
Phylogeny and Morphology Determine Vulnerability to Global Warming in Pristimantis Frogs

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Organisms
2.3. Trait Variation–Critical Thermal Maximum (CTmax)
2.4. Measuring Environmental Exposure
2.4.1. Habitat Type and Elevation
2.4.2. Temperature
2.5. Phylogeny
2.6. Statistical Analyses
Environment or Phylogeny Shapes Local CTmax?
3. Results
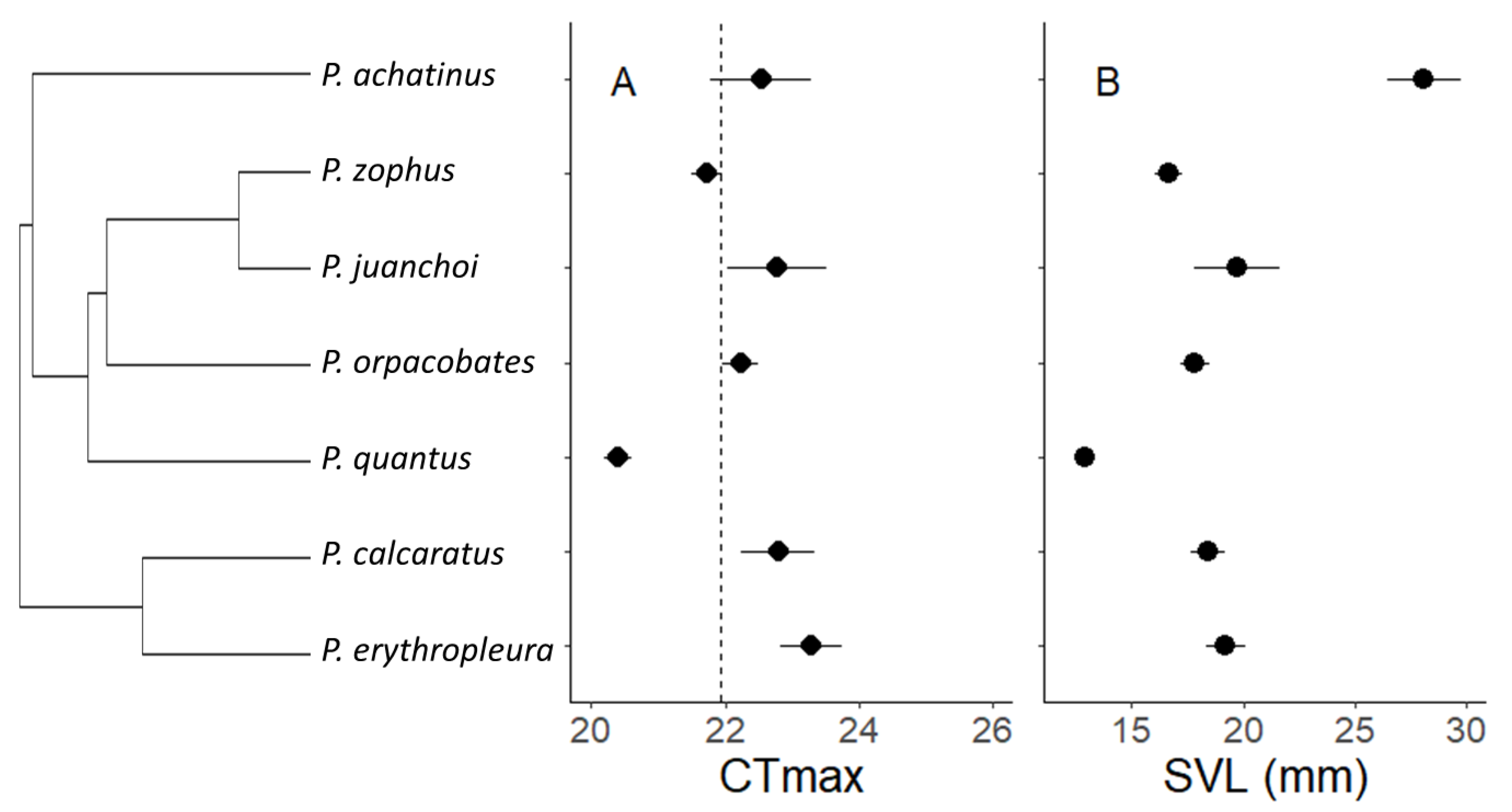
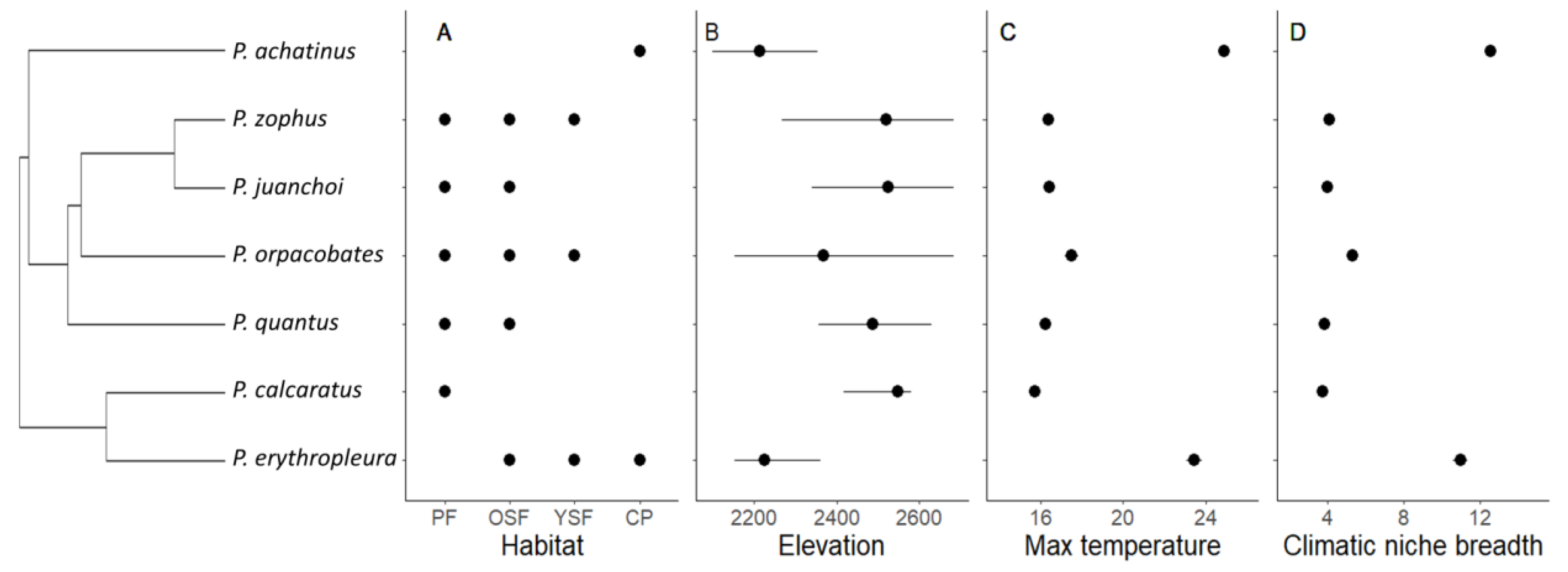
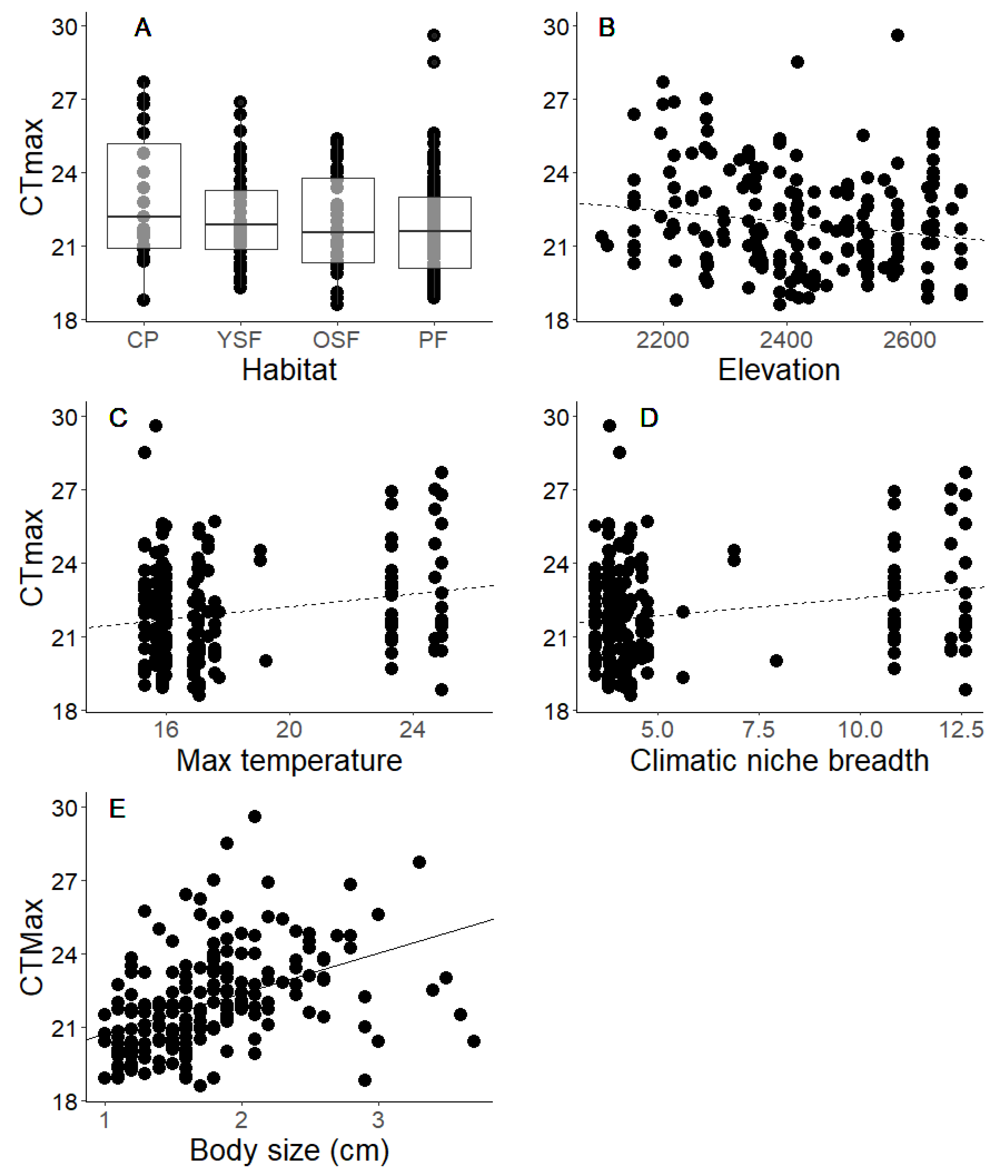
3.1. Critical Thermal Maximum (CTmax) and Environmental Exposure
3.2. Environment or Phylogeny Determine Local CTmax?
4. Discussion
4.1. Local Environmental Variables Did Not Influence CTmax Variation
4.2. Phylogeny Is Strongly Related to CTmax
4.3. Body Length Is Positively Related to CTmax
5. Conservation Implication
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheffers, B.R.; De Meester, L.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The broad footprint of climate change from genes to biomes to people. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef]
- Cahill, A.E.; Aiello-Lammens, M.E.; Fisher-Reid, M.C.; Hua, X.; Karanewsky, C.J.; Yeong Ryu, H.; Sbeglia, G.C.; Spagnolo, F.; Waldron, J.B.; Warsi, O.; et al. How does climate change cause extinction? Proc. R. Soc. B: Biol. Sci. 2013, 280, 20121890. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, B.R.; Edwards, D.P.; Diesmos, A.; Williams, S.E.; Evans, T.A. Microhabitats reduce animal's exposure to climate extremes. Glob. Change Biol. 2014, 20, 495–503. [Google Scholar] [CrossRef]
- Williams, S.E.; Shoo, L.P.; Isaac, J.L.; Hoffman, A.A.; Langham, G. Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biol. 2008, 6, e325. [Google Scholar] [CrossRef] [PubMed]
- Cowles, R.B.; Bogert, C.M. A preliminary study of the thermal requirements of desert reptiles. Bull. Am. Mus. Nat. Hist. 1944, 83, 263–296. [Google Scholar]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, J.A.M.; Jess, M.; Williams, S.E. Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [Green Version]
