
Exploring Functional Diversity and Community Structure of Diazotrophic Endophytic Bacteria Associated with Pennisetum glaucum Growing under Field in a Semi-Arid Region

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Variety and Sampling Site
2.2. Culture Media
2.3. Isolation of Endophytic Bacteria
2.4. Enterobacterial Repetitive Intergenic Consensus Sequences-PCR (ERIC-PCR)
2.5. Amplification and Analysis of 16S Ribosomal RNA Gene
2.6. 16S Ribosomal RNA Gene Sequencing
2.7. Diversity Index and Relative Species Abundance (RSA)
2.8. Estimation of Plant Growth-Promoting Properties
2.8.1. Acetylene Reduction Assay
2.8.2. Amplification of nifH Gene
2.8.3. Phosphate Solubilization, IAA, and Siderophore Production
2.8.4. Test of Cellulolytic and Pectinolytic Activity
2.9. Evaluation of Cross-Infection Ability of Bacterial Isolates and Their Effects on Plant Growth
2.10. Plant and Dominant Bacterial Strain Used
2.11. Inoculation of Wheat Plant with Bacterial Strains
2.12. RNA Extraction and cDNA Preparation
2.13. Quantitative-PCR of Defense-Related Genes
2.14. The qPCR Based Quantification of Bacterial Colonization: Colonization Studies
2.15. Statistical Analysis
3. Results
