
Horizontal Distribution Characteristics and Environmental Factors of Shrubland Species Diversity in Hainan Island, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
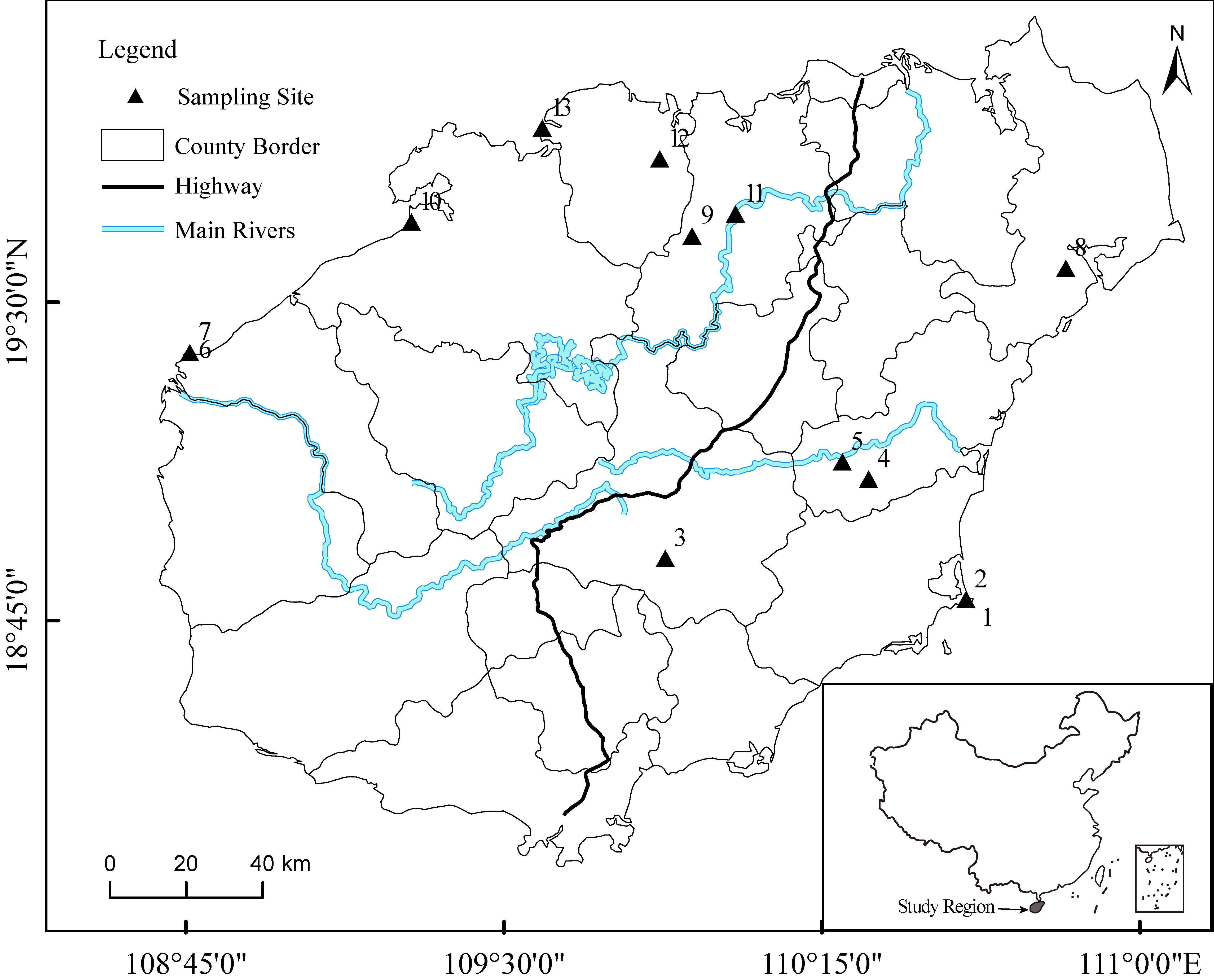
2.1. Study Site
2.2. Community Surveys
2.3. Soil Physicochemical Properties
2.4. Analytical Approach
2.4.1. Measurement of Shrub Community Diversity
2.4.2. Data Processing and Analysis
3. Results
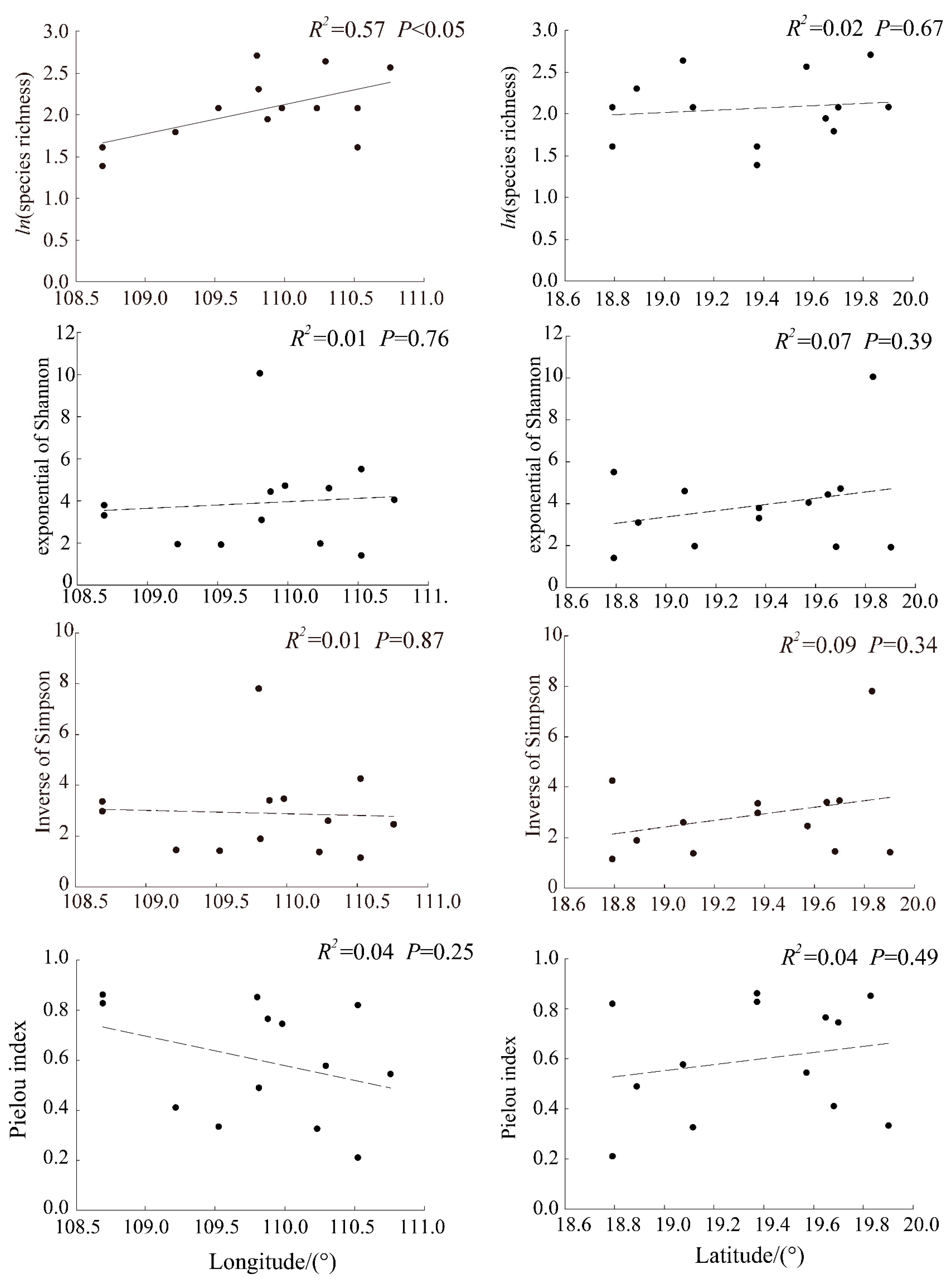
3.1. Horizontal Differentiation of Species Diversity Index
3.2. The Correlation between Species Diversity Index and Environmental Factors
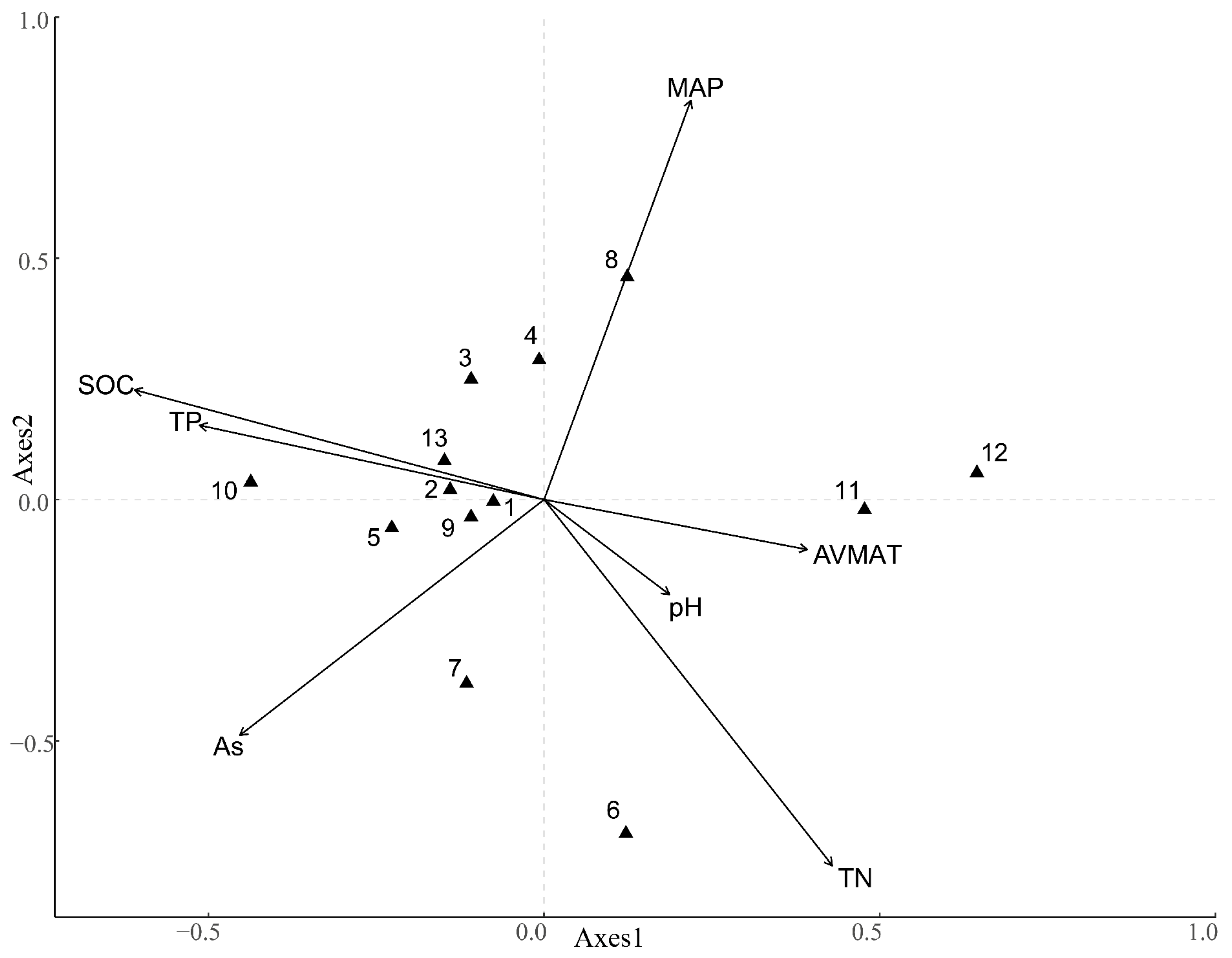
3.3. The Sequencing Analysis of Species Diversity and Environmental Factors
4. Discussion
4.1. The Horizontal Distribution of Shrub Species Diversity in Hainan
4.2. The Relationship between Shrub Species Diversity and Hydrothermal Factors in Hainan
4.3. The Relationship between Shrub Species Diversity and Edaphic Factors in Hainan
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, D.Q. Biodiversity conservation strategies and priority area analysis in China. In Research and Methods in Ecohealth and Conservation; GDRI Ecosystem Health and Environmental Disease Ecology: Kunming, China, 2016. [Google Scholar]
- Pritchard, H.W.; Moat, J.F.; Ferraz, J.B.S.; Marks, T.R.; Camargo, J.L.C.; Nadarajan, J.; Ferraz, I.D.K. Innovative approaches to the preservation of forest trees. For. Ecol. Manag. 2014, 333, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.Y.; Wang, Z.H.; Zheng, C.Y.; Fang, J.Y. Biodiversity in China’s mountains. Front. Ecol. Environ. 2006, 4, 347–352. [Google Scholar] [CrossRef]
- Zhao, G.S.; Liu, J.Y.; Kuang, W.H.; Ouyang, Z.; Xie, Z.L. Disturbance impacts of land use change on biodiversity conservation priority areas across China: 1990–2010. J. Geogr. Sci. 2015, 25, 515–529. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Uusivuori, J.; Kuuluvainen, J. Econometric analysis of the causes of forest land use changes in Hainan, China. Can. J. For. Res. 2000, 30, 1913–1921. [Google Scholar] [CrossRef]
- Guo, J.; Gong, P.; Dronova, I.; Zhu, Z. Forest cover change in China from 2000 to 2016. Int. J. Remote Sens. 2022, 43, 593–606. [Google Scholar] [CrossRef]
- Mi, X.C.; Feng, G.; Hu, Y.B.; Zhang, J.; Chen, L.; Corlett, R.T.; Hughes, A.C.; Pimm, S.; Schmid, B.; Shi, S.H.; et al. The global significance of biodiversity science in China: An overview. Natl. Sci. Rev. 2021, 8, nwab032. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Qu, H.; Liu, R.; Zhang, Z.X. Vulnerability of forest vegetation to anthropogenic climate change in China. Sci. Total Environ. 2018, 621, 1633–1641. [Google Scholar] [CrossRef]
