
Alnus glutinosa Riparian Woodlands of Italy and Corsica: Phytosociological Classification and Floristic Diversity
 , , , , , ,
, , , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
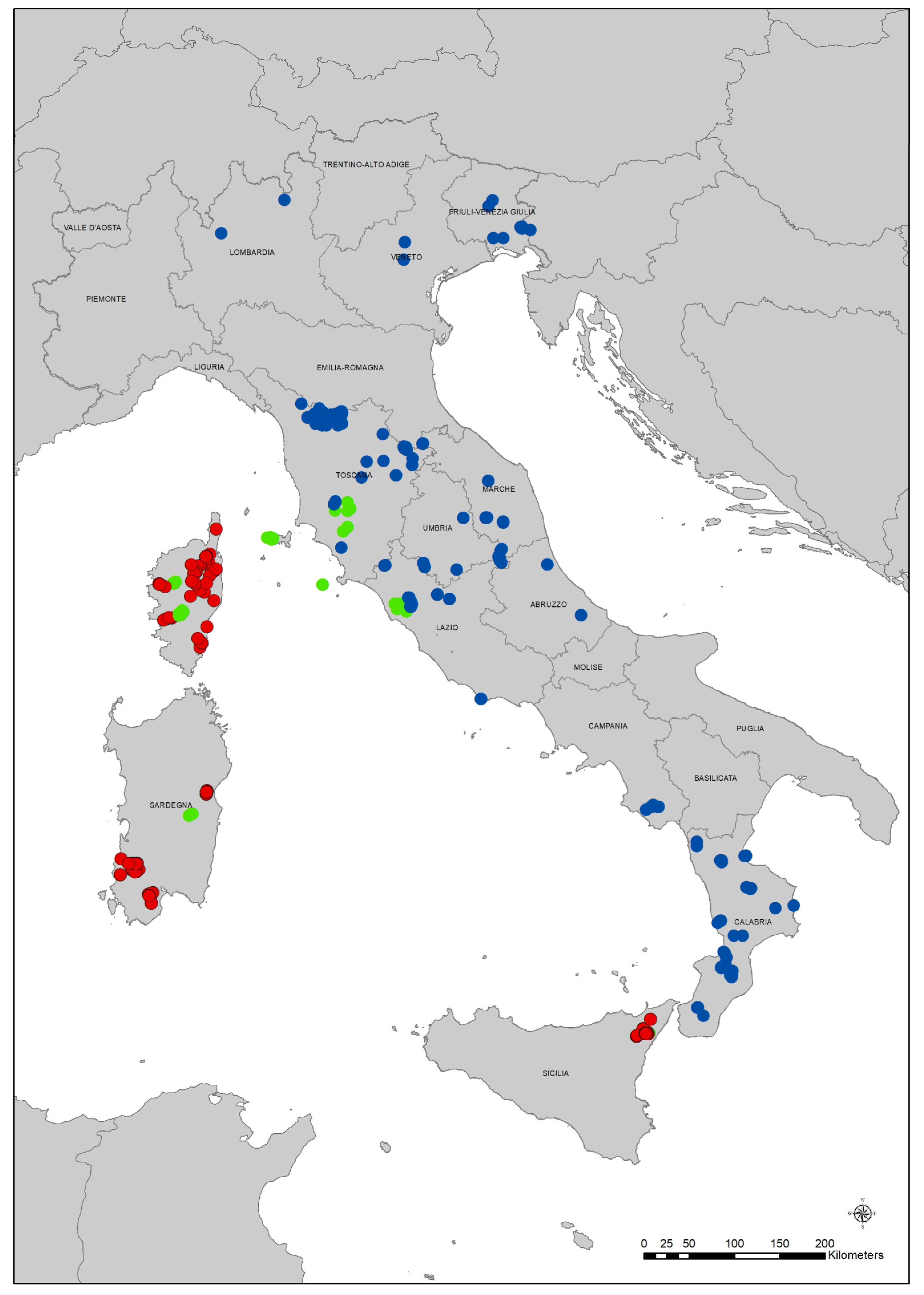
2.1. Phytosociological Data
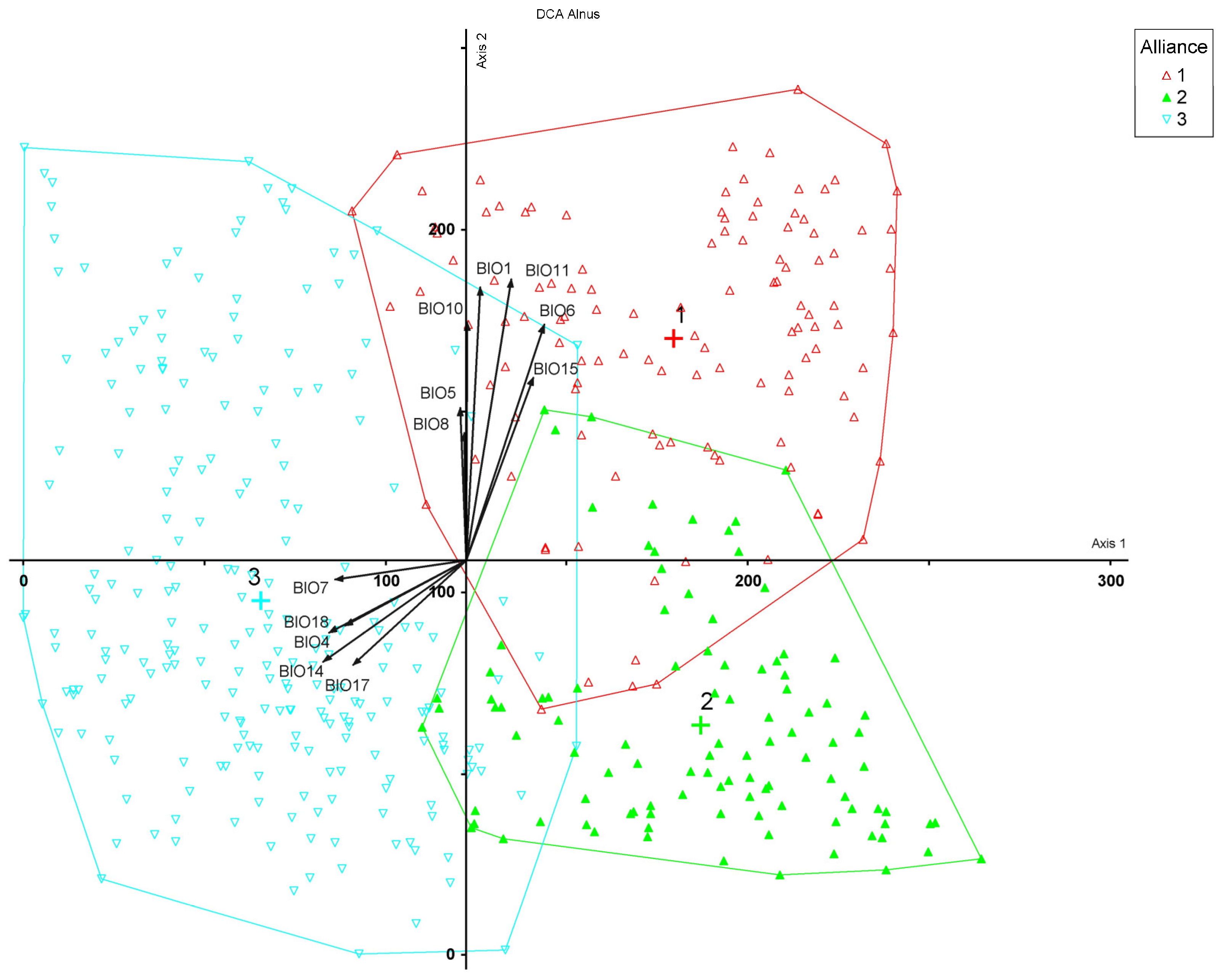
2.2. Statistical Analysis
2.3. Plant Communities Processed
2.4. Taxonomic, Syntaxonomic Nomenclature and Bioclimatic Classification
3. Results and Discussions
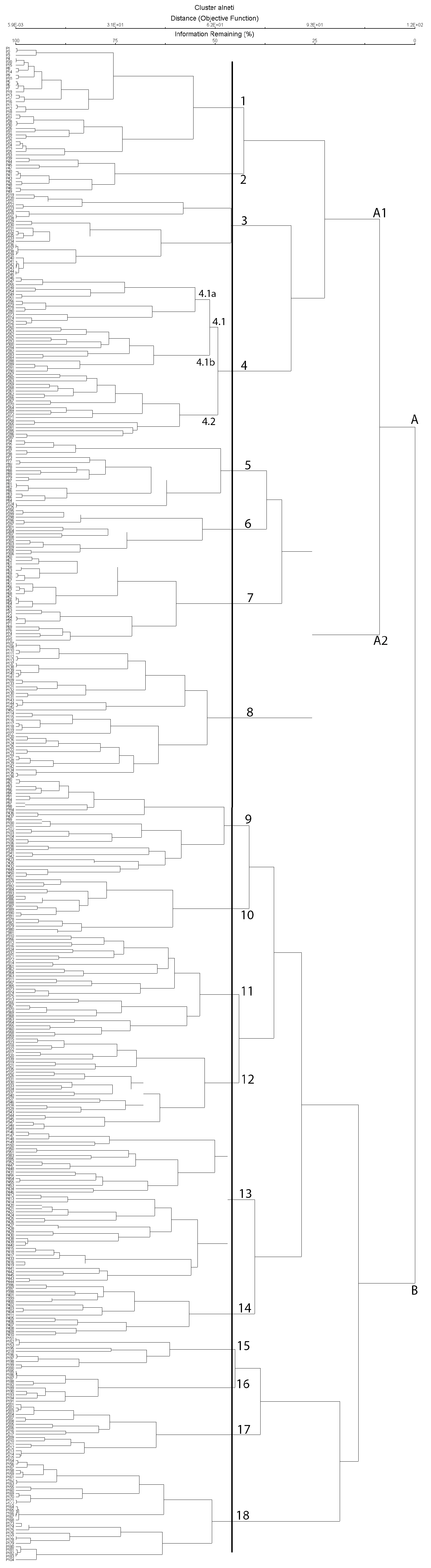
3.1. Syntaxonomical Scheme
3.2. Description of the Vegetation
- 1.Osmundo-Alnetum glutinosae Van den Berghen 1971 (Cluster 1, Figure 5)

- 2. Oenantho crocatae-Alnetum glutinosae Arrigoni et al. 1996 (Cluster 2)
- 3. Platano orientalis-Alnetum glutinosae (Brullo et Spampinato 1990) Sciandrello et al. nom. nov. hoc loco (Cluster 3, Figure 6)

- 4.1. Eupatorio corsici-Alnetum glutinosae (Litard. 1928) Dierschke 1975 (Cluster 4.1a)
- 4.1. Eupatorio corsici-Alnetum glutinosae (Litard. 1928) Dierschke 1975 scolopendrietosum nom. nov. hoc loco (Cluster 4.1b)
- A2. Struthioptero-Alnenion glutinosaesuball. nov. hoc loco (Cluster A2)
- 5. Glechomo sardoae-Alnetum glutinosae Arrigoni 1986 (Cluster 5, Figure 5)
- 6. Gentiano asclepiadeae-Alnetum glutinosae (Litard. and Malcuit 1926) Gamisans 1977 corr. Renaux et al. (2019) (Cluster 6)
- 7. Carici pallescentis-Alnetum glutinosae Landi and Angiolini 2010 (Cluster 7, Figure 7)

- 8. Carpino betuli-Alnetum glutinosae (Di Pietro et al. 2010) Sciandrello et al. nom. nov. hoc loco (Cluster 8, Figure 7)
- B. Ligustro vulgaris–Alnion glutinosae Poldini, Sburlino and Venanzoni 2015 in Biondi et al. 2015 (Cluster B)
- 9. Aro italici-Alnetum glutinosae Gafta and Pedrotti 1996 (Cluster 9)
- 10. Aegopodio podagrariae-Alnetum glutinosae (Blasi and Frondoni 1996) Sciandrello et al. nom. nov. hoc loco (Cluster 10)
- 11. Geranio nodosi-Alnetum glutinosaeass. nov. hoc loco subass. typicum (Cluster 11)
- 12. Geranio nodosi-Alnetum glutinosae cornetosum sanguineaesubass. nov. (Cluster 12)
- 13. Hyperico hircini-Alnetum glutinosae Gafta and Pedrotti 1996 (Cluster 13)
- 14. Lamio orvalae-Alnetum glutinosae Dakskobler 2016 (Cluster 14)
- 15. Polysticho setiferi-Alnetum glutinosae Brullo, Scelsi, Spampinato 2001 (Cluster 15, Figure 7)
- 16. Euphorbio corallioides-Alnetum glutinosae Brullo and Furnari in Barbagallo et al. 1982 (Cluster 16, Figure 7)
- 17. Asperulo taurinae-Alnetum glutinosaeass. nov. hoc loco (Cluster 17)
- 18. Angelico sylvestris-Alnetum glutinosae Brullo and Spampinato 1997 (Cluster 18, Figure 7)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Localities of Relevés
References
- Naiman, R.J.; Decamps, H.; Pollock, M. The Role of Riparian Corridors in Maintaining Regional Biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Lowrance, R. The potential role of riparian forests as buffer zones. In Buffer Zones: Their Processes and Potential in Water Protection; Haycock, N., Burt, T., Goulding, K., Pinay, G., Eds.; Quest Environmental: Harpenden, UK, 1996; pp. 128–133. [Google Scholar]
- Riis, T.; Kelly-Quinn, M.; Aguiar, F.C.; Manolaki, P.; Bruno, D.; Bejarano, M.D.; Clerici, N.; Fernandes, M.R.; Franco, J.C.; Pettit, N.; et al. Global Overview of Ecosystem Services Provided by Riparian Vegetation. BioScience 2020, 70, 501–514. [Google Scholar] [CrossRef]
- European Commission. Interpretation manual of European Union Habitats—EUR28; European Commission DG Environment: Brussels, Belgium, 2013; pp. 1–144. [Google Scholar]
- Cooper, S.D.; Lake, P.S.; Sabater, S.; Melack, J.M.; Sabo, J.L. The effects of land use changes on streams and rivers in mediterranean climates. Hydrobiologia 2013, 719, 383–425. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Bacchetta, G. Knowledge gaps and challenges for conservation of Mediterranean wetlands: Evidence from a comprehensive inventory and literature analysis for Sardinia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 2621–2631. [Google Scholar] [CrossRef]
- Ekka, A.; Pande, S.; Jiang, Y.; der Zaag, P. Anthropogenic Modifications and River Ecosystem Services: A Landscape Perspective. Water 2020, 12, 2706. [Google Scholar] [CrossRef]