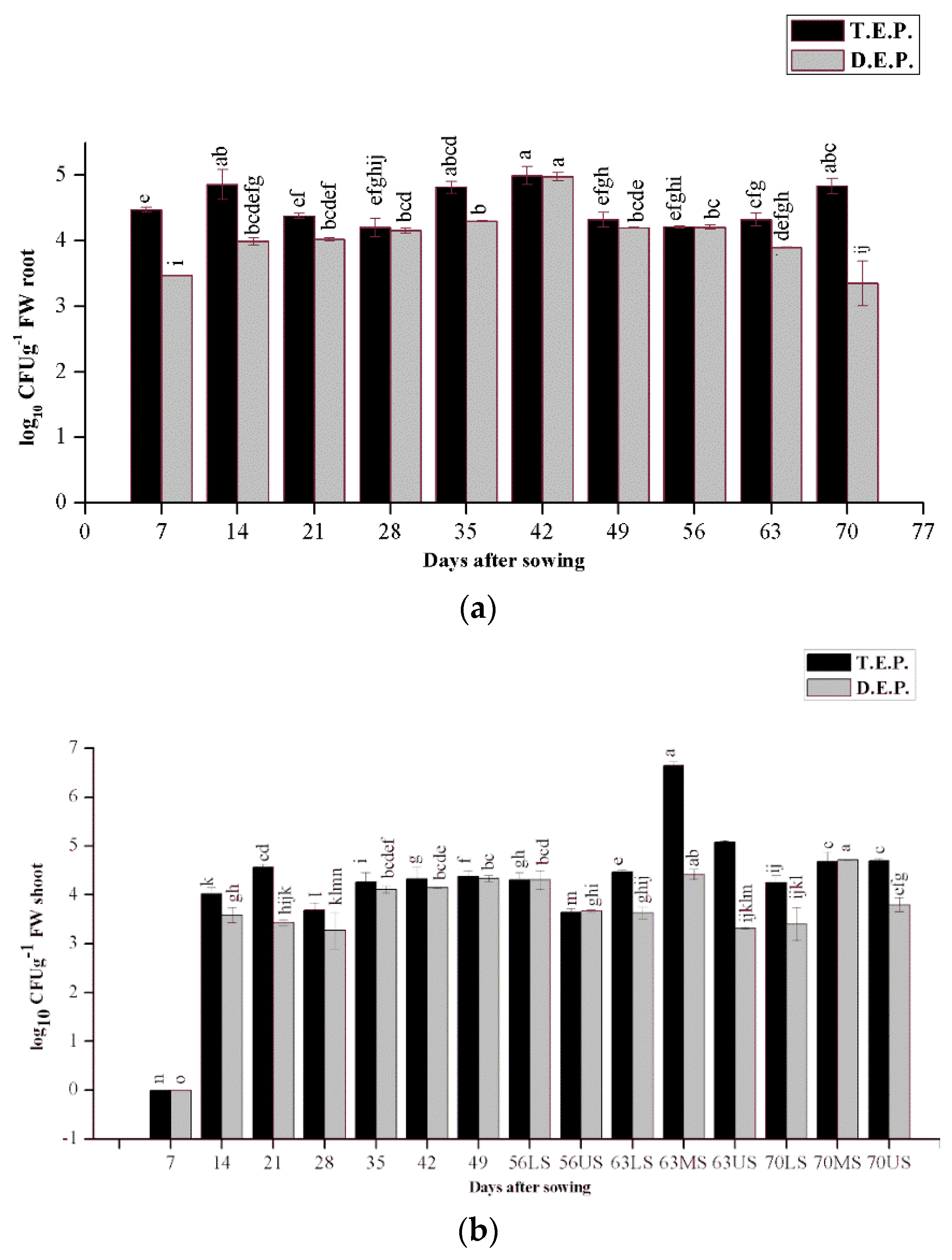
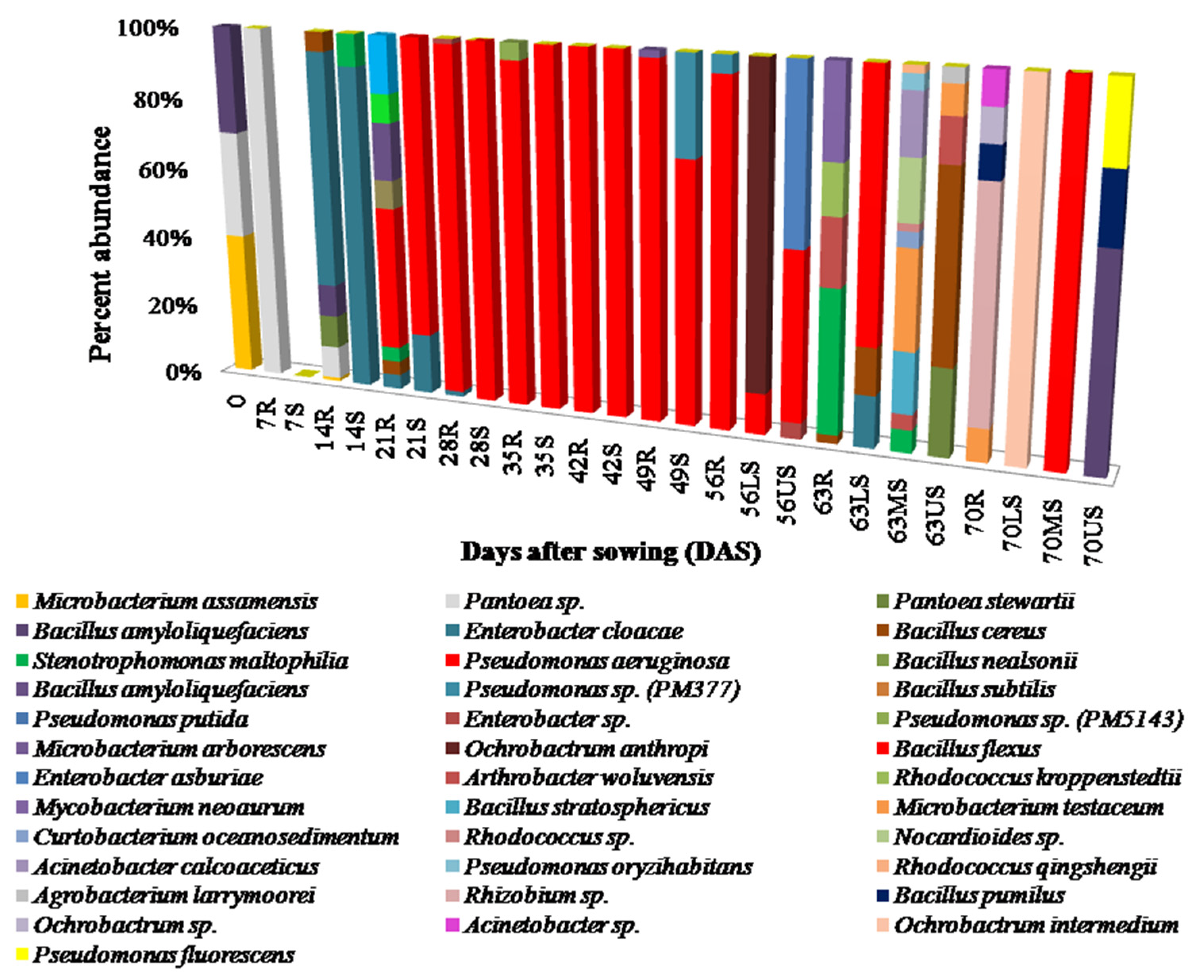
3.1. Isolation of Endophytic Bacteria and Study of Population Dynamics
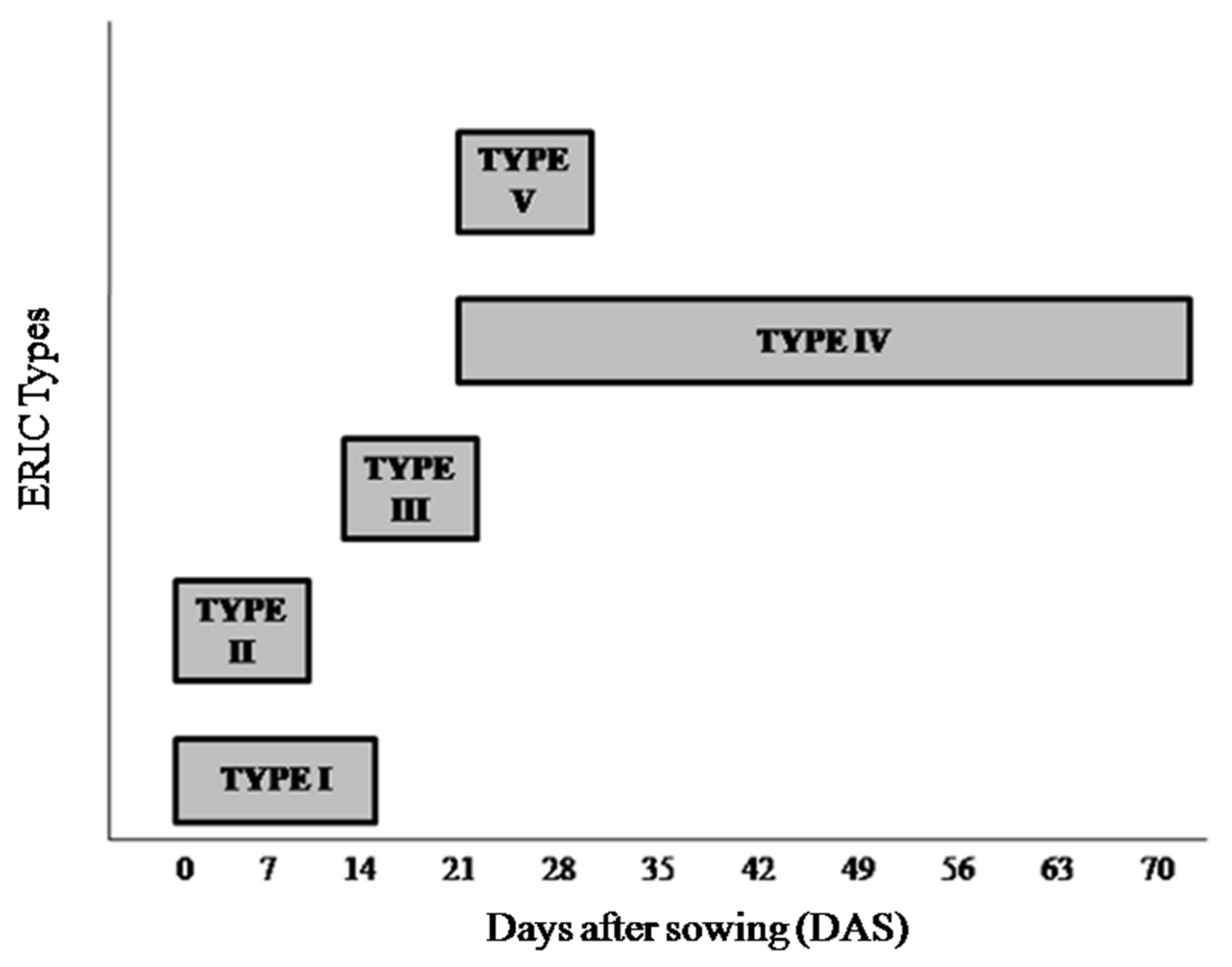
3.2. Study of Molecular Diversity Using ERIC-PCR as a Biomarker
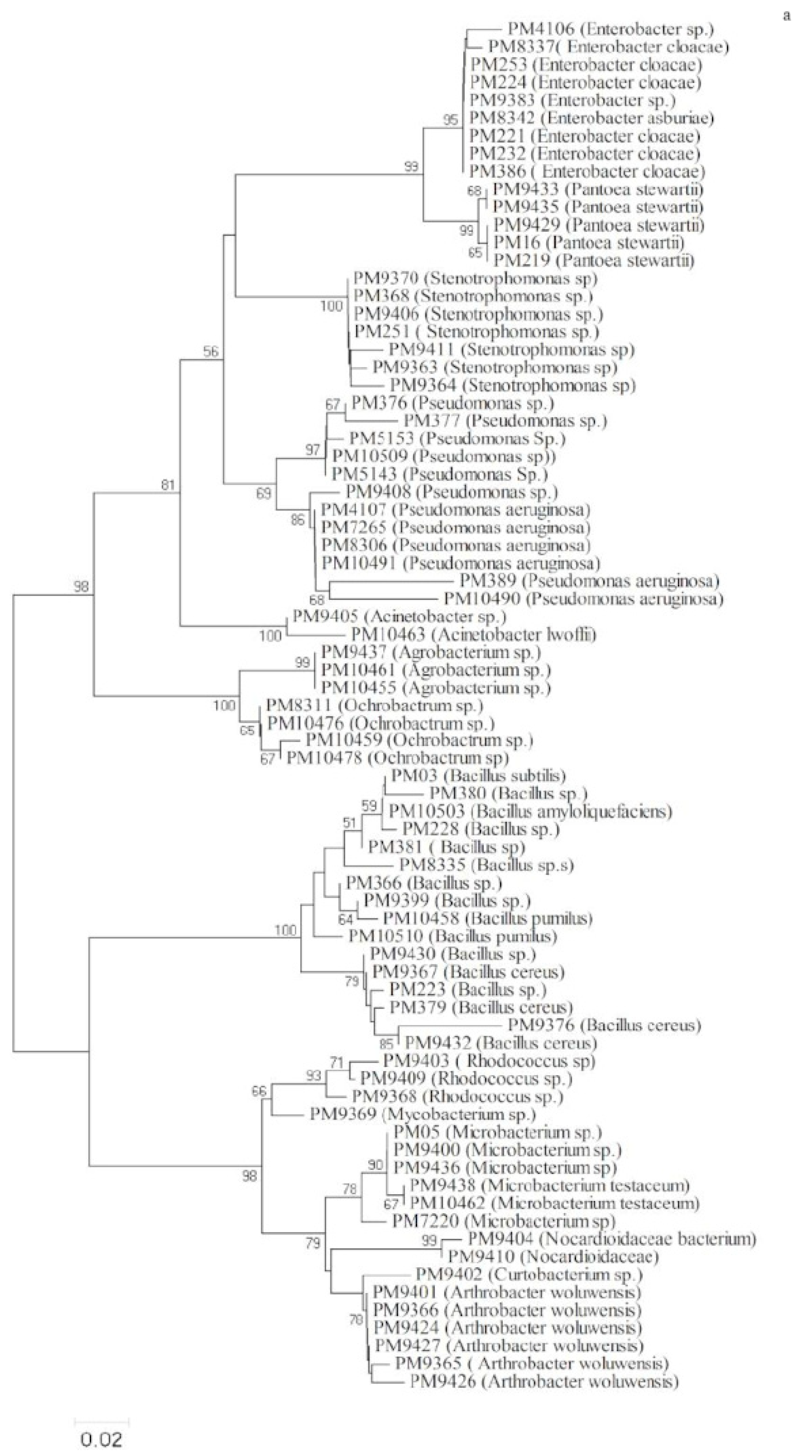
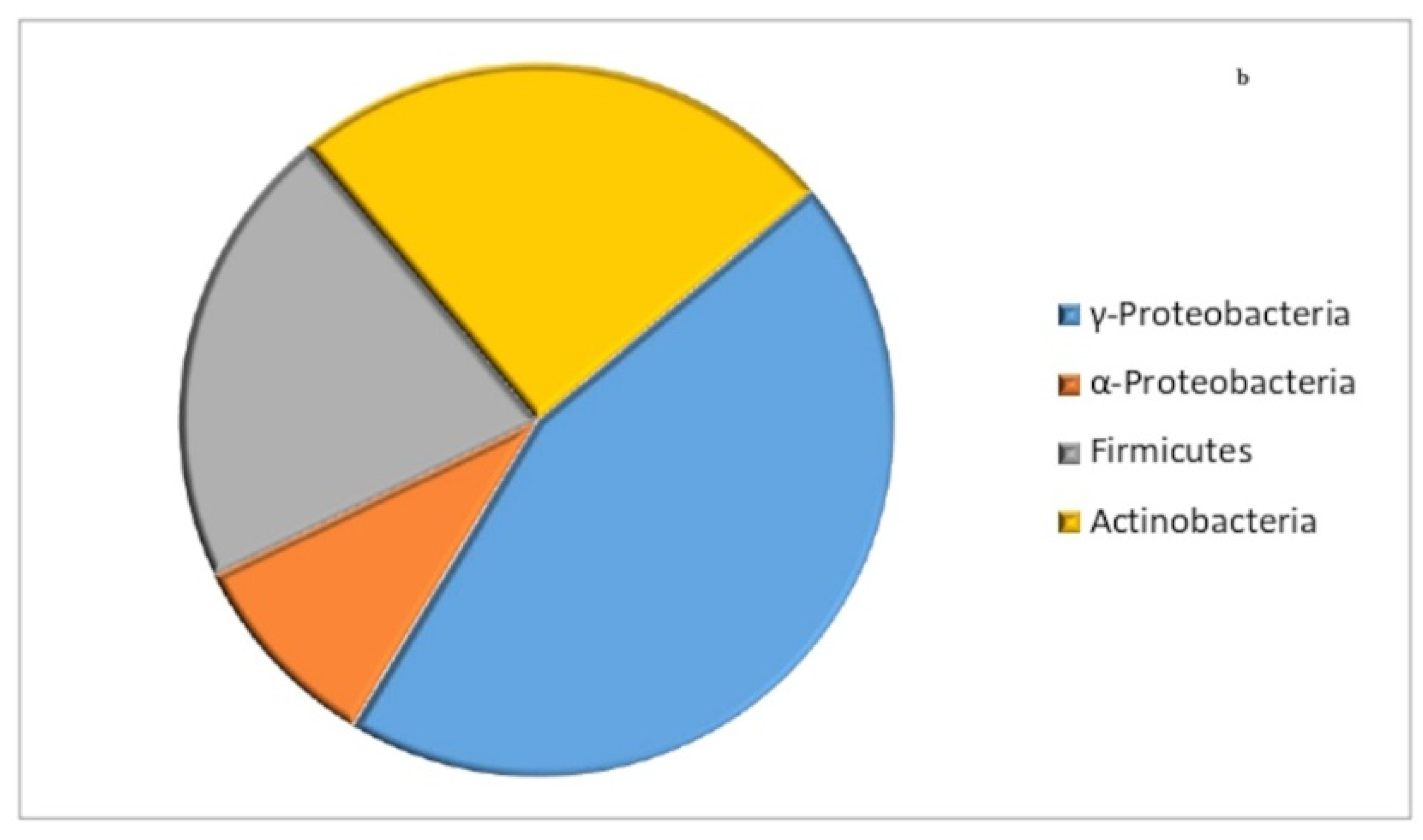
3.3. Phylogenetic Analysis Based on 16S rRNA Gene Sequence Analysis
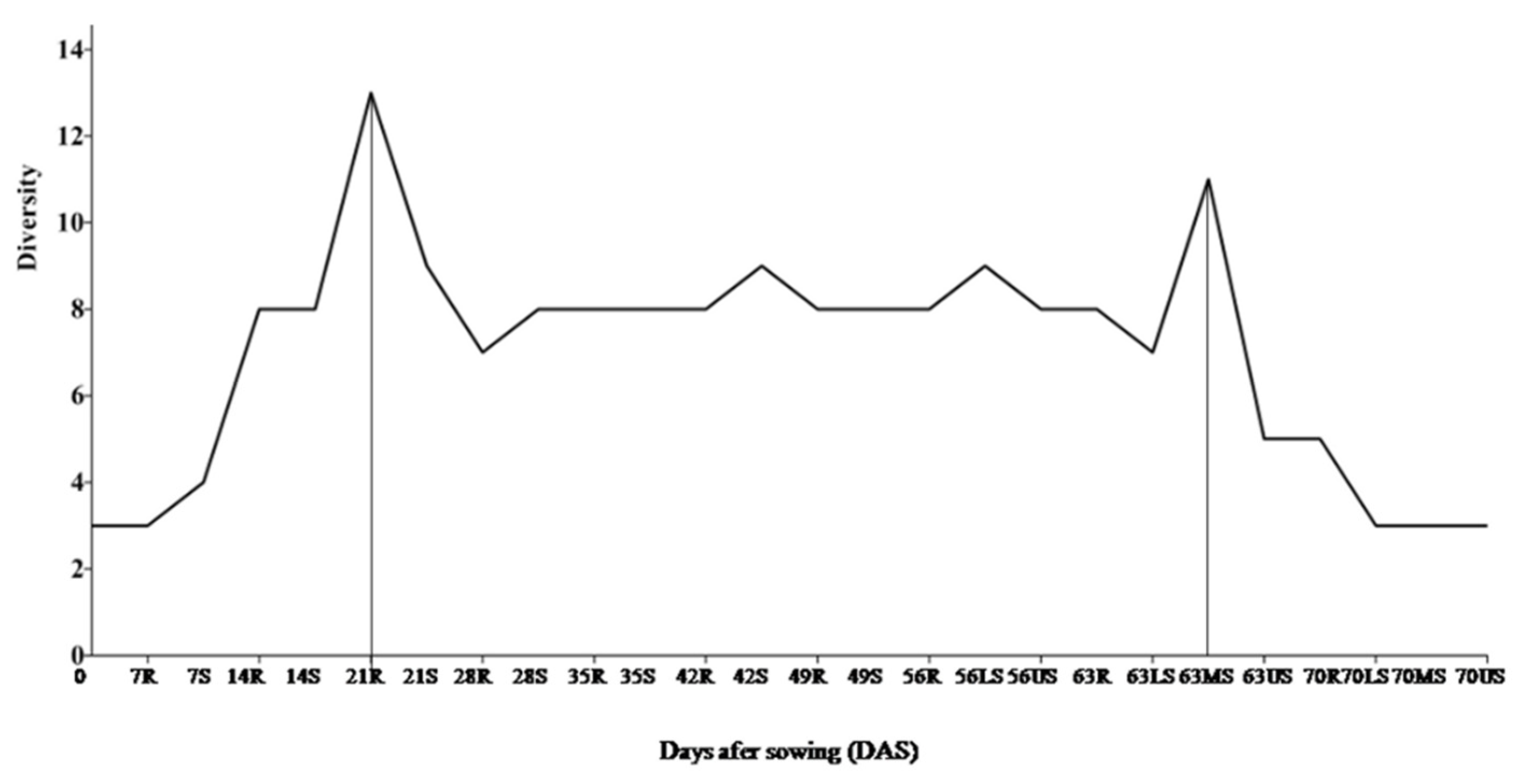
3.4. Periodic Molecular Diversity and Dynamics of Diazotrophic Bacteria
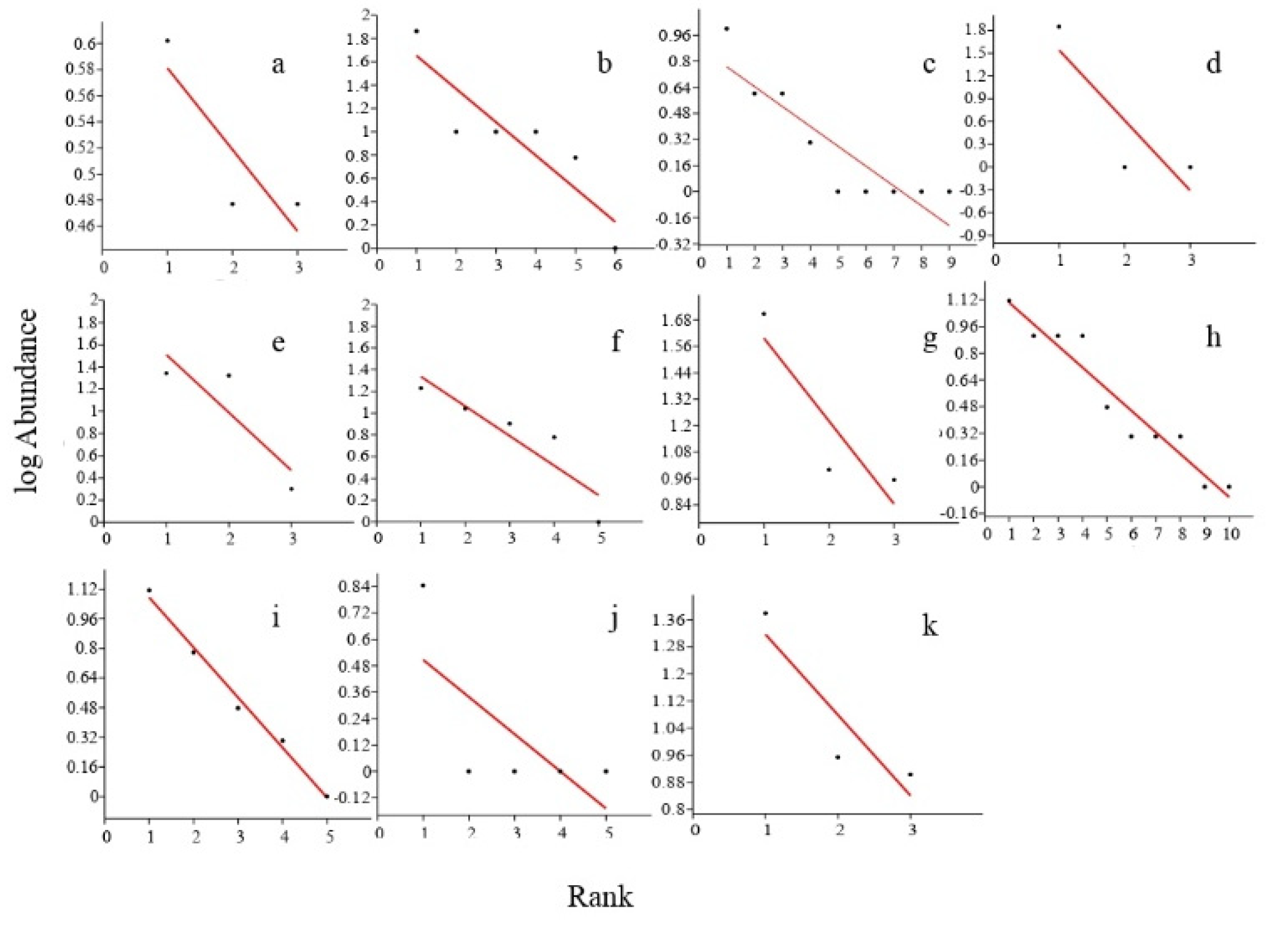
3.5. Relative Species Abundance
3.6. Test of Plant Growth Promotion Activities
3.6.1. Confirmation of Diazotrophy
3.6.2. Phosphate Solubilization, IAA and Siderophore Production
3.6.3. Tests for Cellulolytic and Pectinolytic Activities
3.7. In Vitro Studies of Plant Growth
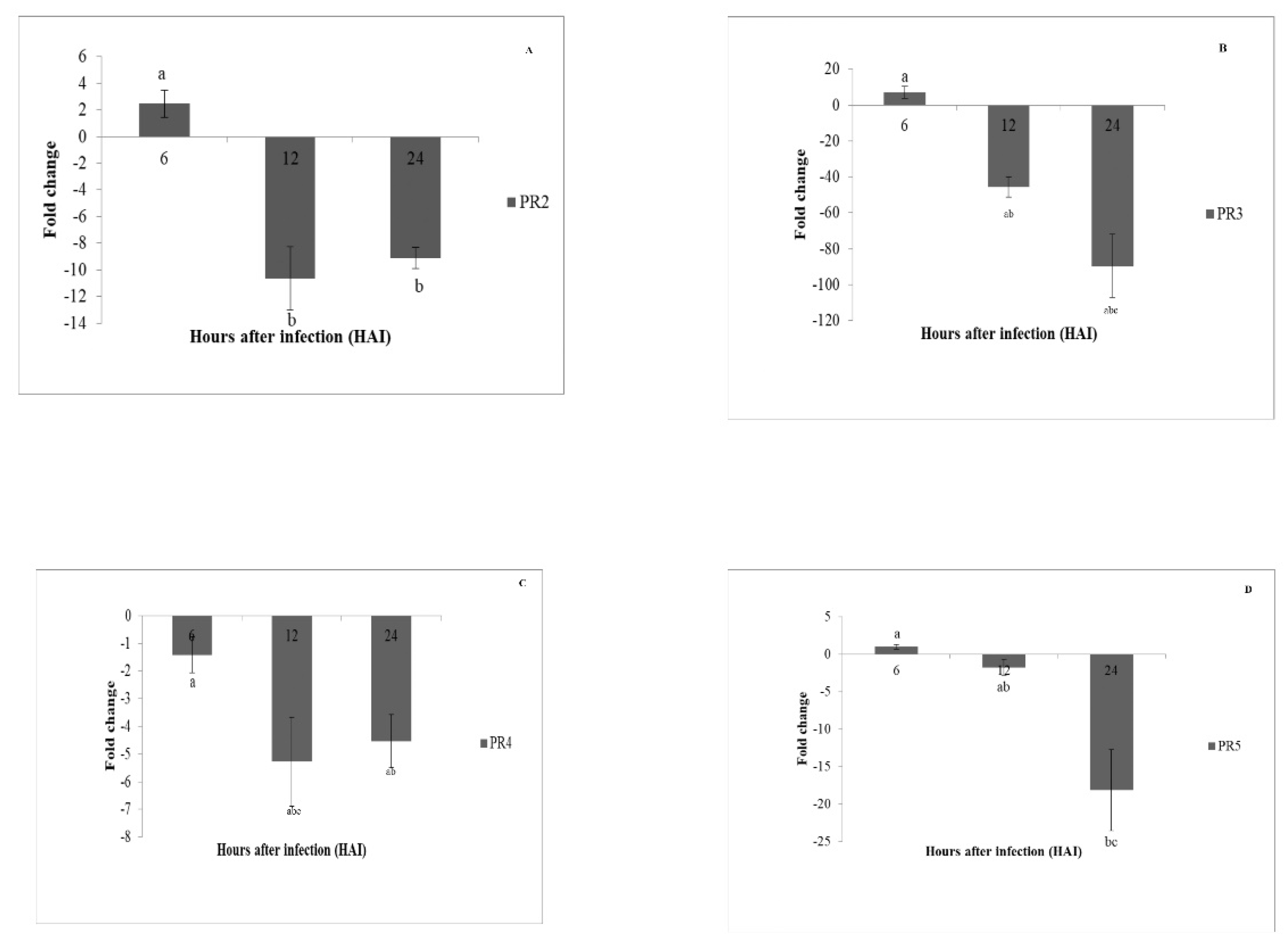
3.8. Gene Expression Analysis of Pathogenesis Related Genes
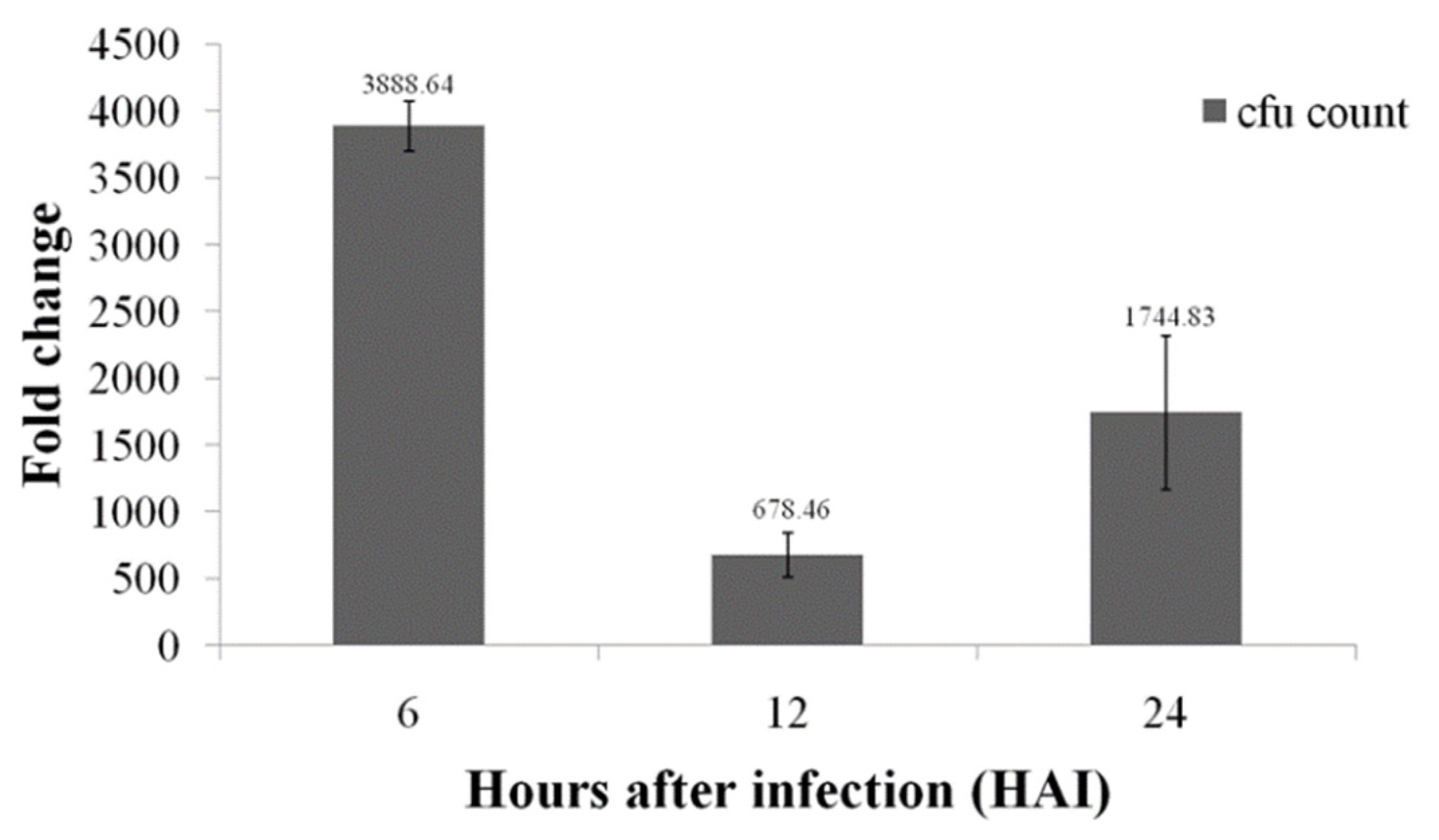
3.9. Colonization Studies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhattacharjee, R.; Singh, A.; Mukhopadhyay, S.N. Use of nitrogen-fixing bacteria as biofertilizer for non-legumes: Prospects and challenges. Appl. Microbiol. Biotechnol. 