- Zhang, Z.J.; He, J.S.; Li, J.S.; Tang, Z.Y. Distribution and conservation of threatened plants in China. Biol. Conserv. 2015, 192, 454–460. [Google Scholar] [CrossRef]
- Liu, H.D.; Chen, Q.; Liu, X.; Xu, Z.Y.; Dai, Y.C.; Liu, Y.; Chen, Y.F. Variation patterns of plant composition/diversity in Dacrydium pectinatum communities and their driving factors in a biodiversity hotspot on Hainan Island, China. Glob. Ecol. Conserv. 2020, 22, e01034. [Google Scholar] [CrossRef]
- Zhu, Z.X.; Nizamani, M.M.; Sahu, S.K.; Kunasingam, A.; Wang, H.F. Tree abundance, richness, and phylogenetic diversity along an elevation gradient in the tropical forest of Diaoluo Mountain in Hainan, China. Acta Oecol. 2019, 101, 103481. [Google Scholar] [CrossRef]
- Zhu, L.; Sun, O.J.; Sang, W.G.; Li, Z.Y.; Ma, K.P. Predicting the spatial distribution of an invasive plant species (Eupatorium adenophorum) in China. Landsc. Ecol. 2007, 22, 1143–1154. [Google Scholar] [CrossRef]
- Kratochwil, A. Biodiversity in ecosystems: Some principles. In Tasks for Vegetation Science; Springer: Dordrecht, The Netherlands, 1999; Volume 34, pp. 5–38. [Google Scholar]
- Duffy, J.E.; Godwin, C.M.; Cardinale, B.J. Biodiversity effects in the wild are common and as strong as key drivers of productivity. Nature 2017, 549, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.F.; Mirotchnick, N.; Jain, M.; Palmer, M.I.; Naeem, S. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships. Ecology 2011, 92, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Palmer, M.W. The coexistence of species in fractal landscapes. Am. Nat. 1992, 139, 375–397. [Google Scholar] [CrossRef]
- Xu, M.; Ma, L.; Jia, Y.Y.; Liu, M. Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS ONE 2017, 12, e0174231. [Google Scholar] [CrossRef]
- Stomp, M.; Huisman, J.; Mittelbach, G.G.; Litchman, E.; Klausmeier, C.A. Large-scale biodiversity patterns in freshwater phytoplankton. Ecology 2011, 92, 2096–2107. [Google Scholar] [CrossRef]
- Wu, A.C.; Deng, X.W.; Ren, X.L.; Xiang, W.H.; Zhang, L.; Ge, R.; Niu, Z.E.; He, H.L.; He, L.J. Biogeographic patterns and influencing factors of the species diversity of tree layer community in typical forest ecosystems in China. Acta Ecol. Sin. 2018, 38, 7727–7738. [Google Scholar]