- González, E.; González-Sanchis, M.; Cabezas, A.; Comìn, F.A.; Muller, E. Recent changes in the riparian forest of a large regulated mediterranean river: Implications for management. Environ. Manag. 2010, 45, 669–681. [Google Scholar] [CrossRef]
- Dierschke, H. Zur Syntaxonomischen Stellung und Gliederung der Ufer und Auenwälder Südeuropas. Colloq. Phytosociol. 1984, 9, 115–129. [Google Scholar]
- Wallnöfer, S.; Mucina, L.; Grass, V. Querco-Fagetea. In Die Pflanzengesellschaften Österreichs. Teil III. Wälder und Gebüsche; Mucina, L., Grabherr, G., Wallnofer, S., Eds.; Gustav Fischer: Jena, Germany, 1993; pp. 85–236. [Google Scholar]
- Douda, J. Svaz LBA Alnion incanae Pawlowski et al. 1928. In Vegetace Ceske republiky 4. Lesnı a krovinna vegetace [Vegetation of the Czech Republic 4. Forest and Shrub vegetation]; Chytry, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 198–219. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Passarge, H. Neue Vorschlage zur Systematik nordmitteleurop aischer Waldgesellschaften. Feddes Repert. 1968, 77, 75–103. [Google Scholar] [CrossRef]
- Biurrun, I.; Campos, J.A.; García-Mijangos, I.; Herrera, M.; Loidi, J. Floodplain forests of the Iberian Peninsula: Vegetation classification and climatic features. Appl. Veg. Sci. 2016, 19, 336–354. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef]
- Biondi, E.; Allegrezza, M.; Casavecchia, S.; Galdenzi, D.; Gasparri, R.; Pesaresi, S.; Poldini, L.; Sburlino, G.; Vagge, I.; Venanzoni, R. New syntaxonomic contribution to the Vegetation Prodrome of Italy. Plant Biosyst. 2015, 149, 603–615. [Google Scholar] [CrossRef]
- Douda, J.; Boublík, K.; Slezák, M.; Biurrun, I.; Nociar, J.; Havrdová, A.; Doudová, J.; Aćić, S.; Brisse, H.; Brunet, J.; et al. Vegetation classification and biogeography of European floodplain forests and alder carrs. Appl. Veg. Sci. 2016, 19, 147–163. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Fuente, V.; Sánchez-Mata, D. Alisedas mediterráneo-iberoAtlanticas en la Península Iberica. Stud. Bot. 1986, 5, 9–38. [Google Scholar]
- Dierschke, H. Die Schwarzerlen (Alnus glutinosa) Uferwälder Korsikas. Phytocoenologia 1975, 2, 229–243. [Google Scholar] [CrossRef]
- Angius, R.; Bacchetta, G. Boschi e boscaglie ripariali del Sulcis-Iglesiente (Sardegna sud-occidentale, Italia). Braun-Blanquetia 2009, 45, 1–63. [Google Scholar]
- Douda, J.; Doudová, J.; Drašnarová, A.; Kuneš, P.; Hadincová, V.; Krak, K.; Zákravský, P.; Mandák, B. Migration patterns of subgenus Alnus in Europe since the Last Glacial Maximum. A systematic review. PLoS ONE 2014, 9, e88709. [Google Scholar] [CrossRef]
- Mandák, B.; Vít, P.; Krak, K.; Trávníček, P.; Havrdová, A.; Hadincová, V.; Zákravský, P.; Jarolímová, V.; Bacles, C.F.E.; Douda, J. Flow cytometry, microsatellites and niche models reveal the origins and geographical structure of Alnus glutinosa populations in Europe. Ann. Bot. 2016, 117, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria, 1964; pp. 1–330. [Google Scholar]
- Westhoff, V.; van der Mareel, E. The Braun Blanquet approach. In Classification of Plant Communities, 2nd ed.; Whittaker, R.H., Ed.; Dr Junk: The Hague, The Netherlands, 1978; pp. 287–399. [Google Scholar]
- Dengler, J.; Berg, C.; Jansen, F. New ideas for modern phytosociological monographs. Ann. Bot. (Roma) 2005, 5, 193–210. [Google Scholar]
- Dengler, J.; Chytrý, M.; Ewald, J. Phytosociology. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; pp. 2767–2779. [Google Scholar]
- Biondi, E. Phytosociology today: Methodological and conceptual evolution. Plant Biosyst. 2011, 145 (Suppl. S1), 19–29. [Google Scholar] [CrossRef]
- Pott, R. Phytosociology: A modern geobotanical method. Plant Biosyst. 2011, 145, 9–18. [Google Scholar] [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; Studer, M.; Roudier, P.; Gonzalez, J.; Kozlowski, K.; Schubert, E.; et al. Cluster: Cluster Analysis Basics and Extensions. R Package Version 2.1.1. Available online: https://CRAN.R-project.org/package=cluster (accessed on 15 October 2022).