2008, 80, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Panwar, J.; Akhtar, M.; Jha, P. Endophytic nitrogen-fixing bacteria as biofertilizer. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 183–221. [Google Scholar]
- Afzala, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Sankar, A.; Devika, O.S.; Singh, S.; Parihar, M.; Rakshit, A.; Sayyed, R.Z.; Gafur, A.; Ansari, M.J.; Danish, S.; et al. Optimizing nutrient use efficiency, productivity, energetics, and economics of red cabbage following mineral fertilization and biopriming with compatible rhizosphere microbes. Sci. Rep. 2021, 11, 15680. [Google Scholar] [CrossRef]
- Lodewyckx, C.; Vangronsveld, J.; Porteous, F.; Moore, E.R.B.; Taghavi, S.; Mezgeay, M.; Van Der Lelie, D. Endophytic bacteria and their potential applications. Crit. Rev. Plant Sci. 2002, 21, 583–606. [Google Scholar] [CrossRef]
- Hameeda, B.; Rupela, O.P.; Reddy, G.; Satyavani, K. Application of plant growth-promoting bacteria associated with composts and macrofauna for growth promotion of pearl millet (Pennisetum glaucum L.). Biol. Fertil. Soils 2006, 43, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Pallavi, S.P.; Srinivas, P.; Kumar, V.P.; Chandra, S.J.; Reddy, S.R. Plant growth promoting activities of fluorescent pseudomonads associated with some crop plants. Afr. J. Microbiol. Res. 2010, 4, 1491–1494. [Google Scholar]
- Manga, V.K.; Yadav, O.P. Effect of seed size on developmental traits and ability to tolerate drought in pearl millet. J. Arid Environ. 1995, 29, 169–172. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Van Loon, L.; Van Strien, E. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Jacobs, S.; Zechmann, B.; Molitor, A.; Trujillo, M.; Petutschnig, E.; Lipka, V.; Kogel, K.-H.; Schäfer, P. Broad-spectrum suppression of innate immunity is required for colonization of Arabidopsis roots by the fungus Piriformospora indica. Plant Physiol. 2011, 156, 726–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamioudis, C.; Pieterse, C.M. Modulation of host immunity by beneficial microbes. Mol. Plant-Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.; Panwar, J.; Jha, P.N. Natural occurrence of P. aeruginosa, a dominant cultivable diazotrophic endophytic bacterium colonizing Pennisetum glaucum (L.) R. Br. Appl. Soil Ecol. 2013, 64, 252–261. [Google Scholar]