- González-del-Pliego, P.; Scheffers, B.R.; Freckleton, R.P.; Basham, E.W.; Araújo, M.B.; Acosta-Galvis, A.R.; Medina Uribe, C.A.; Haugaasen, T.; Edwards, D.P. Thermal tolerance and the importance of microhabitats for Andean frogs in the context of land use and climate change. J. Anim. Ecol. 2020, 89, 2451–2460. [Google Scholar] [CrossRef]
- Freckleton, R.P.; Jetz, W. Space versus phylogeny: Disentangling phylogenetic and spatial signals in comparative data. Proc. R. Soc. B: Biol. Sci. 2009, 276, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Pesquera, L.M.; Tejedo, M.; Olalla-Tárraga, M.Á.; Duarte, H.; Nicieza, A.; Solé, M. Testing the climate variability hypothesis in thermal tolerance limits of tropical and temperate tadpoles. J. Biogeogr. 2016, 43, 1166–1178. [Google Scholar] [CrossRef]
- Araujo, M.B.; Ferri-Yanez, F.; Bozinovic, F.; Marquet, P.A.; Valladares, F.; Chown, S.L. Heat freezes niche evolution. Ecol. Lett. 2013, 16, 1206–1219. [Google Scholar] [CrossRef] [Green Version]
- Llewelyn, J.; Macdonald, S.; Hatcher, A.; Moritz, C.; Phillips, B.L. Thermoregulatory behaviour explains countergradient variation in the upper thermal limit of a rainforest skink. Oikos 2016, 126, 748–757. [Google Scholar] [CrossRef]
- Phillips, B.L.; Muñoz, M.M.; Hatcher, A.; Macdonald, S.L.; Llewelyn, J.; Lucy, V.; Moritz, C. Heat hardening in a tropical lizard: Geographic variation explained by the predictability and variance in environmental temperatures. Funct. Ecol. 2015, 30, 1161–1168. [Google Scholar] [CrossRef]
- Kellermann, V.; van Heerwaarden, B.; Sgrò, C.M.; Hoffmann, A.A. Fundamental evolutionary limits in ecological traits drive Drosophila species distributions. Science 2009, 325, 1244–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, N.; Jetz, W.; Freckleton, R.P. Phylogenetic comparative approaches for studying niche conservatism. J. Evol. Biol. 2010, 23, 2529–2539. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.F. Stabilizing selection and the comparative analysis of adaptation. Evol. Int. J. Org. Evol. 1997, 51, 1341–1351. [Google Scholar] [CrossRef]