- McCune, B.; Mefford, M.J. Multivariate Analysis of Ecological Data, version 6; MjM Software PC-ORD: Gleneden Beach, OR, USA, 2011; pp. 1–34. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Tichý, L. JUICE, software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Tichý, L.; Chytrý, M. Statistical determination of diagnostic species for site groups of unequal size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Arrigoni, P.V. Contributo alla conoscenza della vegetazione del Monte Gennargentu in Sardegna. Boll. Soc. Sarda Sci. Nat. 1986, 25, 63–96. [Google Scholar]
- Arrigoni, P.V.; Di Tommaso, P.L.; Camarda, I.; Satta, V. La vegetazione dell’Azienda Forestale “Sa Pruna” (Dorgali—Sardegna centro-orientale). Parlatorea 1996, 1, 47–59. [Google Scholar]
- Camarda, I.; Lucchese, F.; Pignatti Wikus, E.; Pignatti, S. La vegetazione dell’area Pantaleo-Gutturu Mannu-Punta Maxia-Monte Arcosu nel Sulcis-Iglesiente (Sardegna sud-occidentale). Webbia 1995, 49, 141–177. [Google Scholar] [CrossRef] [Green Version]
- Blasi, C.; Frondoni, R. Frammenti di boschi igrofili nell’area tolfetano-cimina (Lazio). Ann. Bot. (Roma) 1996, 54, 171–182. [Google Scholar]
- Stanisci, A.; Presti, G.; Blasi, C. I boschi igrofili del Parco Nazionale del Circeo (Italia centrale). Ecol. Mediterr. 1998, 24, 73–88. [Google Scholar] [CrossRef]
- Ceschin, S.; Salerno, G. La vegetazione del basso corso del Fiume Tevere e dei suoi affluenti (Lazio, Italia). Fitosociologia 2008, 45, 39–74. [Google Scholar]
- Di Pietro, R.; Azzella, M.M.; Facioni, L. The Forest Vegetation of the Tolfa-Ceriti Mountains (Northern Latium—Central Italy). Hacquetia 2010, 9, 91–150. [Google Scholar] [CrossRef] [Green Version]
- Cutini, M.; Cancellieri, L.; Cioffi, M.T.; Licursi, C. Phytosociology and phytogeography of fragmented Alnus glutinosa forests in a Tyrrhenian district (Central Italy). Ecol. Mediterr. 2010, 36, 55–73. [Google Scholar] [CrossRef]
- Arrigoni, P.V. Flora e vegetazione della Macchia lucchese di Viareggio (Toscana). Webbia 1990, 44, 1–62. [Google Scholar] [CrossRef]
- Casini, S.; Chiarucci, A.; De Dominicis, V. Phytosociology and ecology of the Chianti woodlands. Fitosociologia 1995, 29, 115–136. [Google Scholar]
- Arrigoni, P.V.; Papini, P. La vegetazione del sistema fluviale Lima—Serchio (Toscana meridionale). Parlatorea 2003, 6, 95–129. [Google Scholar]
- Tomei, P.E.; Bertacchi, A.; Sani, A.; Consiglio, M. La vegetazione della Tenuta di San Rossore. Note esplicative della Carta della Vegetazione di San Rossore 1.10.000. Ente Parco Regionale Migliarino San Rossore. 2004. [Google Scholar]
- Viciani, D.; Gabellini, A.; Gonnelli, V.; De Dominicis, V. La vegetazione della riserva naturale Monti Rognosi (Arezzo, Toscana) ed i suoi aspetti di interesse botanico-conservazionistico. Atti Soc. Toscana Sci. Nat. Mem. 2004, 111, 27–42. [Google Scholar]
- Foggi, B.; Cartei, L.; Pignotti, L.; Signorini, M.A.; Viciani, D.; Dell’Olmo, L.; Menicagli, E. Il paesaggio vegetale dell’Isola d’Elba (Arcipelago Toscano). Studio fitosociologico e cartografico. Fitosociologia 2006, 43, 3–95. [Google Scholar]
- Landi, M.; Angiolini, C. Osmundo-Alnion woods in Tuscany (Italy): A phytogeographical analysis from a west European perspective. Plant Biosyst. 2010, 144, 93–110. [Google Scholar] [CrossRef]
- Lastrucci, L.; Paci, F.; Raffaelli, M. The wetland vegetation of the Natural Reserves and neighbouring stretches of the Arno river in the Arezzo province (Tuscany, Central Italy). Fitosociologia 2010, 47, 29–59. [Google Scholar]
- Sani, A.; Monacci, F.; Trimarchi, S.; Tomei, P.E. La vegetazione della Tenuta di Migliarino. Inter Nos. 2011, 2, 5–72. [Google Scholar]
- Francalancia, C.; Marconi, D. Ontanete ad Alnus glutinosa nei bacini dei fiumi Tronto, Tenna e Chienti. Fitosociologia 1994, 27, 91–95. [Google Scholar]
- Gafta, D.; Pedrotti, F. Tipificazione di due nuove associazioni forestali ripariali per la penisola italiana. Doc. Phytosociol. 1996, 15, 413–415. [Google Scholar]
- Taffetani, F.; Orlandini, S.; Zitti, S. Paesaggio vegetale di un’area pre-appenninica dell’Italia centrale: Il Bosco dei Monaci Bianchi nelle Marche (Italia). Fitosociologia 2009, 46, 27–47. [Google Scholar]
- Biondi, E.; Baldoni, M. La vegetazione del fiume Marecchia (Italia Centrale). Biogeographia 1994, 17, 51–87. [Google Scholar] [CrossRef]
- Rosati, L.; Di Pietro, R.; Blasi, C. La vegetazione forestale della Regione Temperata del “Flysch del Cilento” (Italia meridionale). Fitosociologia 2005, 42, 33–65. [Google Scholar]
- Barbagallo, C.; Brullo, S.; Furnari, F.; Longhitano, N.; Signorello, P. Studio fitosociologico e cartografia della vegetazione (1:25.000) del territorio di Serra S. Bruno (Calabria); CNR-AQ/1/227; Consiglio Nazionale delle Ricerche: Rome, Italy, 1982; pp. 1–19. [Google Scholar]
- Brullo, S.; Spampinato, G. Indagine sulle ripisilve della Calabria (Italia meridionale). Lazaroa 1997, 18, 105–151. [Google Scholar]
- Brullo, S.; Scelsi, F.; Spampinato, G. La vegetazione dell’Aspromonte. Studio fitosociologico; Laruffa Editore: Reggio Calabria, Italy, 2001; p. 372. [Google Scholar]
- Brullo, S.; Spampinato, G. La vegetazione dei corsi d’acqua della Sicilia. Boll. Acc. Gioenia Sci. Nat. 1990, 23, 119–252. [Google Scholar]
- Gamisans, J. La Végétation des Montagnes Corses. Ph.D. Thesis, University of Marseille, Marseille, France, 1975; 295p. [Google Scholar]
- Gamisans, J. Forêts marécageuses, ripisylves et fruticées ripicoles de Corse. Bull. Soc. Bot. Cent. Ouest 2013, 44, 383–452. [Google Scholar]
- Renaux, B.; Timbal, J.; Gauberville, C.; Thebaud, G.; Bardat, J.; Lalanne, A.; Royer, J.M.; Seytre, L. Contribution au Prodrome des végétations de France: Les Carpino betuli-Fagetea sylvaticae Jakucs 1967. Doc. Phytosociol. 2019, 11, 1–423. [Google Scholar]
- Poldini, L.; Sburlino, G. Lamio orvalae-Alnetum glutinosae Dakskobler 2016, nuova associazione ripariale per L’Italia settentrionale (Friuli Venezia Giulia, Veneto, Lombardia) con note sulle corrispondenti cenosi poste a sud del Po. Gortania 2020, 42, 5–21. [Google Scholar]
- Venanzoni, R.; Gigante, D. Contributo alla conoscenza della vegetazione degli ambienti umidi dell’Umbria (Italia). Fitosociologia 2000, 37, 13–63. [Google Scholar]
- Catorci, A.; Orsomando, E. Note Illustrative Della Carta Della Vegetazione Del Foglio Nocera Umbra (N. 312-Carta d’Italia I.G.M.—1:50000). Braun-Blanquetia 2001, 23, 7–108. [Google Scholar]
- Francalancia, C.; Orsomando, E. Les forets riveraines de la Valnerina (Italie Centrale). Colloq. Phytosociol. 1984, 9, 155–159. [Google Scholar]
- Lastrucci, L.; Landucci, F.; Gonnelli, V.; Barocco, R.; Foggi, B.; Venanzoni, R. The vegetation of the upper and middle River Tiber (Central Italy). Plant Sociol. 2012, 49, 29–48. [Google Scholar]
- Pirone, G. La vegetazione ripariale nei versanti nord-orientali del Gran Sasso d’Italia e dei Monti della Laga (Abruzzo, Italia). Fitosociologia 2000, 37, 65–86. [Google Scholar]
- Pirone, G.; Ciaschetti, G.; Frattaroli, A.R.; Corbetta, F. La vegetazione della Riserva Naturale Regionale “Lago di Serranella” (Abruzzo—Italia). Fitosociologia 2003, 40, 55–71. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Pignatti, S. Volume 1: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Volume 2: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Volume 3: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2018. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Volume 4: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole Edizioni Agricole di New Business Media srl: Milano, Italy, 2019. [Google Scholar]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimate of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Theurillat, J.P.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H. International Code of Phytosociological Nomenclature. Appl. Veg. Sci. 2020, 24, e12491. [Google Scholar]
- Andrew King, R.; Ferris, C. Chloroplast DNA phylogeography of Alnus glutinosa (L.) Gaertn. Mol. Ecol. 1998, 7, 1151–1161. [Google Scholar] [CrossRef]
- Havrdová, A.; Douda, J.; Krak, K.; Vít, P.; Hadincová, V.; Zákravský, P.; Mandák, B. Higher genetic diversity in recolonized areas than in refugia of Alnus glutinosa triggered by continent-wide lineage admixture. Mol. Ecol. 2015, 24, 4759–4777. [Google Scholar] [CrossRef] [PubMed]
- Mansion, G.; Rosenbaum, G.; Schoenenberger, N.; Bacchetta, G.; Rosselló, J.A.; Conti, E. Phylogenetic analysis informed by geological history supports multiple, sequential invasions of the Mediterranean Basin by the angiosperm family Araceae. Syst. Biol. 2008, 57, 269–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciandrello, S.; Guarino, R.; Minissale, P.; Spampinato, G. The endemic vascular flora of Peloritani Mountains (NE Sicily): Plant functional traits and phytogeographical relationships in the most isolated and fragmentary micro-plate of the Alpine orogeny. Plant Biosyst. 2015, 149, 838–854. [Google Scholar] [CrossRef] [Green Version]
- van Hinsbergen, D.J.J.; Torsvik, T.H.; Schmid, S.M.; Maţenco, L.C.; Maffione, M.; Vissers, R.L.M.; Gürer, D.; Spakman, W. Orogenic architecture of the Mediterranean region and kinematic reconstruction of its tectonic evolution since the Triassic. Gondwana Res. 2020, 81, 79–229. [Google Scholar] [CrossRef]
- Rivas-Martınez, S.; Dıaz, T.E.; Fernandez-Gonzalez, F.; Izco, J.; Loidi, J.; Lousa, M.; Penas, A. Vascular plant communities of Spain and Portugal. Addenda to the syntaxonomical checklist of 2001. Part II. Itinera Geobot. 2002, 15, 433–922. [Google Scholar]