- Gupta, G.; Gupta, A.D.; Jha, P.N. Biocontrolling and colonization potential of P. aeruginosa PM389 Isolated from Pennisetum glaucum under field conditions. Int. J. Res. Emerg. Sci. Technol. 2015, 2, 104–116. [Google Scholar]
- Ghosh, S.P. Agro-Climatic Zone Specific Research-Indian Perspective under NARP (ICAR); Indian Council of Agriculture Research, India: New Delhi, India, 1991. [Google Scholar]
- Shyampura, R.L.; Singh, S.K.; Singh, R.S.; Jain, B.L.; Gajbhiye, K.S. Soil Series of Rajasthan; Udaipur: Technical Bulletin, NBSS & LUP; Indian Council of Agriculture Research, India: New Delhi, India, 2002. [Google Scholar]
- Cavalcante, V.A.; Döbereiner, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugarcane. Plant Soil 1988, 108, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Misra, N.; Gupta, G.; Jha, P.N. Assessment of mineral phosphate solubilizing properties and molecular characterization of zinc tolerant bacteria. J. Basic Microbiol. 2012, 52, 549–558. [Google Scholar] [CrossRef]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 2004, 19, 6823–6831. [Google Scholar] [CrossRef]
- NCBI Database. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 2 December 2013).