- Labra, A.; Pienaar, J.; Hansen, T.F. Evolution of thermal physiology in Liolaemus lizards: Adaptation, phylogenetic inertia, and niche tracking. Am. Nat. 2009, 174, 204–220. [Google Scholar] [CrossRef]
- Kellermann, V.; Loeschcke, V.; Hoffmann, A.A.; Kristensen, T.N.; Fløjgaard, C.; David, J.R.; Svenning, J.-C.; Overgaard, J. Phylogenetic constraints in key functional traits behind species’ climate niches: Patterns of desiccation and cold resistance across 95 Drosophila species. Evol. Int. J. Org. Evol. 2012, 66, 3377–3389. [Google Scholar] [CrossRef]
- González-Del-Pliego, P.; Freckleton, R.P.; Edwards, D.P.; Koo, M.S.; Scheffers, B.R.; Pyron, R.A.; Jetz, W. Phylogenetic and Trait-Based Prediction of Extinction Risk for Data-Deficient Amphibians. Curr. Biol. CB 2019, 29, 1557–1563.e1553. [Google Scholar] [CrossRef]
- González del Pliego, P.; Scheffers, B.R.; Basham, E.W.; Woodcock, P.; Wheeler, C.; Gilroy, J.J.; Medina Uribe, C.A.; Haugaasen, T.; Freckleton, R.P.; Edwards, D.P. Thermally buffered microhabitats recovery in tropical secondary forests following land abandonment. Biol. Conserv. 2016, 201, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Hurtt, G.C.; Chini, L.P.; Frolking, S.; Betts, R.A.; Feddema, J.; Fischer, G.; Fisk, J.P.; Hibbard, K.; Houghton, R.A.; Janetos, A.; et al. Harmonization of land-use scenarios for the period 1500–2100: 600 years of global gridded annual land-use transitions, wood harvest, and resulting secondary lands. Clim. Change 2011, 109, 117. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, S.; van Heerwaarden, B.; Kellermann, V.; Sgrò, C.M. Evolutionary capacity of upper thermal limits: Beyond single trait assessments. J. Exp. Biol. 2014, 217, 1918–1924. [Google Scholar] [CrossRef]
- Cooper, N.; Freckleton, R.P.; Jetz, W. Phylogenetic conservatism of environmental niches in mammals. Proc. R. Soc. B: Biol. Sci. 2011, 278, 2384–2391. [Google Scholar] [CrossRef]