- Berghen, V. Notes sur la végétation du sud-ouest de la France VIII. Les fourrés et les bois fangeux. Bull. Jard. Bot. Nat. Belg. 1971, 41, 283–395. [Google Scholar] [CrossRef]
- Bacchetta, G.; Bagella, S.; Biondi, E.; Farris, E.; Filigheddu, R.; Mossa, L. Vegetazione forestale e serie di vegetazione della Sardegna (con rappresentazione cartografica alla scala 1:350.000). Fitosociologia 2009, 46, 3–82. [Google Scholar]
- Brullo, S.; Marcenò, C. Contributo alla conoscenza della classe Quercetea ilicis in Sicilia. Notiz. Soc. Fitosociol. 1985, 19, 183–229. [Google Scholar]
- Sciandrello, S.; Giusso del Galdo, G.; Salmeri, C.; Minissale, P. Vicia brulloi (Fabaceae), a new species from Sicily. Phytotaxa 2019, 418, 57–78. [Google Scholar] [CrossRef]
- Brullo, S.; Cambria, S.; Crisafulli, A.; Tavilla, G.; Sciandrello, S. Taxonomic remarks on the Centaurea aeolica (Asteraceae) species complex. Phytotaxa 2021, 483, 9–24. [Google Scholar] [CrossRef]
- Kajba, D.; Gracan, J. EUFORGEN Technical guidelines for genetic conservation and use for black alder (Alnus glutinosa); International Plant Genetic Resources Institute: Rome, Italy, 2003; pp. 1–4. [Google Scholar]
- Petit, Y.; Hugot, L. Listes hiérarchisées des espèces végétales exotiques envahissantes et potentiellement envahissantes en Corse—Cadre méthodologique—Stratégie territoriale relative aux invasions biologiques végétales, Tome 1; Conservatoire botanique national de Corse/Office de l’environnement de la Corse: Corse, France, 2019; pp. 1–29. [Google Scholar]
- Reymann, J.; Panaïtis, C.; Bioret, F. Prodrome des végétations de Corse. Doc. Phytosociol. 2016, 3, 1–175. [Google Scholar]
- Fenu, G.; Fois, M.; Cañadas, E.M.; Bacchetta, G. Using endemic-plant distribution, geology and geomorphology in biogeography: The case of Sardinia (Mediterranean Basin). Syst. Biodivers. 2014, 12, 181–193. [Google Scholar] [CrossRef]
- Bacchetta, G.; Biondi, E.; Farris, E.; Filigheddu, R.; Mossa, L. A phytosociological study of the deciduous oak woods of Sardinia (Italy). Fitosociologia 2004, 41, 53–65. [Google Scholar]
- Fantini, S.; Fois, M.; Casula, P.; Fenu, G.; Calvia, G.; Bacchetta, G. Structural heterogeneity and old-growthness: A first regional-scale assessment of Sardinian forests. Ann. For. Res. 2020, 63, 103–120. [Google Scholar] [CrossRef]
- Clement, B.; Touffet, J. Contribution à l’étude des groupements préforestiers issus des landes méso-higrophiles, des tourbières et des prairies marécageuses de Bretagne. Colloq. Phytosociol. 1979, 8, 229–237. [Google Scholar]
- Noirfalise, A.; Dethioux, M. Synopsis des forêts alluviales de Belgique. Colloq. Phytosociol. 1984, 9, 217–226. [Google Scholar]
- Landi, M.; Frignani, F.; Bonini, I.; Casini, F.; Saveri, C.; De Dominicis, V.; Angiolini, C. Flora and vegetation in the catchment area of the stream “La Bolza” in the Merse valley (Siena, southern Tuscany). Webbia 2009, 64, 187–212. [Google Scholar] [CrossRef]
- Landi, M.; Angiolini, C. Contributo alla conoscenza della distribuzione di Osmunda regalis L. in Toscana. Inf. Bot. Ital. 2007, 39, 113–122. [Google Scholar]
- De Dominicis, V.; Casini, S. Memoria illustrativa per la carta della vegetazione della Val di Farma (Colline Metallifere). Atti Soc. Toscana Sci. Nat. Mem. B 1979, 86, 1–36. [Google Scholar]
- Rivas-Martınez, S.; Fernandez-Gonzalez, F.; Loidi, J.; Lousa, M.; Penas, A. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobot. 2001, 14, 5–341. [Google Scholar]
- Spada, F. Primi lineamenti della vegetazione del comprensorio tolfetano-cerite. Accad. Naz. Lincei (Roma) 1977, 227, 37–50. [Google Scholar]
- Montelucci, G. Note preliminari sulla flora e sulla vegetazione delle cerrete di Manziana e di Canale Monterano. Acc. Naz. Lincei (Roma) 1977, 227, 51–74. [Google Scholar]
- Lucchese, F.; Pignatti, S. Sguardo sulla vegetazione del Lazio marittimo. Accad. Naz. Lincei (Roma) 1990, 264, 5–48. [Google Scholar]
- Petriccione, B.; Pani, F. Primo contributo alla conoscenza sintassonomica dei boschi igrofili nel Lazio. Ann. Bot. (Roma) 1990, 48, 125–153. [Google Scholar]
- Blasi, C.; Cutini, M.; Fortini, P.; Di Marzio, P. I boschi caducifogli del comprensorio Barbarano Romano-Canale Monterano (Lazio settentrionale). Ann. Bot. (Roma) 1995, 51, 279–296. [Google Scholar]
- Montanari, C.; Gentile, S. Ricerche sulla vegetazione arbustiva e arborea di greto nei fiumi Vara e Magra (Liguria orientale). Not. Fitosoc. 1979, 14, 17–40. [Google Scholar]
- Abbate, G.; Blasi, C.; Fascetti, S.; Michetti, L.; Filesi, L. La vegetazione del parco suburbano Valle del Treja; Tip. Borgia: Regione Lazio, Italy; Dip Biol. Vegetale Univ. “La Sapienza”: Roma, Italy, 1990; pp. 1–32. [Google Scholar]
- Biondi, E.; Baldoni, M. La vegetazione del fiume Esino (Marche-Italia centrale). Stud. Bot. 1993, 11, 209–257. [Google Scholar]
- Tchou, Y.T. Études écologiques et phytosociologiques sur les fôrets riveraines du Bas-Languedoc (Populetum albae). Vegetatio 1946, 1, 2–28. [Google Scholar]
- Oberdorfer, E. Der europäische Auenwald. Beitr. Nat. Forsch. Südwestdtschl. Beih. 1953, 12, 23–69. [Google Scholar]
- Navarro, C. Contribución al estudio de la flora y vegetación del Duranguesado y la Busturia (Vizcaya). Ph.D. Thesis, Publ. Univ. Complutense de Madrid, Madrid, Spain, 1982; pp. 1–398. [Google Scholar]
- Fukarek, P. Forest plant communites of Yugoslavia. In Proc. Book of Second Congress of Ecologists of Yugoslavia; Zadar-Plitvice, 1979; Volume 1, pp. 55–69. [Google Scholar]
- Dakskobler, I. Phytosociological analysis of riverine forests in the Vipava and Reka Valleys (Southwestern Slovenia). Folia Biol. Geol. 2016, 57, 5–61. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, G.; Bernardo, L.; Passalacqua, N.G. Carta delle serie di vegetazione della Regione Calabria. In La Vegetazione d’Italia; Blasi, C., Ed.; Palombi Editori: Roma, Italy, 2010. [Google Scholar]
- Spampinato, G.; Cameriere, P.; Caridi, D.; Crisafulli, A. Carta della biodiversità del Parco Nazionale dell’Aspromonte. Quad. Bot. Ambient. Appl. 2008, 19, 3–36. [Google Scholar]
- Spampinato, G. Phytocoenotic diversity in Southern Italy. Bocconea 2009, 23, 33–49. [Google Scholar]
- Maiorca, G.; Spampinato, G.; Crisafulli, A.; Cameriere, P. Flora vascolare e vegetazione della Riserva Naturale Regionale “Foce del Fiume Crati” (Calabria, Italia meridionale). Webbia 2007, 62, 121–174. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rel. Numbers | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 |
| Rel. Numbers Cluster | 219 | 220 | 221 | 222 | 223 | 226 | 227 | 228 | 229 | 230 | 231 | 232 | 233 | 234 | 235 | 236 | 237 | 238 | 239 | 240 | 241 | 242 | 243 | 244 | 245 |
| Rel. Numbers from the literature | 12 | 13 | 16 | 14 * | 15 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Area (m2) | 50 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 150 | 150 | 150 | 150 | 100 | 100 | 150 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Aspect | - | - | - | - | - | - | - | - | NE | NW | SE | SE | SE | E | E | E | E | SE | SE | NE | NE | SE | E | E | E |
| Slope (°) | - | - | - | - | - | - | - | - | 8 | 5 | 10 | 10 | 10 | 10 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Tree cover (%) | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Shrum cover (%) | - | - | - | - | - | 20 | 40 | 20 | 60 | 50 | 50 | 60 | 60 | 50 | 50 | 60 | 50 | 70 | 60 | 40 | 60 | 50 | 80 | 70 | 80 |
| Herbaceous cover (%) | - | - | - | - | - | - | - | - | 40 | 30 | 40 | 40 | 40 | 40 | 50 | 50 | 50 | 30 | 40 | 60 | 40 | 50 | 30 | 40 | 30 |
| Altitude (m s.l.m.) | 380 | 400 | 420 | 450 | 490 | 465 | 466 | 467 | 270 | 285 | 430 | 470 | 490 | 400 | 280 | 442 | 455 | 460 | 458 | 600 | 600 | 576 | 575 | 572 | 570 |
| Richness floristic | 7 | 18 | 12 | 11 | 12 | 11 | 20 | 19 | 28 | 20 | 17 | 26 | 23 | 25 | 29 | 27 | 28 | 27 | 29 | 27 | 29 | 30 | 27 | 24 | 27 |
| Char. Ass. | |||||||||||||||||||||||||
| Platanus orientalis | 2 | 2 | 2 | 2 | 1 | 1 | 2 | 3 | 1 | 1 | 1 | 1 | 2 | . | 1 | 1 | 2 | 1 | + | 1 | 2 | + | 1 | 2 | 1 |
| Char. Alno glutinosae-Populetea albae and Populetalia albae | |||||||||||||||||||||||||
| Alnus glutinosa | 5 | 4 | 5 | 4 | 5 | 5 | 4 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Polystichum setiferum | . | . | . | . | . | . | . | . | . | . | + | 1 | + | 1 | 1 | 1 | + | + | + | 2 | 1 | 1 | 1 | 1 | 1 |
| Brachypodium sylvaticum subsp. sylvaticum | 2 | 3 | 3 | . | 1 | . | . | . | 2 | 1 | 2 | 2 | 2 | 2 | 3 | 2 | 3 | 2 | 1 | 2 | 1 | 1 | + | + | 1 |
| Carex pendula | . | . | + | . | . | . | . | . | 1 | 1 | . | 1 | + | . | 1 | + | + | 1 | + | + | + | + | 1 | + | + |
| Sambucus nigra | . | . | . | . | . | . | . | . | 1 | 1 | . | 1 | 2 | 1 | 1 | 2 | 1 | 1 | + | + | + | . | . | . | . |
| Populus nigra | . | 1 | 1 | + | . | 1 | 2 | 1 | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . |
| Salix alba | . | . | . | . | . | . | 1 | . | 1 | 1 | . | . | . | 1 | 2 | . | . | . | . | . | . | . | . | . | . |
| Ulmus minor | . | . | . | . | . | . | . | . | . | . | . | 1 | . | 1 | . | 1 | 1 | . | 1 | . | . | . | . | . | . |
| Carex remota | . | . | . | . | . | + | + | 1 | . | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . |
| Salix gussonei | 1 | + | 2 | 3 | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Equisetum telmateia | . | . | . | . | . | . | . | . | . | . | 1 | . | . | 1 | 1 | . | . | . | . | . | . | . | . | . | . |
| Char. Osmundo regalis-Alnion glutinosae | |||||||||||||||||||||||||
| Athyrium filix-femina | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | + | + | . | . | 1 | . | + | . | . | . |
| Osmunda regalis | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Char. Hyperico hircini–Alnenion glutinosae | |||||||||||||||||||||||||