- Fang, J.M. Spatial patterns of species diversity of seed plants in China and their climatic explanation. Biodiv. Sci. 2008, 16, 470. [Google Scholar] [CrossRef]
- Shi, J.X.; Xu, M.S.; Fang, X.C.; Zheng, L.T.; Zhang, Y.; Bao, D.F.; Yang, A.N.; Yan, E.R. Latitudinal variability and driving factors of functional diversity in Pinus thunbergii communities across sea-islands in Eastern China. Chin. J. Plant Ecol. 2021, 45, 163–173. Available online: https://www.plant-ecology.com/EN/Y2021/V45/I2/163 (accessed on 3 July 2022). [CrossRef]
- Vazquez, J.A.; Garnish, T.J. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Mazatlán. J. Ecol. 1998, 86, 999–1020. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Q.; Chen, D.D.; Shai, W.T.; Zhao, L.; Xu, S.X.; Zhao, X.Q. Patterns of plant species diversity along an altitudinal gradient and its effect on above-ground biomass in alpine meadows in Qinghai-Tibet Plateau. Biodiv. Sci. 2015, 23, 451–462. [Google Scholar] [CrossRef]
- Li, M.J.; He, Z.S.; Jiang, L.; Gu, X.G.; Jin, M.R.; Chen, B.; Liu, J.F. Distribution pattern and driving factors of species diversity and phylogenetic diversity along altitudinal gradient on the south slope of Daiyun Mountain. Acta Ecol. Sin. 2021, 41, 1148–1157. [Google Scholar] [CrossRef]
- Xu, Y.J.; Chen, Y.N.; Li, W.H.; Fu, A.H.; Ma, X.D.; Gui, D.W.; Chen, Y.P. Distribution pattern and environmental interpretation of plant species diversity in the mountainous region of Ili River Valley, Xinjiang, China. Chin. J. Plant Ecol. 2010, 34, 1142–1154. [Google Scholar]
- Qin, H.; Zhang, Y.B.; Dong, G.; Zhang, F. Altitudinal patterns of taxonomic, phylogenetic and functional diversity of f-orest communities in Mount Guandi, Shanxi, China. Chin. J. Plant Ecol. 2019, 43, 762–773. [Google Scholar] [CrossRef]
- Weng, C.L.; Zhang, T.T.; Wu, D.H.; Chen, S.W.; Jin, Y.; Ren, H.B.; Yu, M.J.; Luo, Y.Y. Drivers and patterns of α- and β-diversity in ten main forest community types in Gutianshan, eastern China. Biodiv. Sci. 2019, 27, 33–41. [Google Scholar] [CrossRef]
- Zhao, S.Q.; Fang, J.Y.; Zong, Z.J.; Zhu, B.; Shen, H.H. Composition, structure and species diversity of plant commun-ities along an altitudinal gradient on the northern slope of Mt. Changbai, Northeast China. Biodiv. Sci. 2004, 12, 164–173. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Fang, J.Y. A review on the elevational patterns of plant species diversity. Biodiv. Sci. 2004, 12, 20–28. [Google Scholar] [CrossRef]