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Mahaffee, W.F.; Kloepper, J.W. Temporal changes in the bacterial communities of soil, rhizosphere, and endorhiza associated with field-grown Cucumber (Cucumis sativus L.). Microb. Ecol. 1997, 34, 210–223. [Google Scholar] [CrossRef]
- Odum, E.P.; Barrett, J.W. Fundamentals of Ecology; Brooks/Cole-Thomson Learning; Thomson Brooks/Cole: Belmont, CA, USA, 2005. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Mehta, S.; Nautiyal, C.S. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr. Microbiol. 2001, 43, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N. Assay of inorganic phosphate, total phosphate and phosphatases. In Methods in Enzymology; Ginsburg, N.V., Ed.; Academic Press Publisher: New York, NY, USA, 1996; Volume 8, pp. 115–118. [Google Scholar]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indole-acetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasana, R.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A rapid and easy method for the detection of microbial cellulases on agar plates using Gram’s Iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Reinhold-Hurek, B.; Hurek, T.; Gillis, M.; Hoste, B.; Vancanneyt, M.; Kersters, K.; De Ley, J. Azoarcus gen. nov., nitrogen-fixing proteobacteria associated with roots of kallar grass (Leptochloa fusca (L.) Kunth), and description of two species, Azoarcus indigens sp. nov. and Azoarcus communis sp. nov. Int. J. Syst. Bacteriol. 1993, 43, 574–584. [Google Scholar] [CrossRef]
- Jha, P.; Kumar, A. Characterization of novel plant growth promoting endophytic bacterium Achromobacter xylosoxidans from wheat plant. Microb. Ecol. 2009, 58, 179–188. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Zahir, Z.A. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 2004, 96, 473–480. [Google Scholar] [CrossRef]
- Garima, G.; Das, A.; Jha, P.; Jha, P.N. Endophytic Bacteria Pseudomonas aeruginosa PM389 Subsists Host’s (Triticum aestivum) Immune Response for Gaining Entry Inside the Host. J. Pure Appl. Microbiol. 2021, 15, 2486–2497. [Google Scholar]
- Gupta, G.; Das Gupta, A.; Jha, P.N. Comparative analysis of defence responses Triticum aestivum L in response to the endophytic Pseudomonas aeruginosa PM389 and the non-host microbial pathogens Erwinia carotovora and Fusarium monaliforme. Res. J Biotech. 2022, 17, 107–117. [Google Scholar] [CrossRef]
- SpeciesPrimer: A bioinformatics pipeline dedicated to the design of qPCR primers for the quantification of bacterial species. PeerJ. 2020, 8, e8544. [CrossRef] [Green Version]
- Kloepper, J.W.; Beauchamp, C.J. A review of issues related to measuring colonization of plant roots by bacteria. Can. J. Microbiol. 1992, 38, 1219–1232. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists, 1st ed.; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Zafar-ul-Hye, M.; Akbar, M.N.; Iftikhar, Y.; Abbas, M.; Zahid, A.; Fahad, S.; Datta, R.; Ali, M.; Elgorban, A.M.; Ansari, M.J.; et al. Rhizobacteria inoculation and caffeic acid alleviated drought stress in lentil plants. Sustainability 2021, 13, 9603. [Google Scholar] [CrossRef]
- Andreote, F.D.; Rocha, U.N.; Araújo, W.L.; Azevedo, J.L.; Van Overbeek, L.S. Effect of bacterial inoculation, plant genotype and developmental stage on root-associated and endophytic bacterial communities in potato (Solanum tuberosum). Antonie Leeuwenhoek 2010, 97, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; Postma, J.; Van Veen, J.A.; Van Elsas, J.D. Effect of above-ground wplant species on soil microbial community structure and its impact on suppression of Rhizoctonia solani AG3. Environ Microbiol. 2006, 8, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Kaplan, H.; Sessitsch, A.; Nowak, J.; Ait Barka, E.; Clément, C. Endophytic colonization of Vitis vinifera L. by Burkholderia phytofirmans strain PsJN: From the rhizosphere to inflorescence tissues. FEMS Microbiol. Ecol. 2008, 63, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. CRC Crit. Rev. Plant Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Ferreira, A.; Quecine, M.C.; Lacava, P.T.; Oda, S.; Azevedo, J.L.; Araújo, W.L. Diversity of endophytic bacteria from Eucalyptus species seeds and colonization of seedlings by Pantoea agglomerans. FEMS Microbiol. Lett. 2008, 287, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilley, A.K.; Fry, J.C.; Bailey, M.J.; Day, M.J. Comparison of aerobic heterotrophic taxa isolated from four root domains of mature sugar beet (Beta vulgaris). FEMS Microbiol. Ecol. 1996, 21, 231–242. [Google Scholar] [CrossRef]
- Germida, J.; Siciliano, S.D.; Renato De Freitas, J.; Seib, A.M. Diversity of root-associated bacteria associated with field-grown canola (Brassica napus L.) and wheat (Triticum aestivum L.). FEMS Microbiol. Ecol. 1998, 26, 43–50. [Google Scholar] [CrossRef]
- Sarkar, D.; Rakshit, A.; Parewa, H.P.; Danish, S.; Alfarraj, S.; Datta, R. Bio-priming with compatible rhizospheric microbes enhances growth and micronutrient uptake of red cabbage. Land 2022, 11, 536. [Google Scholar] [CrossRef]
- Farina, R.; Beneduzi, A.; Ambrosini, A. Diversity of plant growth-promoting rhizobacteria communities associated with the stages of canola growth. App. Soil Ecol. 2012, 55, 44–52. [Google Scholar] [CrossRef]
- Bacon, C.; Hinton, D. Bacterial endophytes: The endophytic niche, its occupants, and its utility. In Plant-Associated Bacteria; Gnanamanickam, S.S., Ed.; Springer: Dordrecht, The Netherlands, 2007; Volume 3, pp. 155–194. [Google Scholar]
- Sarkar, D.; Rakshit, A. Amalgamation of farmers’ bio-priming knowledge in integrated nutrient management for sustainable management of red cabbage soil under Middle Gangetic Plains, India. Environ. Manag. 2022. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.; Webb, J.S.; Kjelleberg, S. Microbial colonization and competition on the marine alga Ulva australis. Appl. Environ. Microbiol. 2006, 72, 5547–5555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, R.; Baraniya, D.; Wang, Y.F.; Kelkar, A.; Meena, R.S.; Yadav, G.S.; Teresa Ceccherini, M.; Formanek, P. Amino acid: Its dual role as nutrient and scavenger of free radicals in soil. Sustainability 2017, 9, 1402. [Google Scholar] [CrossRef] [Green Version]