| Hypericum hircinum | . | + | 2 | . | + | . | . | . | + | 1 | 2 | 2 | 1 | 1 | 1 | 2 | 1 | 3 | 1 | 1 | 2 | 2 | 1 | 1 | 2 |
| Lamium flexuosum | . | . | . | . | . | 1 | 1 | 1 | 1 | 1 | . | + | 1 | + | . | 2 | 1 | + | + | 1 | + | + | + | 1 | + |
| Fraxinus ornus | . | . | . | . | . | . | . | . | . | . | . | 1 | 1 | 1 | 1 | + | . | . | . | . | + | 1 | 1 | 1 | 1 |
| Ficus carica | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 1 | 1 | 1 | 1 | 2 | 1 | . | 1 | + | . | . | 1 |
| Other species | |||||||||||||||||||||||||
| Rubus ulmifolius | . | 2 | . | + | 1 | 1 | 2 | 1 | 2 | 1 | 2 | 2 | 2 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 3 | 2 | 2 | 2 | 3 |
| Hedera helix subsp. helix | . | . | . | . | + | 1 | + | + | 1 | 1 | 1 | 1 | 2 | + | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 2 | 2 | 3 | 2 |
| Eupatorium cannabinum | 2 | 1 | . | 1 | . | . | . | . | + | + | 2 | 1 | 2 | 1 | 1 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | + | + |
| Clematis vitalba | . | . | + | . | . | + | + | . | + | 1 | . | 1 | 1 | . | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 2 | 1 | 1 | + |
| Euphorbia characias | . | . | . | + | + | . | + | + | . | . | 1 | 1 | 1 | + | + | + | 1 | + | + | + | + | . | . | + | + |
| Mentha suaveolens | . | . | . | . | . | . | . | . | 1 | + | 2 | 1 | 1 | + | 1 | . | + | + | + | 1 | + | + | + | + | + |
| Holcus lanatus subsp. lanatus | . | . | . | . | . | . | . | . | 1 | + | . | . | + | + | + | + | + | 1 | 1 | 1 | + | 1 | 1 | + | + |
| Mycelis muralis | . | . | . | . | . | . | + | + | . | + | . | . | . | . | . | + | 1 | + | 1 | 1 | 1 | + | + | + | + |
| Viola alba subsp. dehnhardtii | . | . | . | + | + | . | + | + | 1 | . | + | . | . | . | + | . | . | . | . | 1 | 1 | 1 | + | 1 | + |
| Dioscorea communis | . | + | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 1 | 1 | 1 | 1 | + | . | + | + | + | + |
| Pteridium aquilinum subsp. aquilinum | . | . | . | . | . | . | . | . | . | . | + | + | . | 1 | + | + | 1 | + | + | 1 | 1 | + | . | . | . |
| Equisetum arvense | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | + | + | 1 | + | 1 | 1 | + | + |
| Asparagus acutifolius | . | . | . | . | . | . | + | + | + | . | . | 1 | 1 | . | . | + | + | . | + | . | . | + | . | . | + |
| Anthriscus nemorosa | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 | + | 1 | 2 | 1 | 1 | + | 1 | + |
| Agrostis castellana | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | 2 | 1 | 1 | 3 | 2 | 1 | + | + | + |
| Asplenium onopteris | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | + | + | + | + | + | + | + |
| Tussilago farfara | . | . | + | . | . | . | . | . | + | . | . | . | . | . | . | + | + | + | . | 1 | + | + | + | . | . |
| Rubia peregrina | . | . | . | . | . | . | 1 | + | . | . | . | . | . | . | . | . | . | . | . | 1 | + | + | + | 1 | + |
| Adiantum capillus-veneris | . | . | . | . | . | . | . | . | . | . | 1 | 1 | + | . | 1 | 1 | 1 | 1 | + | . | . | . | . | . | . |
| Equisetum ramosissimum | 1 | 1 | 1 | . | + | . | . | . | + | . | . | . | 1 | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Allium subhirsutum | . | + | . | . | . | . | . | 1 | 1 | 1 | . | 1 | . | + | 1 | . | . | . | . | . | . | . | . | . | . |
| Scutellaria columnae subsp. gussonei | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | 1 | + | . | + | . | + | . | + |
| Agrostis stolonifera | . | . | . | . | . | . | . | . | 1 | . | 1 | 1 | + | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Clinopodium arundanum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | + | + | + | . | . |
| Ruscus aculeatus | . | . | . | . | . | . | . | . | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | + | + |
| Nerium oleander | . | . | . | . | . | 2 | 1 | 2 | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Allium triquetrum | . | . | . | . | . | . | . | . | 1 | . | . | 1 | 1 | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Salix purpurea | . | 1 | . | 1 | 1 | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Parietaria judaica | . | . | . | . | . | . | . | . | . | . | 1 | 1 | . | 2 | + | . | . | . | . | . | . | . | . | . | . |
| Convolvulus silvaticus | . | . | . | . | . | . | . | . | 1 | . | . | 1 | . | 1 | 1 | . | . | . | . | . | . | . | . | . | . |
| Dittrichia viscosa | 1 | 1 | . | 1 | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Torilis africana | . | . | + | . | . | + | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Epilobium parviflorus | . | . | . | . | . | . | . | . | . | . | + | + | . | 2 | + | . | . | . | . | . | . | . | . | . | . |
| Arum italicum | . | . | . | . | . | . | . | . | . | + | . | . | . | + | 1 | . | . | . | . | . | . | . | . | . | . |
| Prunella vulgaris subsp. vulgaris | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | + | . | . |
| Geranium versicolor | . | . | . | . | . | 1 | + | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Pulicaria dysenterica | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | + | . | . | . | . | . | . |
| Stachys sylvatica | . | . | . | . | . | . | 1 | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ajuga reptans | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Viola reichenbachiana | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Galium aparine | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Angelica sylvestris subsp. sylvestris | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cyclamen repandum subsp. repandum | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ficaria verna | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Euphorbia meuselii | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Smilax aspera | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Poa trivialis | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Rosa canina | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Cirsium creticum subsp. triumfettii | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Laurus nobilis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . |
| Geranium purpureum | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Rel. Numbers | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27* | 28 | 29 | 30 | 31 |
| Rel. Numbers Cluster | 310 | 311 | 312 | 313 | 314 | 316 | 324 | 332 | 353 | 354 | 355 | 356 | 357 | 358 | 359 | 360 | 361 | 362 | 363 | 364 | 365 | 366 | 367 | 368 | 369 | 370 | 371 | 372 | 373 | 374 | 375 |
| Area (m2) | 150 | 40 | 100 | 100 | 50 | 100 | 100 | 50 | 100 | 200 | 100 | 20 | 150 | 100 | 100 | 50 | 30 | 100 | 50 | 50 | 50 | 50 | 30 | 50 | 80 | 50 | 150 | 100 | 50 | 80 | 80 |
| Aspect | W | W | NE | NW | NW | NE | - | - | - | - | - | - | W | - | - | - | - | - | N | N | - | - | - | W | E | - | - | SE | - | - | |
| Slope (°) | 35 | 25 | 25 | 20 | 15 | 20 | 0 | 0 | 5 | 0 | 0 | 0 | 35 | 0 | 5 | 0 | 5 | 0 | 15 | 15 | 5 | 5 | 5 | 0 | 10 | 20 | 0 | 0 | 10 | 0 | 0 |
| Cover (%) | 95 | 90 | 95 | 90 | 85 | 90 | 100 | 90 | 80 | 80 | 50 | 70 | 95 | 95 | 95 | 80 | 95 | 95 | 95 | 95 | 100 | 95 | 100 | 80 | 95 | 95 | 95 | 90 | 90 | 95 | 80 |
| Altitude (m s.l.m.) | 703 | 700 | 862 | 598 | 711 | 843 | 813 | 850 | 839 | 861 | 747 | 709 | 649 | 783 | 739 | 729 | 709 | 521 | 539 | 559 | 639 | 671 | 659 | 629 | 589 | 587 | 689 | 609 | 531 | 533 | 549 |
| Richness floristic | 18 | 20 | 18 | 10 | 9 | 19 | 26 | 25 | 35 | 30 | 16 | 11 | 28 | 24 | 18 | 18 | 14 | 24 | 17 | 19 | 16 | 16 | 18 | 12 | 19 | 16 | 33 | 26 | 28 | 18 | 29 |
| Char. Ass. | |||||||||||||||||||||||||||||||
| Geranium nodosum | 2 | 2 | 1 | + | . | 2 | 2 | + | 1 | 1 | . | 2 | 1 | 1 | 2 | 2 | . | r | . | . | + | + | r | . | + | r | 2 | 1 | 1 | + | r |
| Cardamine heptaphylla | . | . | . | . | . | . | . | r | r | + | + | . | . | + | + | + | . | . | + | . | . | . | . | . | . | . | r | . | . | . | . |
| Char. Ligustro vulgaris-Alnion glutinosae | |||||||||||||||||||||||||||||||
| Circaea lutetiana | 1 | + | 1 | . | . | + | 2 | 1 | + | + | r | 1 | + | . | . | r | + | 1 | 2 | + | . | . | 1 | . | + | + | 2 | 2 | . | . | + |
| Aegopodium podagraria | + | r | . | . | . | . | . | . | . | + | . | . | r | + | 2 | . | . | . | + | . | . | . | r | . | + | . | 1 | r | r | + | + |
| Geum urbanum | + | + | . | . | . | . | + | + | . | . | . | . | + | . | . | . | + | + | + | + | . | . | . | . | . | . | 1 | . | + | + | + |
| Rubus caesius | 2 | 1 | . | . | . | + | . | r | . | . | . | 1 | . | . | . | . | . | 2 | 3 | 2 | . | . | . | . | . | + | . | . | + | r | + |
| Sambucus nigra | + | . | . | . | 1 | 2 | + | + | . | . | . | 2 | 1 | . | . | . | . | + | . | . | . | . | . | . | . | . | 1 | 1 | . | . | . |
| Alliaria petiolata | r | . | . | . | . | 1 | . | . | . | r | . | . | + | . | . | . | . | 1 | + | . | + | . | . | . | . | . | . | . | . | . | r |
| Stachys sylvatica | . | . | 2 | . | . | r | 1 | . | r | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cardamine impatiens | . | . | . | . | . | . | + | . | + | + | r | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Chaerophyllum temulum | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . | . | . | . | . | . |
| Char. Alno glutinosae-Populetea albae and Populetalia albae | |||||||||||||||||||||||||||||||
| Alnus glutinosa | 5 | 4 | 4 | 5 | 4 | 4 | 5 | 5 | 4 | 4 | 3 | 3 | 4 | 3 | 3 | 4 | 5 | 4 | 4 | 5 | 5 | 5 | 5 | 4 | 4 | 4 | 4 | 4 | 4 | 5 | 3 |
| Brachypodium sylvaticum subsp. sylvaticum | + | 3 | . | . | + | . | . | . | 1 | . | . | 1 | 2 | . | . | . | . | 1 | . | + | 2 | . | . | . | . | 1 | r | . | 1 | 1 | 2 |
| Carex pendula | + | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | + | . | 1 | + | + |