- Pianka, E.R. Latitudinal gradients in species diversity-a review of concepts. Am. Nat. 1966, 100, 33–46. [Google Scholar] [CrossRef]
- O’brien, E. Water-energy dynamics, climate, and predicting of woody plant species richness: An interim general model. J. Biogeogr. 1998, 25, 379–398. [Google Scholar] [CrossRef]
- Burke, A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. J. Veg. Sci. 2001, 12, 53. [Google Scholar] [CrossRef]
- Siefert, A.; Ravenscroft, C.; Althoff, D.; Alvarez, Y.; Carter, C.; Glennon, B.E.; Kelsey, L.; Heberling, J.; Mason, J.; Pontes, A.; et al. Scale dependence of vegetation-environment relationships: A meta-analysis of multivariate data. J. Veg. Sci. 2012, 23, 942–951. [Google Scholar] [CrossRef]
- Fang, J.Y.; Li, Y.D.; Zhu, B.; Liu, G.Y. Community structures and species richness in the montane rain forest of Jianfengling, Hainan Island, China. Biodiv. Sci. 2004, 12, 29–43. [Google Scholar] [CrossRef]
- Li, Y.P.; Xu, H.; Li, Y.D.; Luo, T.S.; Chen, D.X.; Zhou, Z.; Lin, M.X.; Yang, H.A. Scale-dependent spatial patterns of species diversity in the tropical montane rain forest in Jianfengling, Hainan Island, China. Chin. J. Plant Ecol. 2016, 40, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.X.; Long, W.X.; Yang, X.B.; Xiong, M.H.; Kang, Y.; Huang, J.; Hong, Z.L.; Zhou, Z.L.; Lu, Y.Q.; Fang, J.; et al. Patterns of plant diversity within and among three tropical cloud forest communities in Hainan Island. Chin. J. Plant Ecol. 2016, 40, 469–479. [Google Scholar] [CrossRef]
- Chen, G.D.; Tong, S.Y.; Du, S.J.; Zhong, S.Y.; Fu, R.; Wu, H.X.; Yu, X.B. Subforest species diversity and dominant population distribution pattern of secondary forest community under different felling lianas treatments. J. West China For. Sci. 2020, 49, 54–60. [Google Scholar]
- Hu, H.F.; Wang, Z.H.; Liu, G.H.; Fu, B.J. Vegetation carbon storage of major shrublands in China. Chin. J. Plant Ecol. 2006, 30, 539–544. [Google Scholar] [CrossRef]
- Spicer, R.A.; Herman, A.B.; Liao, W.; Spicer, T.E.V.; Kodrul, T.M.; Yang, J.; Jin, J.H. Cool tropics in the Middle Eocene: Evidence from the Changchang Flora, Hainan Island, China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 412, 1–16. [Google Scholar] [CrossRef]
- Luo, W.Q.; Fu, S.H.; Yang, X.B.; Chen, Y.K.; Zhou, W.; Yang, Q.; Tao, C.; Zhou, W.S. Distribution patterns of alien invasive plants and their influences on native plants of Hainan Island. Chin. J. Plant Ecol. 2015, 39, 486–500. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Ed.; SSSA: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar] [CrossRef]
