cerrillo, R.; Oyonarte, P. Estimation of above-ground biomass in shrubland ecosystems of southern Spain. Investig. Agraria. Sist. Y Recur. For. 2006, 15, 197–207. [Google Scholar] [CrossRef]
- Corona, P.; Pasta, S.; Giardina, G.; La Mantia, T. Assessing the biomass of shrubs typical of Mediterranean pre-forest communities. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2012, 146, 252–257. [Google Scholar] [CrossRef]
- Zribi, L.; Mouillot, F.; Guibal, F.; Rejeb, S.; Rejeb, M.N.; Gharbi, F. Deep Soil Conditions Make Mediterranean Cork Oak Stem Growth Vulnerable to Autumnal Rainfall Decline in Tunisia. Forests 2016, 7, 245. [Google Scholar] [CrossRef]
- Maetzke, F.; Cullotta, S.; La Mantia, T.; La Mela Veca, D.; Pizzurro, G. Individuazione di aree ecologicamente omogenee e di un sistema di aree a priorità di intervento per l’ampliamento della superficie forestale in Sicilia. For. Riv. Selvic. Ecol. For. 2008, 5, 280–295. [Google Scholar] [CrossRef]
- Damigella, P.P.; Alberghina, O. Osservazioni preliminari su alcuni “perastri” (Pyrus sp.) della Sicilia centro-orientale. Tecinica Agric. 1969, Anno XXI, 486–503. [Google Scholar]
- La Mantia, T.; Bueno, R.S. Colonization of eurasian jay Garrulus glandarius and holm oaks Quercus ilex: The establishment of ecological interactions in urban areas. Avocetta 2016, 40, 85–87. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Pastureland | Pear | t | p | |
|---|---|---|---|---|
| SOC (%) | 3.16 ± 0.68 | 3.86 ± 1.21 | 2.18 | 0.035 |
| SOC stock (Mg ha−1) | 113.9 ± 30.3 | 130.3 ± 47.1 | 1.33 | 0.188 |
| Bulk density (g cm−3) | 1.24 ± 0.21 | 1.09 ± 0.17 | −2.49 | 0.018 |
| Land Use | ID | Age | Tree Structure | DBH (cm) | Canopy Cover (m2) | Height (m) | Bulk Density (g cm−3) | SOC (%) | SOC Stock (Mg ha−1) | Biomass C Stock (Mg ha−1) | Total C Stock (Mg ha−1) | C Biomass/C Total (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pear | P1 | old | multi | 27.3 | 30.66 | 4.8 | 0.87 (±0.03) | 3.50 (±0.43) | 93.41 (±11.37) | 3.42 | 96.83 | 3.53 |
| P2 | old | multi | 32.8 | 32.96 | 5.1 | 1.13 (±0.03) | 6.49 (±0.49) | 224.87 (±17.08) | 4.49 | 229.36 | 1.96 | |
| P3 | old | single | 41.5 | 36.83 | 6.2 | 1.15 (±0.03) | 2.81 (±0.65) | 98.69 (±23.07) | 7.53 | 106.21 | 7.09 | |
| P4 | old | single | 27.4 | 21.38 | 4.6 | 1.28 (±0.06) | 3.70 (±0.19) | 142.88 (±7.27) | 2.29 | 145.16 | 1.58 | |
| P5 | young | multi | 15.3 | 13.20 | 4.1 | 0.94 (±0.04) | 3.11 (±0.9) | 92.83 (±26.78) | 3.99 | 96.82 | 4.12 | |
| P6 | young | multi | 11.6 | 12.78 | 3.2 | 0.86 (±0.06) | 3.67 (±0.36) | 96.45 (±9.59) | 1.12 | 97.58 | 1.15 | |
| P7 | young | single | 16.5 | 8.55 | 3.4 | 1.28 (±0.1) | 3.00 (±0.31) | 122.85 (±12.66) | 0.79 | 123.64 | 0.64 | |
| P8 | young | single | 13.2 | 12.22 | 3.7 | 1.23 (±0.07) | 4.57 (±0.15) | 170.61 (±5.45) | 0.60 | 171.21 | 0.35 | |
| Pastureland | G1 | 1.25 (±0.06) | 3.24 (±0.08) | 117.67 (±3.01) | 1.75 | 119.41 | 1.46 | |||||
| G2 | 1.54 (±0.04) | 2.74 (±0.81) | 128.24 (±37.83) | 1.75 | 129.99 | 1.34 | ||||||
| G3 | 0.92 (±0.02) | 4.02 (±1.18) | 124.02 (±7.88) | 1.75 | 125.77 | 1.39 | ||||||
| G4 | 1.11 (±0.01) | 2.95 (±0.29) | 78.64 (±7.83) | 1.75 | 80.39 | 2.17 | ||||||