- Holátko, J.; Brtnický, M.; Kučerík, J.; Kotianová, M.; Elbl, J.; Kintl, A.; Kynický, J.; Benada, O.; Datta, R.; Jansa, J. Glomalin–Truths, myths, and the future of this elusive soil glycoprotein. Soil Biol. Biochem. 2021, 153, 108116. [Google Scholar] [CrossRef]
- Sarkar, D.; Rakshit, A.; Al-Turki, A.I.; Sayyed, R.Z.; Datta, R. Connecting bio-priming approach with integrated nutrient management for improved nutrient use efficiency in crop species. Agriculture 2021, 11, 372. [Google Scholar] [CrossRef]
- Verma, S.C.; Singh, A.; Chowdhury, S.P.; Tripathi, A.K. Endophytic colonization ability of two deep-water rice endophytes, Pantoea sp. and Ochrobactrum sp. using green fluorescent protein reporter. Biotechnol. Lett. 2004, 26, 425–429. [Google Scholar] [CrossRef]
- Chowdhury, S.; Schmid, M.; Hartmann, A.; Tripathi, A. Identification of diazotrophs in the culturable bacterial community associated with roots of Lasiurus sindicus, a perennial grass of thar desert, India. Microbial Ecol. 2007, 54, 82–90. [Google Scholar] [CrossRef]
- Zehr, J.P.; Jenkins, B.D.; Short, S.M.; Steward, G.F. Nitrogenase gene diversity and microbial community structure: A cross-system comparison. Environ. Microbiol. 2003, 5, 539–554. [Google Scholar] [CrossRef]
- Dean, D.R.; Jacobson, M.R. Biochemical genetics of nitrogenase. In Biological Nitrogen Fixation; Stacey, G., Burris, R.H., Evans, H.J., Eds.; Chapman & Hall: New York, NY, USA, 1992; pp. 763–834. [Google Scholar]
- Lim, C.W.; Park, J.Y.; Lee, S.H.; Hwang, C.H. Comparative proteomic analysis of soybean nodulation using a supernodulation mutant, SS2-2. Biosci. Biotech. Biochem. 2010, 74, 2396–2404. [Google Scholar] [CrossRef] [Green Version]
- Den Herder, J.; Lievens, S.; Rombauts, S.; Holsters, M.; Goormachtig, S. A symbiotic plant peroxidase involved in bacterial invasion of the tropical legume Sesbania rostrata. Plant Physiol. 2007, 144, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Naffaa, W.; Ravel, C.; Boyer, N.; Guillaumin, J.-J. Peroxidase activity of perennial ryegrass and tall fescue seedlings artificially infected with endophytes. Agronomie 1999, 19, 611–619. [Google Scholar] [CrossRef]
- Mitra, R.M.; Long, S.R. Plant and bacterial symbiotic mutants define three transcriptionally distinct stages in the development of the Medicago truncatula/Sinorhizobium meliloti symbiosis. Plant Physiol. 2004, 134, 595–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stintzi, A.; Heitz, T.; Prasad, V.; Wiedemann-Merdinoglu, S.; Kauffmann, S.; Geoffroy, P.; Legrand, M.; Fritig, B. Plant ‘pathogenesis-related’proteins and their role in defense against pathogens. Biochimie 1993, 75, 687–706. [Google Scholar] [CrossRef]
- Staehelin, C.; Schultze, M.; Kondorosi, É.; Mellor, R.B.; Boiler, T.; Kondorosi, A. Structural modifications in Rhizobium meliloti Nod factors influence their stability against hydrolysis by root chitinases. Plant J. 1994, 5, 319–330. [Google Scholar] [CrossRef]
- Camehl, I.; Oelmüller, R. Do ethylene response factors 9 and-14 repress PR gene expression in the interaction between Piriformospora indica and Arabidopsis? Plant Sig. Behav. 2010, 5, 932–936. [Google Scholar] [CrossRef] [Green Version]
- Lane, B.G. Oxalate, Germins, and Higher-Plant Pathogens. IUBMB Life 2002, 53, 67–75. [Google Scholar] [CrossRef]
- Miché, L.; Battistoni, F.; Gemmer, S.; Belghazi, M.; Reinhold-Hurek, B. Upregulation of jasmonate-inducible defense proteins and differential colonization of roots of Oryza sativa cultivars with the endophyte Azoarcus sp. Mol. Plant-Microbe Interact. 2006, 19, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, S.; Sato, F. Plant pathogenesis-related proteins: Molecular mechanisms of gene expression and protein function. J. Biochem. 1999, 125, 1–8. [Google Scholar] [CrossRef]
- Anelli, T.; Sitia, R. Protein quality control in the early secretory pathway. EMBO 2008, 27, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Tintor, N.; Lu, X.; Rauf, P.; Pajerowska-Mukhtar, K.; Häweker, H.; Dong, X.; Robatzek, S.; Schulze-Lefert, P. Receptor quality control in the endoplasmic reticulum for plant innate immunity. EMBO J. 2009, 28, 3439–3449. [Google Scholar] [CrossRef] [Green Version]
- Millet, Y.A.; Danna, C.H.; Clay, N.K.; Songnuan, W.; Simon, M.D.; Werck-Reichhart, D.; Ausubel, F.M. Innate immune responses activated in Arabidopsis roots by microbe-associated molecular patterns. Plant Cell Online 2010, 22, 973–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshmanan, V.; Kitto, S.L.; Caplan, J.L.; Hsueh, Y.-H.; Kearns, D.B.; Wu, Y.-S.; Bais, H.P. Microbe-associated molecular patterns-triggered root responses mediate beneficial rhizobacterial recruitment in Arabidopsis. Plant Physiol. 2012, 160, 1642–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (DAS) | Shannon’s Index (H) | Evenness (eH/S) | Dominance (D) |
|---|---|---|---|
| 0 (Seeds) | 1.0890 | 0.9903 | 0.3400 |
| 7 | 0.0000 | 1.0000 | 1.0000 |
| 14R | 1.1270 | 0.5146 | 0.4683 |
| 14S | 0.3046 | 0.6781 | 0.8347 |
| 21R | 1.7990 | 0.6714 | 0.2256 |
| 21S | 0.4506 | 0.7846 | 0.7222 |
| 28R | 0.1446 | 0.3852 | 0.9463 |
| 28S | 0.0000 | 1.0000 | 1.0000 |
| 35R | 0.1888 | 0.6039 | 0.9109 |
| 35S | 0.0000 | 1.0000 | 1.0000 |
| 42R | 0.0000 | 1.0000 | 1.0000 |
| 42S | 0.0000 | 1.0000 | 1.0000 |
| 49R | 0.0938 | 0.5492 | 0.9629 |
| 49S | 0.5890 | 0.9011 | 0.6005 |
| 56R | 0.1976 | 0.6092 | 0.9056 |
| 56(L)S | 0.3514 | 0.7105 | 0.8006 |
| 56(U)S | 0.8439 | 0.7751 | 0.4588 |
| 63R | 1.3910 | 0.8036 | 0.2764 |
| 63(L)S | 0.7724 | 0.7217 | 0.5678 |
| 63(M)S | 1.9810 | 0.7254 | 0.1667 |
| 63(U)S | 1.2680 | 0.7106 | 0.3504 |
| 70R | 1.1600 | 0.6377 | 0.4380 |
| 70(L)S | 0.0000 | 1.0000 | 1.0000 |