- Pierzynski, G.M. Methods of Phosphorus Analysis for Soils, Sediments, Residuals, and Waters; North Carolina State University: Manhattan, KS, USA, 2000. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Constable and Co. Ltd.: London, UK, 1962. [Google Scholar]
- Hurlbert, S.H. The non-concept of species diversity: A critique and alternative parameters. Ecology 1971, 52, 577–586. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’ Index. Glob. Ecol. Biogeogr. 2013, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.Y.; Wang, X.P.; Shen, Z.H.; Tang, Z.Y.; He, J.S.; Yu, D.; Jiang, Y.; Wang, Z.H.; Zheng, C.Y.; Zhu, J.L.; et al. Methods and protocols for plant community inventory. Biodiv. Sci. 2009, 17, 533–548. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.H.; Jost, L. Unifying species diversity, phylogenetic diversity, functional diversity, and related similarity and differentiation measures through Hill numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Roberts, D.W.; Cooper, S.V. Concepts and techniques of vegetation mapping. In Proceedings of the Land Classifications Based on Vegetation: Applications for Resource Management, Moscow, ID, USA, 17–19 November 1987; pp. 90–96. [Google Scholar]
- Von, W.H.; Hanspach, J.; Bruelheide, H.; Wesche, K. Pluralism and diversity: Trends in the use and application of ordination methods 1990–2007. J. Veg. Sci. 2009, 20, 695–705. [Google Scholar] [CrossRef]
- Kucuk, A.; Clare, A.G.; Jones, L. An estimation of the surface tension for silicate glass melts at 1400 C using statistical analysis. Glass Technol. 1999, 40, 149–153. [Google Scholar]
- Patiño, J.; González-Mancebo, J.M. Exploring the effect of host tree identity on epiphyte bryophyte communities in different Canarian subtropical cloud forests. Plant Ecol. 2011, 212, 433–449. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y.D.; Luo, T.S.; Chen, D.X.; Lin, M.X. Environmental factors correlated with species diversity in different tropical rain forest types in Jianfengling, Hainan Island, China. Chin. J. Plant Ecol. 2013, 37, 26–36. [Google Scholar] [CrossRef]
- Deng, L.P.; Bai, X.J.; Qin, S.J.; Wei, Y.W.; Zhou, Y.B.; Li, L.L.; Niu, S.S.; Han, M.N. Spatial distribution and scale effect of species diversity of secondary forests in montane region of eastern Liaoning Province, China. Chin. J. Appl. Ecol. 2016, 27, 2197–2204. [Google Scholar] [CrossRef]