- Menke, S.B.; Holway, D.A.; Fisher, R.N.; Jetz, W. Characterizing and predicting species distributions across environments and scales: Argentine ant occurrences in the eye of the beholder. Glob. Ecol. Biogeogr. 2009, 18, 50–63. [Google Scholar] [CrossRef]
- Llewelyn, J.; Macdonald, S.L.; Hatcher, A.; Moritz, C.; Phillips, B.L. Intraspecific variation in climate-relevant traits in a tropical rainforest lizard. Divers. Distrib. 2016, 22, 1000–1012. [Google Scholar] [CrossRef]
- Ribeiro, P.L.; Camacho, A.; Navas, C.A. Considerations for assessing maximum critical temperatures in small ectothermic animals: Insights from leaf-cutting ants. PLoS ONE 2012, 7, e32083. [Google Scholar] [CrossRef] [Green Version]
- Basham, E.W.; González del Pliego, P.; Acosta-Galvis, A.R.; Woodcock, P.; Medina Uribe, C.A.; Haugaasen, T.; Gilroy, J.J.; Edwards, D.P. Quantifying carbon and amphibian co-benefits from secondary forest regeneration in the Tropical Andes. Anim. Conserv. 2016, 19, 548–560. [Google Scholar] [CrossRef]
- Navas, C.A.; Antoniazzi, M.M.; Carvalho, J.E.; Suzuki, H.; Jared, C. Physiological basis for diurnal activity in dispersing juvenile Bufo granulosus in the Caatinga, a Brazilian semi-arid environment. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 647–657. [Google Scholar] [CrossRef]
- Catenazzi, A.; Lehr, E.; Vredenburg, V.T. Thermal physiology, disease, and amphibian declines on the eastern slopes of the Andes. Conserv. Biol. J. Soc. Conserv. Biol. 2014, 28, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Robledo, C.; Kuprewicz, E.K.; Staines, C.L.; Erwin, T.L.; Kress, W.J. Limited tolerance by insects to high temperatures across tropical elevational gradients and the implications of global warming for extinction. Proc. Natl. Acad. Sci. USA 2016, 113, 680–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowakowski, A.J.; Watling, J.I.; Whitfield, S.M.; Todd, B.D.; Kurz, D.J.; Donnelly, M.A. Tropical amphibians in shifting thermal landscapes under land use and climate change. Conserv. Biol. 2017, 31, 96–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffers, B.R.; Brunner, R.M.; Ramirez, S.D.; Shoo, L.P.; Diesmos, A.; Williams, S.E. Thermal buffering of microhabitats is a critical factor mediating warming vulnerability of frogs in the Philippine biodiversity hotspot. Biotropica 2013, 45, 628–635. [Google Scholar] [CrossRef]