| Solanum dulcamara | . | . | . | . | . | r | + | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . |
| Carex remota | . | . | . | . | . | . | . | . | 2 | 1 | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Salix alba | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | 2 | . | . |
| Fraxinus angustifolia subsp. oxycarpa | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Saponaria officinalis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . |
| Other species | |||||||||||||||||||||||||||||||
| Melica uniflora | + | 1 | 1 | 1 | . | + | 1 | 1 | + | 1 | + | 2 | + | 1 | 1 | . | . | 1 | + | . | . | + | 1 | r | r | + | 1 | + | r | . | 1 |
| Hedera helix subsp. helix | . | . | . | 2 | . | . | . | . | . | . | 1 | 2 | + | + | . | + | + | . | . | . | . | 1 | 1 | r | 1 | 1 | 1 | 1 | + | + | 1 |
| Geranium robertianum | . | + | + | . | . | + | . | 3 | + | . | + | . | + | . | + | 1 | . | . | . | . | r | . | r | . | r | r | + | + | + | . | + |
| Corylus avellana | . | . | . | . | 1 | . | . | + | . | . | 1 | . | 1 | 1 | 1 | . | + | . | . | 1 | 2 | 1 | 3 | 1 | 3 | 3 | 1 | 1 | . | . | . |
| Dryopteris filix-mas | r | . | 1 | 1 | 1 | + | + | . | . | r | . | . | . | . | + | . | + | . | . | 1 | . | + | . | . | . | . | 1 | 1 | . | r | + |
| Viola reichenbachiana | . | . | + | . | . | . | + | r | . | + | . | . | . | 1 | . | r | . | . | . | . | . | . | + | r | + | r | r | r | r | . | + |
| Ranunculus lanuginosus | 2 | + | . | . | . | . | + | 1 | 1 | + | r | . | 1 | + | . | . | . | . | . | . | r | r | + | + | . | . | . | . | . | . | . |
| Mycelis muralis | + | . | + | . | . | + | + | 1 | . | . | . | . | . | . | r | r | . | . | r | . | . | . | . | . | . | r | + | r | r | . | . |
| Acer pseudoplatanus | + | 1 | . | . | . | . | . | . | r | + | + | . | 2 | + | . | + | . | . | . | . | 2 | . | . | . | . | . | . | 1 | 2 | . | . |
| Clematis vitalba | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | + | . | r | r | . | . | . | + | + | r | + | r |
| Euphorbia dulcis | . | + | . | . | . | . | . | . | r | . | r | . | . | r | r | r | . | + | . | . | . | . | . | . | r | . | r | + | . | . | . |
| Saxifraga rotundifolia | + | . | + | + | . | . | . | . | r | r | . | . | r | + | r | . | . | . | . | . | . | . | . | . | . | r | . | + | . | . | . |
| Urtica dioica | . | . | + | . | + | + | . | . | . | + | . | . | . | . | . | 1 | + | + | . | . | . | . | . | . | . | . | + | . | . | . | + |
| Fraxinus ornus | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . | . | . | + | + | . | . | . | . | . | r | . | + | r | + |
| Equisetum arvense | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | 1 | r | + | 1 | + | . | + | . | . | . | r |
| Rubus hirtus | . | . | 1 | . | 3 | . | . | . | . | . | . | . | + | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | + | + | + | + |
| Acer campestre | . | 2 | . | . | . | . | . | 1 | . | . | . | . | + | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | 1 | 1 | + |
| Lunaria rediviva | . | . | . | . | . | . | . | . | r | . | . | . | . | . | r | + | + | + | 1 | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Cornus sanguinea | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 2 | . | . | . | . | 2 | . | . | . | + | 1 | + |
| Chaerophyllum hirsutum (incl. C. calabrum) | . | . | . | . | . | 2 | r | . | 2 | . | r | . | . | . | . | . | + | . | . | r | . | . | . | . | . | . | . | . | . | . | . |
| Cardamine bulbifera | . | . | . | . | . | . | . | 1 | . | . | r | . | + | + | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Fagus sylvatica subsp. sylvatica | . | . | . | . | . | . | . | . | . | 1 | . | . | . | 3 | 3 | + | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | r |
| Petasites albus | . | . | 2 | . | 1 | + | . | . | r | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . |
| Stellaria nemorum subsp. montana | . | . | . | . | . | . | 1 | . | . | 1 | 1 | . | r | 1 | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Crataegus monogyna | . | 1 | + | . | . | . | . | . | . | . | . | . | + | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . |
| Castanea sativa | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | 1 | r | . | r | . | . | . | r |
| Anemonoides nemorosa | . | r | . | . | . | . | . | . | r | + | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Poa sylvicola | . | . | . | . | . | . | + | . | + | r | . | . | r | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . |
| Lolium giganteum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 | + | + | . | . | . | . | . | . | + | . | . | . |
| Campanula trachelium | . | . | r | + | . | . | . | . | r | . | . | . | r | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Oxalis acetosella | . | . | . | 1 | . | . | + | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | + | r | . | . | . | . | . | . | . |
| Polystichum setiferum | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | 1 | . | . | . | + | . | . | . | . | 1 |
| Viola alba subsp. dehnhardtii | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | r | r | r | . |
| Athyrium filix-femina | . | . | . | . | . | . | 1 | . | + | . | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Equisetum telmateia | . | + | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . |
| Carex sylvatica | . | . | . | . | . | . | + | + | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . |
| Carpinus betulus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | 1 | . | . | 1 | . | . | . | . | 2 |
| Primula vulgaris | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | r | . | . | . | r | . | . |
| Festuca heterophylla | . | . | . | . | . | . | . | . | + | + | . | . | . | + | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . |
| Ranunculus repens | . | . | . | . | . | . | + | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | + |
| Rubus idaeus subsp. idaeus | . | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . | . | . | 1 | + | . | . | . |
| Euphorbia amygdaloides | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | + | + | + |
| Helleborus viridis subsp. bocconei | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | r | . | r | . | . | . | . | . | r |
| Impatiens parviflora | . | + | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | r | . | . | + | . | . | . | . | . | . | . | . | . | . |
| Galeopsis speciosa | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | r | . | . | . | . | . | r | . | . |
| Senecio ovatus | . | . | . | . | r | . | + | . | r | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Veronica urticifolia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | r | . | . | . | r | . | . | . |
| Lamium galeobdolon | . | . | . | . | . | . | . | . | . | . | . | 1 | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Mercurialis perennis | . | . | . | . | . | + | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Robinia pseudoacacia | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | + | . | . | . | . | . | . | . | . | 1 | . | . | . |
| Poa nemoralis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | + | . | . | . | . | . | + | . | . | . | . |
| Thalictrum aquilegiifolium subsp. aquilegiifolium | . | . | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . |
| Parietaria officinalis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | + | . | . | . | . | . | + | . | . | . | . | . |
| Scrophularia nodosa | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | r |
| Asarum europaeum | . | . | . | 2 | . | . | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Dactylis glomerata | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | 1 | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Polygonatum multiflorum | . | . | . | . | . | . | . | . | r | . | . | . | . | + | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . |
| Myosotis sylvatica | . | . | . | . | . | . | . | . | + | + | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Pteridium aquilinum subsp. aquilinum | . | . | . | . | . | . | . | . | + | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Dioscorea communis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | + | . | . | . | . |
| Symphytum tuberosum subsp. angustifolium | . | . | . | . | . | + | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ficaria verna | . | . | . | . | . | . | . | . | + | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Solidago virgaurea | r | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Salvia glutinosa | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . |
| Arctium minus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | + | . | . | . | . | . | . | . | . | . | . | . |
| Moehringia trinervia | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Juglans regia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | 2 | . | . | . | . |
| Asplenium trichomanes subsp. quadrivalens | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . |
| Gentiana asclepiadea | . | . | . | . | . | . | . | . | + | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Rosa arvensis | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . |
| Sedum cepaea | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | r | . | . | . | . | . | . | . | . | . | . | . |
| Rubus ulmifolius | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ficus carica | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . |
| Agrostis stolonifera | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Prunella vulgaris subsp. vulgaris | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Lathyrus venetus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . |
| Rumex sanguineus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ajuga reptans | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . |
| Arctium lappa | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ostrya carpinifolia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . |
| Dactylorhiza maculata | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Rosa sempervirens | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Lapsana communis | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Lonicera etrusca | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Prenanthes purpurea | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Prunus avium | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Clinopodium vulgare | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . |
| Fragaria vesca subsp. vesca | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . |
| Emerus major | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . |
| Chelidonium majus | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Daphne laureola | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Rumex conglomeratus | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Bromopsis ramosa | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . |
| Cardamine chelidonia | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Populus tremula | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Pulmonaria hirta | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Impatiens noli-tangere | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . |
| Adoxa moschatellina subsp. moschatellina | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Myosotis scorpioides | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Rel. Numbers | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 * | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 |
| Rel. Numbers Cluster | 315 | 317 | 318 | 319 | 320 | 321 | 322 | 323 | 325 | 326 | 327 | 328 | 329 | 330 | 331 | 333 | 334 | 335 | 337 | 339 | 340 | 343 | 344 | 345 | 346 | 347 | 348 | 349 |
| Area (m2) | 100 | 50 | 50 | 50 | 30 | 100 | 50 | 50 | 20 | 50 | 45 | 80 | 100 | 50 | 100 | 50 | 50 | 300 | 100 | 70 | 70 | 200 | 80 | 60 | 200 | 100 | 100 | 100 |
| Aspect | W | W | NE | N | E | - | - | - | E | SE | SE | - | - | - | - | - | - | N | - | - | NE | - | - | - | NW | - | - | - |
| Slope (°) | 20 | 5 | 25 | 3 | 30 | 0 | 0 | 0 | 5 | 10 | 45 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 30 | 32 | 3 | 3 | 6 | 2 | 2 | 2 |
| Cover total (%) | 100 | 95 | 90 | 85 | 90 | 95 | 85 | 100 | 100 | 100 | 90 | 90 | 90 | 90 | 90 | 90 | 100 | 100 | 85 | 100 | 95 | 100 | 95 | 90 | 100 | 100 | 90 | 100 |
| Altitude (m s.l.m.) | 240 | 265 | 145 | 283 | 287 | 631 | 173 | 286 | 263 | 263 | 235 | 540 | 540 | 600 | 560 | 740 | 550 | 620 | 330 | 251 | 265 | 520 | 440 | 445 | 630 | 320 | 340 | 380 |
| Richness floristic | 25 | 23 | 28 | 25 | 25 | 22 | 25 | 16 | 21 | 30 | 39 | 31 | 32 | 34 | 33 | 29 | 32 | 25 | 23 | 36 | 31 | 32 | 31 | 30 | 46 | 39 | 37 | 28 |
| Char. Ass. | ||||||||||||||||||||||||||||
| Geranium nodosum | . | 1 | + | + | + | . | + | 1 | 1 | 2 | + | 1 | + | 1 | 1 | 2 | 1 | . | r | + | 1 | 2 | 1 | 1 | 2 | . | + | + |
| Cornus sanguinea | 1 | . | 1 | . | . | . | . | 1 | 2 | 1 | + | 1 | + | + | 1 | . | 2 | . | 3 | 1 | + | + | 1 | 1 | . | + | 1 | 1 |
| Char. Ligustro vulgaris-Alnion glutinosae | ||||||||||||||||||||||||||||
| Aegopodium podagraria | . | . | . | . | 1 | . | + | . | 3 | 3 | 2 | + | + | 2 | 3 | 3 | 2 | . | 2 | . | 3 | . | 2 | 1 | + | 1 | . | 1 |
| Circaea lutetiana | 1 | . | + | . | + | 1 | + | 1 | 1 | 2 | + | 1 | + | . | 1 | + | . | 1 | . | . | . | 1 | . | . | 1 | . | + | . |
| Rubus caesius | . | . | 1 | . | . | . | . | . | . | 1 | . | 3 | 1 | 1 | 1 | . | . | . | 2 | . | . | 2 | 1 | . | . | + | + | 2 |
| Geum urbanum | . | + | . | . | + | + | . | . | . | + | + | . | . | . | + | . | . | + | . | + | . | . | . | . | + | . | + | . |
| Petasites hybridus subsp. hybridus | . | . | . | . | . | . | . | . | . | . | . | + | 1 | . | . | . | . | . | . | . | r | . | 1 | 2 | + | 1 | + | 2 |
| Alliaria petiolata | . | . | . | . | . | + | . | . | . | + | . | . | . | . | 2 | . | + | . | . | + | r | + | . | . | . | . | + | . |
| Stachys sylvatica | . | . | . | + | . | + | . | . | . | . | . | 1 | . | 1 | . | + | . | . | . | . | . | + | . | . | 1 | . | . | . |
| Cardamine impatiens | . | + | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | 1 | . | . | . | . | . | . | 2 | . | . | . |
| Chaerophyllum temulum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | + | + | + | + | . |
| Char. Alno glutinosae-Populetea albae and Populetalia albae | ||||||||||||||||||||||||||||
| Alnus glutinosa | 4 | 4 | 5 | 5 | 4 | 4 | 5 | 4 | 4 | 4 | 4 | 4 | 5 | 4 | 5 | 5 | 5 | 5 | 5 | 3 | 5 | 5 | 5 | 5 | 4 | 4 | 4 | 4 |
| Brachypodium sylvaticum subsp. sylvaticum | + | . | + | . | . | . | + | . | . | . | 2 | + | + | 2 | 1 | . | 2 | + | 1 | . | + | + | 1 | 1 | . | 1 | + | + |
| Carex pendula | 1 | . | 1 | + | + | . | + | 1 | . | . | + | . | + | + | 1 | + | + | . | . | . | . | . | 1 | 1 | 1 | 2 | + | . |
| Sambucus nigra | 1 | 1 | 1 | 1 | . | 1 | 2 | + | . | 2 | . | . | . | . | 1 | . | . | . | r | 1 | + | 1 | . | . | . | + | 1 | . |
| Solanum dulcamara | R | . | . | . | . | . | + | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Carex remota | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . |
| Salix alba | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | 1 | 1 | . | . |
| Populus nigra | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | 2 |
| Fraxinus angustifolia subsp. oxycarpa | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . |
| Saponaria officinalis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . |
| Other species | ||||||||||||||||||||||||||||
| Hedera helix subsp. helix | 3 | 2 | 2 | 2 | 2 | + | 1 | 3 | 1 | 2 | 1 | . | . | + | + | . | 2 | + | 3 | 2 | 1 | + | 1 | 3 | 1 | 2 | 3 | + |
| Clematis vitalba | 1 | + | 1 | 1 | 1 | 1 | + | + | 1 | 1 | . | + | 1 | . | . | . | 1 | 1 | 1 | + | . | 1 | 1 | 1 | + | + | + | 1 |
| Melica uniflora | + | + | + | + | . | + | + | + | 1 | 1 | + | + | + | . | + | 1 | . | . | . | r | . | + | . | . | 2 | + | 1 | 1 |
| Corylus avellana | 1 | 1 | . | 1 | 2 | . | . | 1 | + | . | 1 | + | + | + | . | . | . | . | r | r | 1 | 1 | 1 | . | 2 | 1 | 2 | 1 |
| Acer campestre | . | + | + | . | + | . | . | . | + | . | + | r | + | 1 | + | + | + | . | 3 | 1 | 2 | . | . | + | . | + | 1 | + |
| Fraxinus ornus | . | 1 | . | 1 | + | + | + | . | . | . | . | + | . | 1 | . | + | 1 | . | 1 | 1 | + | + | 2 | 1 | . | + | . | . |
| Mycelis muralis | . | . | + | + | + | + | . | . | . | . | . | + | + | . | + | + | . | . | + | r | + | + | . | . | + | + | + | 1 |
| Rubus ulmifolius | 1 | + | 1 | 1 | 1 | 2 | 1 | . | . | . | + | . | . | 2 | . | . | . | 2 | . | + | + | . | . | + | 1 | . | . | . |
| Dioscorea communis | + | + | + | + | + | . | . | . | + | + | 2 | . | + | . | . | . | . | + | . | 2 | . | . | + | . | . | . | + | . |
| Polystichum setiferum | 2 | 1 | 1 | . | + | + | + | 1 | 1 | 1 | 2 | . | . | . | . | . | . | . | . | . | . | 2 | . | . | + | . | + | . |
| Crataegus monogyna | . | . | 1 | 1 | . | . | . | . | . | . | + | + | + | 2 | 1 | . | + | + | + | . | + | . | + | . | + | . | . | . |
| Geranium robertianum | . | . | . | + | . | 1 | . | . | . | . | . | 1 | + | . | . | . | . | 1 | r | + | . | 1 | . | . | 2 | + | + | + |
| Primula vulgaris | . | . | . | . | . | . | . | . | + | . | . | . | . | + | + | 1 | + | . | . | + | + | . | + | 1 | . | + | + | + |
| Viola reichenbachiana | . | + | 1 | . | . | . | . | . | . | . | 1 | . | + | r | . | 1 | . | . | + | . | . | . | . | 1 | . | + | + | + |
| Euphorbia dulcis | . | + | . | . | . | . | . | . | . | . | . | + | + | r | + | + | + | . | . | . | . | + | + | . | + | . | . | . |
| Ostrya carpinifolia | . | . | . | 1 | . | . | . | . | . | . | 1 | 2 | . | . | . | 1 | 1 | 1 | . | . | 2 | + | . | 1 | + | . | . | . |
| Urtica dioica | + | . | . | . | . | + | + | . | . | 1 | . | 1 | . | . | + | . | . | 2 | . | + | + | . | . | . | 1 | . | . | . |
| Symphytum tuberosum subsp. angustifolium | 1 | . | . | . | . | . | . | . | . | . | + | . | . | + | . | . | + | . | + | . | . | + | . | . | + | + | + | + |
| Campanula trachelium | . | . | . | + | . | . | + | + | . | . | . | . | + | . | . | . | . | . | . | . | . | . | + | + | . | + | + | + |
| Robinia pseudoacacia | . | . | . | . | . | . | + | . | . | . | 1 | 1 | . | . | . | . | + | . | r | 2 | . | 1 | 2 | . | . | . | 2 | . |
| Ranunculus lanuginosus | + | . | + | . | . | . | + | . | . | . | . | . | . | . | + | + | . | . | . | . | . | + | . | . | . | 1 | 1 | + |
| Eupatorium cannabinum | . | . | + | . | . | . | + | . | + | + | + | + | . | . | . | . | . | . | . | . | . | . | 1 | 1 | . | + | . | . |
| Euonymus europaeus | . | + | . | + | + | . | . | 2 | . | + | . | . | . | . | . | . | . | . | . | + | + | + | + | . | . | . | . | . |
| Dryopteris filix-mas | + | . | . | + | + | + | . | . | . | . | + | . | . | . | . | . | . | . | r | 1 | . | . | . | . | + | . | + | . |
| Lamium galeobdolon subsp. flavidum | 1 | 1 | 1 | . | 1 | . | + | + | . | . | . | . | . | + | 2 | . | . | . | + | . | . | . | . | . | . | . | . | . |
| Prunus avium | . | . | . | . | . | . | . | . | . | + | . | + | + | 1 | . | 1 | + | . | . | . | . | . | . | + | + | . | . | . |
| Emerus major | . | + | . | + | . | . | . | . | . | . | + | . | + | + | . | . | . | . | . | . | . | + | . | + | . | . | . | . |
| Galium aparine | . | . | . | . | . | . | . | . | . | + | 1 | . | . | + | + | . | . | + | . | . | . | . | . | . | + | + | . | . |
| Sanicula europaea | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | 2 | + | . | . | . | . | . | + | . | . | + | + | + |