- Strong, W.L. Biased richness and evenness relationships within Shannon–Wiener index values. Ecol. Indic. 2016, 67, 703–713. [Google Scholar] [CrossRef]
- Hill, T.C.J.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.L.; Yang, T.; Liu, H.Y.; Kuang, M.Y.; Wang, R.Q.; Zhang, F.; Gao, X.M.; Yue, M.; Zhang, M.; Zheng, P.F.; et al. Distribution and species diversity patterns of Vitex negundo var. heterophylla Shrublands in North China. Chin. J. Plant Ecol. 2019, 43, 825–833. [Google Scholar] [CrossRef]
- Ma, B.; Zhou, Z.Y.; Zhang, L.L.; Gao, W.X.; Chen, S.K.; Zhang, B.L. The spatial distribution characteristics of plant diversity in Alex Left Banner. Acta Ecol. Sin. 2008, 28, 6099–6106. [Google Scholar]
- Lü, L.S.; Cai, H.Y.; Yang, Y.; Wang, Z.H.; Zeng, H. Geographic patterns and environmental determinants of gymnosperm species diversity in China. Biodiv. Sci. 2018, 26, 1133–1146. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.B.; Ji, S.Y.; Chen, F.P. A review on the species diversity of plant community. Chin. J. Ecol. 2001, 20, 55–60. [Google Scholar]
- Frelich, L.E.; Calcote, R.R.; Davis, M.B.; Pastor, J. Patch formation and maintenance in an old-growth hemlock-hardwood forest. Ecology 1993, 74, 513–527. [Google Scholar] [CrossRef]
- Gong, H.D.; Yang, G.P.; Lu, Z.Y.; Liu, Y.H. Diversity and spatial distribution patterns of trees in an evergreen broad-leaved forest in the Ailao Mountains, Yunnan. Biodiv. Sci. 2011, 19, 143–150. [Google Scholar] [CrossRef]
- He, J.S. Plant species diversity of the degraded ecosystems in the three gorges region. Acta Ecol. Sin. 1998, 18, 399–407. [Google Scholar]
- Sha, W.; Dong, S.K.; Liu, S.L.; Liu, Q.R.; Shi, J.B.; Li, X.W.; Su, X.K.; Wu, Y. Spatial patterns of plant community biomass and species diversity in Aerjin Mountain Nature Reserve and their influencing factors. Chin. J. Ecol. 2016, 35, 330–337. [Google Scholar]
- Wang, A.S.; Ma, J.J.; Gong, H.D.; Fan, G.A.; Wang, M.; Zhao, H.M.; Cheng, J.H. Patterns and drivers of species richness of early spring annual ephemeral plants in northern Xinjiang. Biodiv. Sci. 2021, 29, 735–745. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Crick, J.C.; Grime, J.P. Morphological plasticity and mineral nutrient capture in 2 herbaceous species of contrasted ecology. N. Phytol. 1987, 107, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Brosofske, K.D.; Chen, J.; Crow, T.R. Understory vegetation and site factors: Implications for a managed Wisconsin landscape. For. Ecol. Manag. 2001, 146, 75–87. [Google Scholar] [CrossRef]