- von May, R.; Catenazzi, A.; Corl, A.; Santa-Cruz, R.; Carnaval, A.C.; Moritz, C. Divergence of thermal physiological traits in terrestrial breeding frogs along a tropical elevational gradient. Ecol. Evol. 2017, 7, 3257–3267. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Pyron, R.A. The interplay of past diversification and evolutionary isolation with present imperilment across the amphibian tree of life. Nat. Ecol. Evol. 2018, 2, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, S.P.; Garland, T. Tempo and mode in evolution: Phylogenetic inertia, adaptation and comparative methods. J. Evol. Biol. 2002, 15, 899–910. [Google Scholar] [CrossRef]
- Therneau, T.M. Coxme: Mixed Effects Cox Models. R Package Version 2.2-5. Available online: https://cran.r-project.org/web/packages/coxme/coxme.pdf (accessed on 24 November 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 24 November 2022).
- Freckleton, R.P.; Rees, M. Comparative analysis of experimental data. Methods Ecol. Evol. 2019, 10, 1308–1321. [Google Scholar] [CrossRef] [Green Version]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Muñoz, M.M.; Langham, G.M.; Brandley, M.C.; Rosauer, D.F.; Williams, S.E.; Moritz, C. Basking behavior predicts the evolution of heat tolerance in Australian rainforest lizards. Evol. Int. J. Org. Evol. 2016, 70, 2537–2549. [Google Scholar] [CrossRef]
- Nowakowski, A.J.; Watling, J.I.; Thompson, M.E.; Brusch Iv, G.A.; Catenazzi, A.; Whitfield, S.M.; Kurz, D.J.; Suárez-Mayorga, Á.; Aponte-Gutiérrez, A.; Donnelly, M.A.; et al. Thermal biology mediates responses of amphibians and reptiles to habitat modification. Ecol. Lett. 2018, 21, 345–355. [Google Scholar] [CrossRef]