| G5 | 1.1 (±0.03) | 2.66 (±0.94) | 70.89 (±25.06) | 1.75 | 72.64 | 2.41 | ||||||
| G6 | 1.29 (±0.03) | 2.68 (±0.73) | 107.08 (±28.96) | 1.75 | 108.82 | 1.61 | ||||||
| G7 | 1.3 (±0.01) | 3.39 (±0.23) | 131.59 (±9.04) | 1.75 | 133.33 | 1.31 | ||||||
| G8 | 1.44 (±0.03) | 3.60 (±0.29) | 153.20 (±6.39) | 1.75 | 154.95 | 1.13 |
| SOC | SOC Stock | Bulk Density | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SE | t | p | Estimate | SE | t | p | Estimate | SE | t | p | |
| Intercept | 1.31 | 0.06 | 21.67 | <0.0001 | 130.32 | 10.44 | 12.48 | <0.0001 | 1.09 | 0.02 | 52.47 | <0.0001 |
| Diameter | −0.43 | 0.29 | −1.46 | 0.162 | −28.78 | 50.53 | −0.57 | 0.576 | 0.12 | 0.10 | 1.20 | 0.247 |
| Height | 0.05 | 0.18 | 0.30 | 0.770 | 0.29 | 30.96 | 0.01 | 0.993 | −0.05 | 0.06 | −0.79 | 0.437 |
| Cover | 0.13 | 0.15 | 0.89 | 0.384 | 11.09 | 25.54 | 0.43 | 0.669 | −0.04 | 0.05 | −0.86 | 0.402 |
| Age (young) | −0.30 | 0.17 | −1.78 | 0.092 | −27.32 | 29.43 | −0.93 | 0.366 | 0.03 | 0.06 | 0.49 | 0.628 |
| Tree structure (single) | 0.04 | 0.10 | 0.38 | 0.709 | 11.22 | 16.57 | 0.68 | 0.507 | 0.11 | 0.03 | 3.24 | 0.004 |
| SOC (%) | SOC Stock (Mg ha−1) | Bioclimate | Soil Type | Soil Depth (cm) | Bulk Density (g cm−3) | Region (Country) | Reference |
|---|---|---|---|---|---|---|---|
| 0.74 | - | Thermo | Vertisols | 0–20 | - | Wadi Beja (Tunisia) | [49] |
| 1.22 | 70.0 | Thermo | Vertic Cambisols | 0–40 | 1.43 | Calabria (Italy) | [50] |
| 1.29 | 35.7 | Thermo | Endoleptic Regosols | 0–30 | 1.43 | Sicily (Italy) | [18] |
| 1.12 | - | Meso | Different types | 0–25 | - | Spain | [51] |
| 1.13 | 50.6 | Meso | Dystric Cambisols | 0–30 | 1.47 | La Rioja (Spain) | [9] |
| 1.99 | - | Meso | - | 0–30 | - | Sardinia (Italy) | [52] |
| 2.03 | 82.0 | Meso | Typic Dystroxerept | 0–35 | - | Sardinia (Italy) | [53] |
| 2.11 | - | Meso | - | 0–30 | - | Calabria (Italy) | [52] |
| 2.24 | 54.2 | Meso | Endoleptic Regosols | 0–30 | 1.30 | Sicily (Italy) | [18] |
| 3.86 | 130.3 | Meso | Vertic Haploxeralfs | 0–30 | 1.09 | Sicily (Italy) | Our study (Pyrus) |
| 3.16 | 113.9 | Meso | Vertic Haploxeralfs | 0–30 | 1.24 | Sicily (Italy) | Our study (Grassland) |
| 3.11 | 73.3 | Supra | Haplic Luvisols | 0–30 | 1.27 | Sicily (Italy) | [18] |
| 2.71 | 85.0 | Supra | Phaeozems | 0–30 | 1.32 | Apenin (Italy) | [54] |
| 3.89 | 91.0 | Supra | Cambisols | 0–30 | 1.34 | Apenin (Italy) | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno, R.S.; Badalamenti, E.; Gristina, L.; Novara, A.; La Mantia, T. The Role of Almond-Leaved Pear Pyrus spinosa Forssk. in Mediterranean Pasturelands Carbon Storage and Woodlands Restoration. Land 2023, 12, 2135. https://doi.org/10.3390/land12122135
Bueno RS, Badalamenti E, Gristina L, Novara A, La Mantia T. The Role of Almond-Leaved Pear Pyrus spinosa Forssk. in Mediterranean Pasturelands Carbon Storage and Woodlands Restoration. Land. 2023; 12(12):2135. https://doi.org/10.3390/land12122135
Chicago/Turabian StyleBueno, Rafael Silveira, Emilio Badalamenti, Luciano Gristina, Agata Novara, and Tommaso La Mantia. 2023. "The Role of Almond-Leaved Pear Pyrus spinosa Forssk. in Mediterranean Pasturelands Carbon Storage and Woodlands Restoration" Land 12, no. 12: 2135. https://doi.org/10.3390/land12122135