- Zhou, R.H.; Su, T.C.; Yu, J.; Xiang, L.; Chen, C.L.; Zhang, H.W.; Li, J.; Hao, J.F. Species diversity and soil physicochemical properties of different communities in Bifengxia evergreen broad-leaved forest. Chin. J. Ecol. 2021, 41, 1–8. [Google Scholar]
- Zheng, L.T.; Su, T.; Liu, X.Y.; Yin, F.; Guo, C.; Tuo, B.; Yan, E.R. Species, functional, structural diversity of typical plant communities and their responses to environmental factors in Miao Archipelago, China. Chin. J. Appl. Ecol. 2018, 29, 343–351. [Google Scholar]
- Li, C.Y.; Zhang, W.J.; Lai, Z.M.; Peng, F.; Chen, X.J.; Xue, X.; Wang, T.; You, Q.G.; Du, H.Q. Plant productivity, species diversity, soil properties, and their relationships in an alpine steppe under different degradation degrees at the source of the Yellow River. Acta Ecol. Sin. 2021, 41, 4541–4551. [Google Scholar]
- Long, W.X.; Yang, X.D.; Li, D.H. Patterns of species diversity and soil nutrients along a chronosequence of vegetation recovery in Hainan Island, South China. Ecol. Res. 2012, 27, 561–568. [Google Scholar] [CrossRef]
- Zou, Y.J. Study on Distribution Characteristics of Organic Carbon and Total Nitrogen in Shrub Ecosystem of Hainan Island; Hainan University: Hainan, China, 2017. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Plot No. | Latitude | Longitude | Elevation (m−1) | Exposure (°) | Slope (°) | Dominant Species |
|---|---|---|---|---|---|---|
| 1 | 18.79 | 110.53 | 15 | 245 | 10 | Embelia laeta, Flacourtia indica |
| 2 | 18.81 | 110.52 | 235 | 160 | 25 | Pandanus tectorius |
| 3 | 18.89 | 109.81 | 347 | 40 | 30 | Rhodomyrtus tomentosa |
| 4 | 19.07 | 110.29 | 56 | 280 | 5 | Rhodomyrtus tomentosa |
| 5 | 19.12 | 110.23 | 18 | 147 | 3 | Flueggea virosa |
| 6 | 19.37 | 108.69 | 5 | 208 | 0 | Opuntia, Clerodendrum inerme |
| 7 | 19.38 | 108.68 | 8 | 208 | 0 | Opuntia, Pandanus tectorius |
| 8 | 19.57 | 110.76 | 20 | 140 | 5 | Mallotus paniculatus |
| 9 | 19.65 | 109.88 | 110 | 118 | 5 | Rhodomyrtus tomentosa, Ternstroemia microphylla |
| 10 | 19.68 | 109.21 | 4 | 278 | 0 | Rhodomyrtus, Vitex trifolia |
| 11 | 19.70 | 109.98 | 94 | 88 | 10 | Rhodomyrtus tomentosa |
| 12 | 19.83 | 109.80 | 108 | 80 | 0 | Rhodomyrtus tomentosa |
| 13 | 19.90 | 109.52 | 2 | 279 | 0 | Rhodomyrtus |
| Variable | ln (Species Richness) | Exponential of Shannon | Inverse of Simpson | Pielou Evenness Index |