- van Heerwaarden, B.; Kellermann, V.; Sgrò, C.M. Limited scope for plasticity to increase upper thermal limits. Funct. Ecol. 2016, 21, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: New York, NY, USA, 2009; pp. 1–290. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Huey, R.B.; Deutsch, C.A. Putting the heat on tropical animals. Science 2008, 320, 1296–1297. [Google Scholar] [CrossRef]
- Hof, C.; Rahbek, C.; Araújo, M.B. Phylogenetic signals in the climatic niches of the world's amphibians. Ecography 2010, 33, 242–250. [Google Scholar] [CrossRef]
- Pounds, J.A.; Fogden, M.P.L.; Campbell, J.H. Biological response to climate change on a tropical mountain. Nature 1999, 398, 611–615. [Google Scholar] [CrossRef]
- Grigg, J.W.; Buckley, L.B. Conservatism of lizard thermal tolerances and body temperatures across evolutionary history and geography. Biol. Lett. 2013, 9, 20121056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von May, R.; Catenazzi, A.; Santa-Cruz, R.; Gutierrez, A.S.; Moritz, C.; Rabosky, D.L. Thermal physiological traits in tropical lowland amphibians: Vulnerability to climate warming and cooling. PLoS ONE 2019, 14, e02197592019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Donner, S.D.; Skirving, W.J.; Little, C.M.; Oppenheimer, M.; Hoegh-Guldberg, O.V.E. Global assessment of coral bleaching and required rates of adaptation under climate change. Glob. Change Biol. 2005, 11, 2251–2265. [Google Scholar] [CrossRef]
- Kozak, K.H.; Wiens, J.J. Niche conservatism drives elevational diversity patterns in Appalachian salamanders. Am. Nat. 2010, 176, 40–54. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar] [CrossRef] [Green Version]
- Hone, D.W.E.; Benton, M.J. The evolution of large size: How does Cope’s rule work? Trends Ecol. Evol. 2005, 20, 4–6. [Google Scholar] [CrossRef]
- Huey, R.B.; Hertz, P.E.; Sinervo, B. Behavioral drive versus behavioral inertia in evolution: A null model approach. Am. Nat. 2003, 161, 357–366. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [Green Version]