| 70(M)S | 0.0000 | 1.0000 | 1.0000 |
| 70(U)S | 0.9652 | 0.8751 | 0.4289 |
| Isolate No. | IAA (µg/mL) | Phosphate-Solubilization (mM/mL) | % Siderophore (Typing) | ARA (nmol Ethylene mg Protein−1 hr−1) | nifH |
|---|---|---|---|---|---|
| PM03 | - | 9.4 | - | 40.24 | - |
| PM05 | 10.74 ± 0.49 | - | 12.8 (H) | 7.70 ± 0.20 | - |
| PM16 | - | 2.5 | 2.5 (H) | 21.09 ± 3.80 | + |
| PM219 | 11.80 ± 0.57 | - | - | 30.36 ± 2.54 | + |
| PM221 | 73.70 ± 0.92 | 6.4 | 0.3 (C) | 24.84 ± 5.60 | - |
| PM223 | - | - | - | 35.53 | - |
| PM224 | 80.25 ± 2.10 | 7.5 | - | 26.22 | - |
| PM228 | - | - | - | 32.40 | + |
| PM232 | 19.76 ± 0.90 | 6.8 | 14.2 (C) | 26.28 | - |
| PM251 | - | - | - | 3.45 | - |
| PM253 | 81.30 ± 3.39 | 6.0 | 6.77 (C) | 28.84 | - |
| PM366 | 8.58 ± 0.93 | - | 3 (H) | 30.23 | - |
| PM368 | - | - | 0.5 (C) | 4.55 | - |
| PM376 | 9.40 ± 0.63 | - | 63.5 (C) (H) | 10.09 | - |
| PM377 | - | - | 78.5 (C) (H) | 9.78 | - |
| PM379 | - | - | - | 45.83 ± 2.57 | - |
| PM380 | - | - | - | 20.84 ± 4.13 | + |
| PM381 | - | - | - | 23.20 | - |
| PM386 | 67.30 ± 2.47 | 8.9 | - | 18.98 | - |
| PM389 | - | V.L. | 0.6 (C) (H) | 28.91 ± 1.86 | + |
| PM4106 | 50.35 ± 0.50 | - | - | 20.34 | - |
| PM4107 | - | V.L. | 13 (H) | 25.68 | - |
| PM5143 | 14.09 ± 1.05 | - | - | 9.87 | + |
| PM5153 | 4.47 ± 0.40 | - | 0.3 (H) | 8.87 | + |
| PM7220 | - | - | - | 5.67 | - |
| PM7265 | - | V.L. | 13.43 (H) | 26.00 | - |
| PM8311 | 3.40 ± 0.17 | - | - | 6.72 ± 2.56 | + |
| PM8335 | - | - | - | 31.79 | - |
| PM8337 | 15.47 ± 2.45 | 5.9 | 11.7 | 22.98 | + |
| PM8342 | 31.81 ± 2.20 | 2.9 | 13.05 | 29.98 | - |
| PM9363 | - | - | - | 2.98 | + |
| PM9364 | - | - | - | 1.83 ± 0.87 | + |
| PM9365 | - | - | - | 34.63 ± 1.49 | - |
| PM9366 | - | 2.5 | 5.3 (H) | 28.89 | - |
| PM9367 | - | - | - | 20.79 | - |
| PM9368 | - | - | - | 10.96 ± 3.33 | - |
| PM9369 | - | - | - | 18.94 | - |
| PM9370 | 17.78 ± 1.20 | - | - | 4.32 | - |
| PM9376 | - | - | - | 15.67 | - |
| PM9383 | - | - | - | 7.58 | - |
| PM9399 | - | - | - | 28.89 | - |
| PM9400 | 6.91 ± 2.10 | - | - | 6.86 | - |
| PM9401 | 15.43 ± 0.32 | - | - | 26.78 | - |
| PM9402 | - | - | - | 24.32 ± 6.20 | - |
| PM9403 | - | - | - | 13.58 | - |
| PM9404 | - | - | 0.5 (H) | 56.03 ± 4.66 | - |
| PM9405 | 10.77 ± 1.98 | - | - | 95.56 ± 0.92 | - |
| PM9406 | - | - | - | 4.58 | - |
| PM9408 | 6.50 ± 1.15 | - | 8.8 (H) | 16.43 ± 2.83 | - |
| PM9409 | - | - | - | 13.08 | - |
| PM9410 | - | - | - | 5.90 | - |
| PM9411 | 7.89 ± 0.50 | - | - | 5.25 | - |
| PM9424 | - | - | - | 30.90 | - |
| PM9426 | 8.99 ± 2.78 | - | - | 32.96 ± 1.86 | - |
| PM9427 | 10.23 ± 0.00 | - | - | 30.34 | - |
| PM9429 | - | - | - | 18.65 | - |
| PM9430 | - | - | - | 16.90 | - |
| PM9432 | - | - | - | 8.92 | - |
| PM9433 | - | - | - | 7.89 | - |
| PM9435 | 41.49 ± 0.11 | - | - | 14.56 | - |
| PM9436 | 17.62 ± 3.25 | - | - | 5.67 | - |
| PM9437 | 15.85 ± 0.5 | - | - | 100.89 | - |
| PM9438 | 14.86 ± 1.79 | - | - | 31.86 ± 5.06 | - |
| PM10455 | 12.08 ± 0.32 | - | - | 128.36 ± 3.42 | + |
| PM10458 | - | - | - | 25.43 | - |
| PM10459 | - | - | - | 15.98 | - |
| PM10461 | 19.48 ± 1.79 | - | - | 103.93 ± 1.99 | - |
| PM10462 | 6.34 ± 0.95 | 9.1 | - | 5.68 | - |
| PM10463 | - | - | - | 15.02 | - |
| PM10476 | - | - | - | 8.98 | - |
| PM10478 | - | - | - | 15.67 | - |
| PM10490 | - | V.L. | 3.5 (H) | 23.34 | - |
| PM10491 | - | V.L. | 8.8 (H) | 25.67 | - |
| PM10503 | - | - | - | 47.11 ± 6.22 | + |
| PM10509 | - | - | - | 18.96 | - |
| PM10510 | - | - | - | 57.73 ± 6.9 | - |
| Isolates | Root Length (cm) | Shoot Length (cm) | Fresh Wt. (g) | Vigour Index |
|---|---|---|---|---|
| Control | 2.9 ± 1.0 | 6.7 ± 0.9 | 0.08 ± 0.02 | 960 |
| PM9438 | 3.4 ± 1.9 (17%) * | 7.6 ± 1.9 (13%) | 0.10 ± 0.05 (35%) | 1100 (15%) * |
| PM10455 | 3.65 ± 1.3 (26%) * | 8.2 ± 1.7 (22%) * | 0.15 ± 0.09 (88%) * | 1185 (23%) * |
| PM380 | 3.1 ± 1.6 (7%) | 8.45 ± 1.5 (25%) * | 0.13 ± 0.03 (63%) * | 1155 (20%) * |
| PM389 | 3.5 ± 0.1 (21%) * | 7.9 ± 1.3 (18%) * | 0.1 ± 0.02 (25%) * | 1140 (19%) * |
| PM9426 | 3.8 ± 1.5 (31%) * | 9.6 ± 2.2 (43%) * | 0.10 ± 0.03 (25%) * | 1340 (40%) * |
| PM8311 | 3.3 ± 1.3 (14%) * | 6.9 ± 1.7 (3%) * | 0.08 ± 0.03 (0%) | 1020 (6%) |
| PM9364 | 3.32 ± 1.5 (14%) * | 6.9 ± 2.4 (3%) * | 0.08 ± 0.02 (0%) | 1022 (6%) |
| PM9402 | 4 ± 1.8 (38%) * | 7.8 ± 1.0 (16%) * | 0.09 ± 0.01 (13%) * | 1180 (23%) * |
| PM9363 | 3.7 ± 0.9 (28%) * | 8.2 ± 1.0 (22%) * | 0.11 ± 0.02 (38%) * | 1190 (24%) * |
| PM9368 | 3.7 ± 1.3 (28%) * | 7.1 ± 1.3 (5%) | 0.09 ± 0.02 (13%) | 1080 (23%) * |
| PM9405 | 3.8 ± 1.2 (31%) * | 7.8 ± 1.2 (16%) * | 0.11 ± 0.02 (38%) * | 1160 (21%) * |
| PM10461 | 3.4 ± 1.1 (17%) | 7.6 ± 1.5 (13%) * | 0.09 ± 0.03 (13%) * | 1100 (15%) * |
| PM9404 | 3.7 ± 1.0 (28%) * | 7.6 ± 1.0 (13%) * | 0.10 ± 0.02 (25%) * | 1130 (18%) * |
| PM9408 | 3.7 ± 1.4 (28%) * | 7.5 ± 1.2 (12%) * | 0.10 ± 0.02 (25%) * | 1120 (17%) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, G.; Paul, S.; Singh, S.; Pietramellara, G.; Pathan, S.I.; Danish, S.; Jabborova, D.; Datta, R.; Jha, P.N. Exploring Functional Diversity and Community Structure of Diazotrophic Endophytic Bacteria Associated with Pennisetum glaucum Growing under Field in a Semi-Arid Region. Land 2022, 11, 991. https://doi.org/10.3390/land11070991
Gupta G, Paul S, Singh S, Pietramellara G, Pathan SI, Danish S, Jabborova D, Datta R, Jha PN. Exploring Functional Diversity and Community Structure of Diazotrophic Endophytic Bacteria Associated with Pennisetum glaucum Growing under Field in a Semi-Arid Region. Land. 2022; 11(7):991. https://doi.org/10.3390/land11070991
Chicago/Turabian StyleGupta, Garima, Sangeeta Paul, Sachidanand Singh, Giacomo Pietramellara, Shamina Imran Pathan, Subhan Danish, Dilfuza Jabborova, Rahul Datta, and Prabhat Nath Jha. 2022. "Exploring Functional Diversity and Community Structure of Diazotrophic Endophytic Bacteria Associated with Pennisetum glaucum Growing under Field in a Semi-Arid Region" Land 11, no. 7: 991. https://doi.org/10.3390/land11070991