|---|---|---|---|---|
| MAT/°C | −0.336 ns | −0.029 ns | 0.049 ns | 0.055 ns |
| MTWQ/°C | −0.310 ns | 0.045 ns | 0.139 ns | 0.149 ns |
| MTCQ/°C | −0.286 ns | −0.116 ns | −0.071 ns | −0.093 ns |
| AVMAT/°C | 0.140 ns | 0.305 ns | 0.326 ns | 0.290 ns |
| MAP/mm | 0.729 ** | 0.225 ns | 0.075 ns | −0.321 ns |
| PDQ/mm | 0.745 ** | 0.149 ns | −0.025 ns | −0.335 ns |
| PWQ/mm | 0.678 * | 0.211 ns | 0.075 ns | −0.350 ns |
| Pcov | −0.701 ** | −0.122 ns | 0.044 ns | 0.356 ns |
| Elevation | 0.125 ns | −0.021 ns | −0.069 ns | −0.243 ns |
| Aspect | −0.254 ns | −0.335 ns | −0.314 ns | −0.149 ns |
| Slope | 0.013 ns | −0.235 ns | −0.288 ns | −0.355 ns |
| SOC | −0.067 ns | −0.486 ns | −0.491 ns | −0.595 * |
| TN | −0.349 ns | 0.295 ns | 0.434 ns | 0.561 * |
| TP | −0.089 ns | −0.411 ns | −0.409 ns | −0.478 ns |
| C/N | 0.294 ns | −0.173 ns | −0.274 ns | −0.309 ns |
| N/P | −0.247 ns | 0.406 ns | 0.523 ns | 0.642 * |
| C/P | 0.062 ns | 0.239 ns | 0.245 ns | 0.240 ns |
| pH value | −0.043 ns | 0.137 ns | 0.170 ns | 0.283 ns |
| Factor | Correlation with RDA Ordination Axes | Monte Carlo Permutation Test | ||
|---|---|---|---|---|
| Axis1 | Axis2 | R2 | p-Value | |
| AVMAT/°C | 0.053 | −0.001 | 0.103 | 0.576 |
| MAP/mm | 0.325 | 0.107 | 0.588 | 0.021 * |
| Aspect | 0.112 | −0.113 | 0.346 | 0.114 |
| SOC | −0.175 | −0.194 | 0.265 | 0.215 |
| TN | 0.276 | −0.229 | 0.531 | 0.025 * |
| TP | 0.011 | 0.201 | 0.180 | 0.376 |
| pH value | 0.000 | 0.141 | 0.049 | 0.779 |
| DCA | 0.476 | 0.208 | ||
| RDA | 5.266 | 0.171 | ||
| Percentage explainable of species data | 62.6 | 2.0 | ||
| Cumulative proportion of species–environment relationships | 62.6 | 64.6 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Wei, S.; Mwangi, B.N.; Liu, S.; Huang, J.; Li, Y. Horizontal Distribution Characteristics and Environmental Factors of Shrubland Species Diversity in Hainan Island, China. Land 2022, 11, 1047. https://doi.org/10.3390/land11071047
Yang M, Wei S, Mwangi BN, Liu S, Huang J, Li Y. Horizontal Distribution Characteristics and Environmental Factors of Shrubland Species Diversity in Hainan Island, China. Land. 2022; 11(7):1047. https://doi.org/10.3390/land11071047
Chicago/Turabian StyleYang, Mengmeng, Simin Wei, Brian N. Mwangi, Shizhong Liu, Jianqiang Huang, and Yuelin Li. 2022. "Horizontal Distribution Characteristics and Environmental Factors of Shrubland Species Diversity in Hainan Island, China" Land 11, no. 7: 1047. https://doi.org/10.3390/land11071047