- González-del-Pliego, P.; Scheffers, B.R.; Freckleton, R.P.; Basham, E.W.; Araújo, M.B.; Acosta-Galvis, A.R.; Medina Uribe, C.A.; Haugaasen, T.; Edwards, D.P. Data from: Thermal tolerance and the importance of microhabitats for Andean frogs in the context of land-use and climate change. Dryad Digit. Repos. 2020. [Google Scholar] [CrossRef]
- González del Pliego, P. Amphibian Conservation in the Face of Land-Use Change and Global Warming; University of Sheffield: Sheffield, UK, 2017. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Value (se) | z | p | ||
|---|---|---|---|---|
| Fixed | Cattle pasture (Intercept) | 18.83 (6.31) | 2.98 | <0.001 |
| Young secondary forest | 0.02 (0.77) | 0.03 | 0.97 | |
| Old secondary forest | 0.32 (1.05) | 0.31 | 0.76 | |
| Primary forest | 0.47 (1.14) | 0.42 | 0.68 | |
| Elevation | −0.0004 (0.001) | −0.30 | 0.77 | |
| Max temperature | 0.09 (0.34) | −0.14 | 0.89 | |
| Climatic niche breadth | −0.04 (0.34) | −0.14 | 0.89 | |
| SVL (body size) | 1.49 (0.28) | 5.32 | <0.001 | |
| Random | Std Dev | Variance | ||
| Phylogenetic signal (V) | 0.63 | 0.40 | ||
| Variation in species means (S) | 0.04 | 0.002 | ||
| Error variance (I) | 0.04 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-del-Pliego, P.; Freckleton, R.P.; Scheffers, B.R.; Basham, E.W.; Acosta-Galvis, A.R.; Medina Uribe, C.A.; Haugaasen, T.; Edwards, D.P. Phylogeny and Morphology Determine Vulnerability to Global Warming in Pristimantis Frogs. Land 2023, 12, 130. https://doi.org/10.3390/land12010130
González-del-Pliego P, Freckleton RP, Scheffers BR, Basham EW, Acosta-Galvis AR, Medina Uribe CA, Haugaasen T, Edwards DP. Phylogeny and Morphology Determine Vulnerability to Global Warming in Pristimantis Frogs. Land. 2023; 12(1):130. https://doi.org/10.3390/land12010130
Chicago/Turabian StyleGonzález-del-Pliego, Pamela, Robert P. Freckleton, Brett R. Scheffers, Edmund W. Basham, Andrés R. Acosta-Galvis, Claudia A. Medina Uribe, Torbjørn Haugaasen, and David P. Edwards. 2023. "Phylogeny and Morphology Determine Vulnerability to Global Warming in Pristimantis Frogs" Land 12, no. 1: 130. https://doi.org/10.3390/land12010130