| Acer pseudoplatanus | . | . | . | . | . | + | . | . | . | 2 | . | . | + | + | r | + | + | . | . | . | . | . | . | . | . | . | . | . |
| Solidago virgaurea | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | + | + | . | r | . | . | + | . | + | . | . | + | . |
| Viola riviniana | + | . | . | + | + | + | 1 | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Tussilago farfara | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | + | . | . | . | . | 1 | 1 | . | 2 | 1 | + |
| Pulmonaria hirta | . | . | . | . | . | . | . | . | 1 | + | + | . | . | . | . | + | . | . | . | . | . | . | + | . | + | . | . | . |
| Daphne laureola | . | . | . | . | + | . | . | . | . | . | . | + | + | . | . | . | + | . | . | r | . | . | . | . | . | . | . | + |
| Helleborus viridis subsp. bocconei | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | + | + | + | . | r | . | + | . | . | . | . | . | . | . |
| Viola alba subsp. dehnhardtii | . | . | . | . | . | . | . | . | 1 | + | . | . | . | . | 1 | 1 | 1 | . | . | r | . | . | . | . | . | . | . | . |
| Equisetum arvense | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | + | + | . | . | . | r | . | + | . | . | + | . | . |
| Arum italicum | + | . | . | . | . | . | . | . | + | . | . | . | . | 1 | + | + | . | . | . | r | . | . | . | . | . | . | . | . |
| Carex sylvatica | . | . | . | . | . | . | . | . | . | 1 | r | . | . | . | . | + | . | . | . | . | . | 1 | 1 | . | + | . | . | . |
| Carpinus betulus | . | 2 | . | . | . | . | . | . | . | . | . | . | . | 1 | . | 1 | . | . | . | 1 | . | . | . | . | . | + | + | . |
| Castanea sativa | . | . | . | . | . | 2 | . | . | . | . | + | . | . | . | . | . | . | 1 | . | r | 2 | . | . | . | 1 | . | . | . |
| Arctium lappa | + | . | . | . | . | r | . | . | . | . | . | . | + | . | + | + | . | . | . | . | . | r | . | . | . | . | . | . |
| Dactylis glomerata | . | . | . | . | . | . | . | . | . | . | + | . | . | + | . | . | . | . | . | . | . | + | . | + | . | + | . | . |
| Athyrium filix-femina | 1 | . | . | 1 | 1 | 1 | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . |
| Salvia glutinosa | . | . | . | . | + | . | . | . | . | . | + | . | . | . | . | . | . | . | . | + | . | + | . | . | 2 | . | . | . |
| Cornus mas | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 2 | . | + | 1 | . |
| Lonicera etrusca | . | + | + | . | + | . | . | . | . | . | . | . | . | . | 1 | . | + | . | . | . | . | . | . | . | . | . | . | . |
| Rubia peregrina | + | . | . | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + |
| Festuca heterophylla | . | . | . | . | . | . | . | . | . | . | . | + | . | 1 | . | . | . | . | . | . | . | + | . | . | + | . | . | . |
| Chaerophyllum hirsutum (incl. C. calabrum) | . | . | . | . | . | . | . | . | . | . | . | + | + | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . |
| Rosa canina | . | . | . | . | . | . | . | . | . | . | . | . | + | + | . | + | + | . | . | . | . | . | . | . | . | . | . | . |
| Cardamine bulbifera | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | + | . | . | . | 2 | . | + | . |
| Rosa arvensis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | + | + |
| Ruscus aculeatus | . | 1 | + | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Mentha aquatica | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | + | + | . | . | . | . |
| Ulmus minor | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . |
| Rubus hirtus | . | . | . | . | . | . | 1 | 2 | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . |
| Ficaria verna | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | + | + | . |
| Ligustrum vulgare | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | + | . | + |
| Hypericum androsaemum | + | . | . | . | . | . | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Anemonoides nemorosa | . | . | . | + | . | . | . | . | r | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Prunus spinosa subsp. spinosa | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | + | . | . | . | . | . | . | . | . | + | . | . | . |
| Galium album | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | . | + | . | . |
| Lolium giganteum | . | . | . | . | . | . | . | . | . | . | . | + | + | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Bryonia dioica | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | + | + | . | . | . | . | . | . | . |
| Moehringia trinervia | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | 1 | . | r | . | . | . | . | . | . | . | . |
| Laurus nobilis | . | 1 | + | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Potentilla micrantha | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Quercus cerris | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | 1 | 1 | . | . | . | . |
| Asplenium trichomanes subsp. quadrivalens | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | r | . | . | . | . | . | . | . |
| Pteridium aquilinum subsp. aquilinum | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Equisetum telmateia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | + | . | . | . |
| Ranunculus repens | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . |
| Poa sylvicola | . | . | . | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Scrophularia nodosa | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . |
| Brachypodium rupestre | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | 2 | . | . | . | . |
| Asplenium scolopendrium subsp. scolopendrium | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . |
| Fragaria vesca subsp. vesca | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | + | . | . | . | . | . | . |
| Geranium purpureum | . | . | . | . | . | . | . | . | . | + | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Heracleum sibiricum subsp. ternatum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | r | . | . | . | . | . | . | . |
| Impatiens parviflora | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . |
| Cardamine pratensis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | r | . | . | . | . | . | . | . | . |
| Senecio ovatus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 |
| Cardamine heptaphylla | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Saxifraga rotundifolia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . |
| Ficus carica | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . |
| Cyclamen repandum subsp. repandum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . |
| Rumex sanguineus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Stellaria media subsp. media | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Lonicera caprifolium | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Poa trivialis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . |
| Arisarum proboscideum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Mercurialis perennis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + |
| Agrostis stolonifera | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . |
| Fagus sylvatica subsp. sylvatica | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . |
| Euphorbia amygdaloides | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Poa nemoralis | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . |
| Juglans regia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . |
| Physospermum cornubiense | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Parietaria officinalis | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cyclamen hederifolium | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . |
| Cruciata glabra | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Acer opalus subsp. obtusatum | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Bromopsis ramosa | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . |
| Rumex conglomeratus | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ailanthus altissima | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . |
| Chelidonium majus | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . |
| Selaginella denticulata | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Arisarum vulgare subsp. vulgare | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cardamine chelidonia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . |
| Plantago major | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . |
| Pulicaria dysenterica | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . |
| Sedum cepaea | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Veronica urticifolia | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . |
| Adoxa moschatellina subsp. moschatellina | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciandrello, S.; Angiolini, C.; Bacchetta, G.; Cutini, M.; Dumoulin, J.; Fois, M.; Gabellini, A.; Gennai, M.; Gianguzzi, L.; Landi, M.; et al. Alnus glutinosa Riparian Woodlands of Italy and Corsica: Phytosociological Classification and Floristic Diversity. Land 2023, 12, 88. https://doi.org/10.3390/land12010088
Sciandrello S, Angiolini C, Bacchetta G, Cutini M, Dumoulin J, Fois M, Gabellini A, Gennai M, Gianguzzi L, Landi M, et al. Alnus glutinosa Riparian Woodlands of Italy and Corsica: Phytosociological Classification and Floristic Diversity. Land. 2023; 12(1):88. https://doi.org/10.3390/land12010088
Chicago/Turabian StyleSciandrello, Saverio, Claudia Angiolini, Gianluigi Bacchetta, Maurizio Cutini, Jeremy Dumoulin, Mauro Fois, Antonio Gabellini, Matilde Gennai, Lorenzo Gianguzzi, Marco Landi, and et al. 2023. "Alnus glutinosa Riparian Woodlands of Italy and Corsica: Phytosociological Classification and Floristic Diversity" Land 12, no. 1: 88. https://doi.org/10.3390/land12010088