1. Introduction
Increased tree biodiversity due to land use may seem beneficial, but diversity gains typically occur through the introduction of non-native species or encroachment by native species that were historically rare, with co-occurring losses by historically abundant species and unique ecosystems. Grasslands are one ecosystem for which greater tree diversity and density are a problem for biodiversity. Another example is open forests of savannas and woodlands, where an herbaceous layer coexists with overstory trees. Open forests impart a continuum of conditions and structure across landscapes, which is not provided by uniformly structured successional closed forests with greater tree diversity and density [
1]. Open forests have a limited tree presence, but stand diversity occurs in the herbaceous plants and associated wildlife, which are declining in forests with increased tree diversity and density [
1].
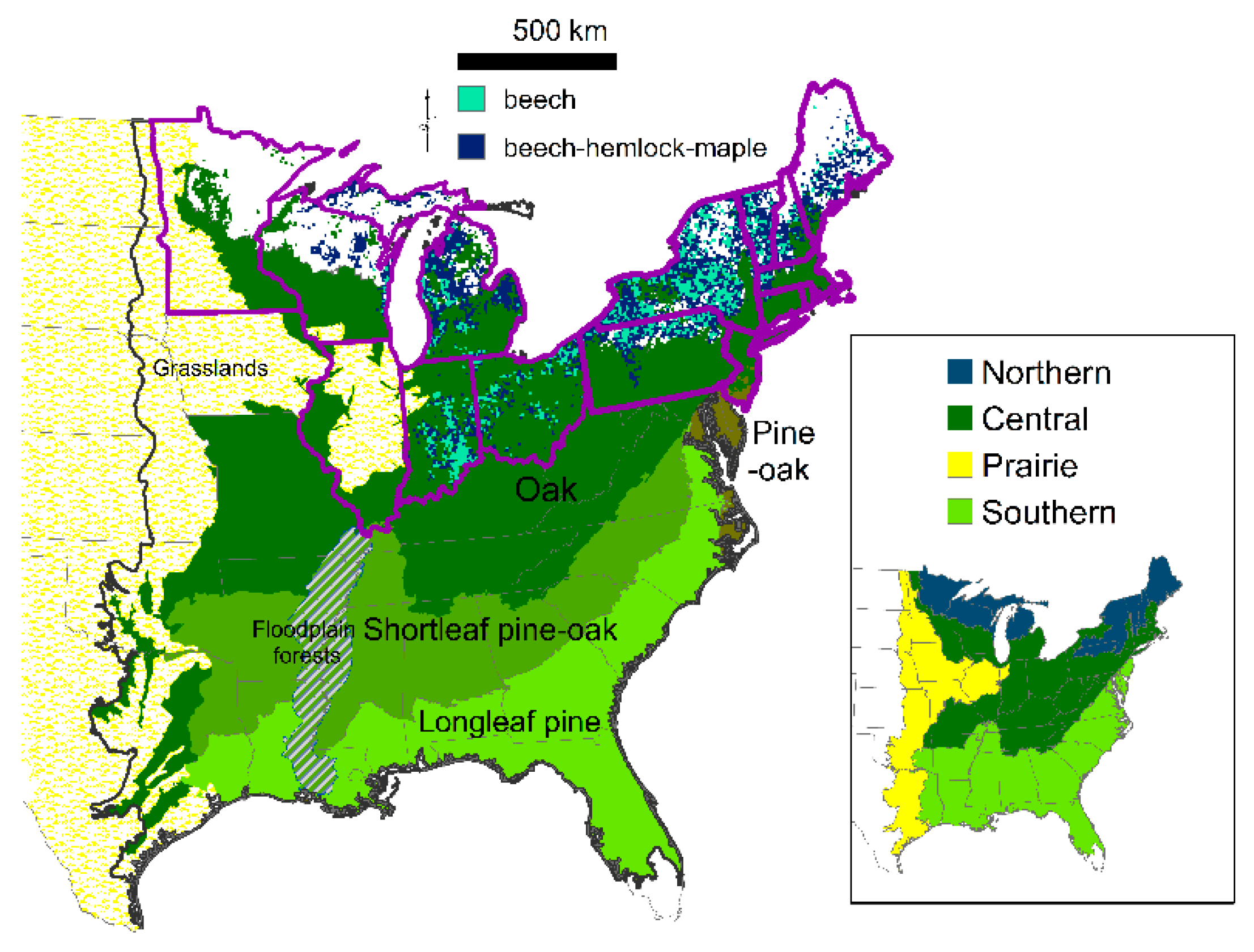
Historically, in the eastern United States, a few foundational tree species represented unique forests with characteristics that supported a distinctive suite of species (
Figure 1). Despite the presence of hundreds of tree species, tree species dominance was a manifestation of infrequent overstory disturbance, combined in many locations with frequent low-severity fires, which reduced tree diversity and density, and maintained growing space for herbaceous plants. For example, open forests of fire-tolerant longleaf pine historically covered the southeastern Coastal Plain, and current remnant stands contain 1630 endemic plants, representing a temperate diversity hotspot [
2]. Open forests of fire-tolerant oaks dominated the central region, with localized forests of shade-tolerant American beech (
Fagus grandifolia, about 5% of all trees), sometimes in conjunction with eastern hemlock (
Tsuga canadensis) or sugar maple (
Acer saccharum), which have similar traits of shade tolerance and a long lifespan. Beech forests were disturbance independent (i.e., old-growth with rare overstory and understory disturbances), but open oak and pine forests were also old-growth forests with long-lived species due to infrequent overstory disturbance [
3]. Although the historical surveys may suffer from measurement bias, for example, a preference for American beech that may be over-representative in records, open oak and pine forest types maintained by low-severity surface fires, modulated by native human management, consistently emerge from the work of countless surveyors and are upheld by pollen evidence [
4] and historical accounts that remain similar, regardless of location or date [
5,
6].
Land-use change with Euro-American settlement incorporated the clearance of historical vegetation, chronic severe disturbance regimes, and surface fire exclusion. Extensive clearcutting during Euro-American settlement was followed by the current successional cycles of overstory tree removal; naturally regenerated forests have a mean age of 40 to 80 years, while pine plantations in the southern U.S. are typically harvested on a 25-year cycle [
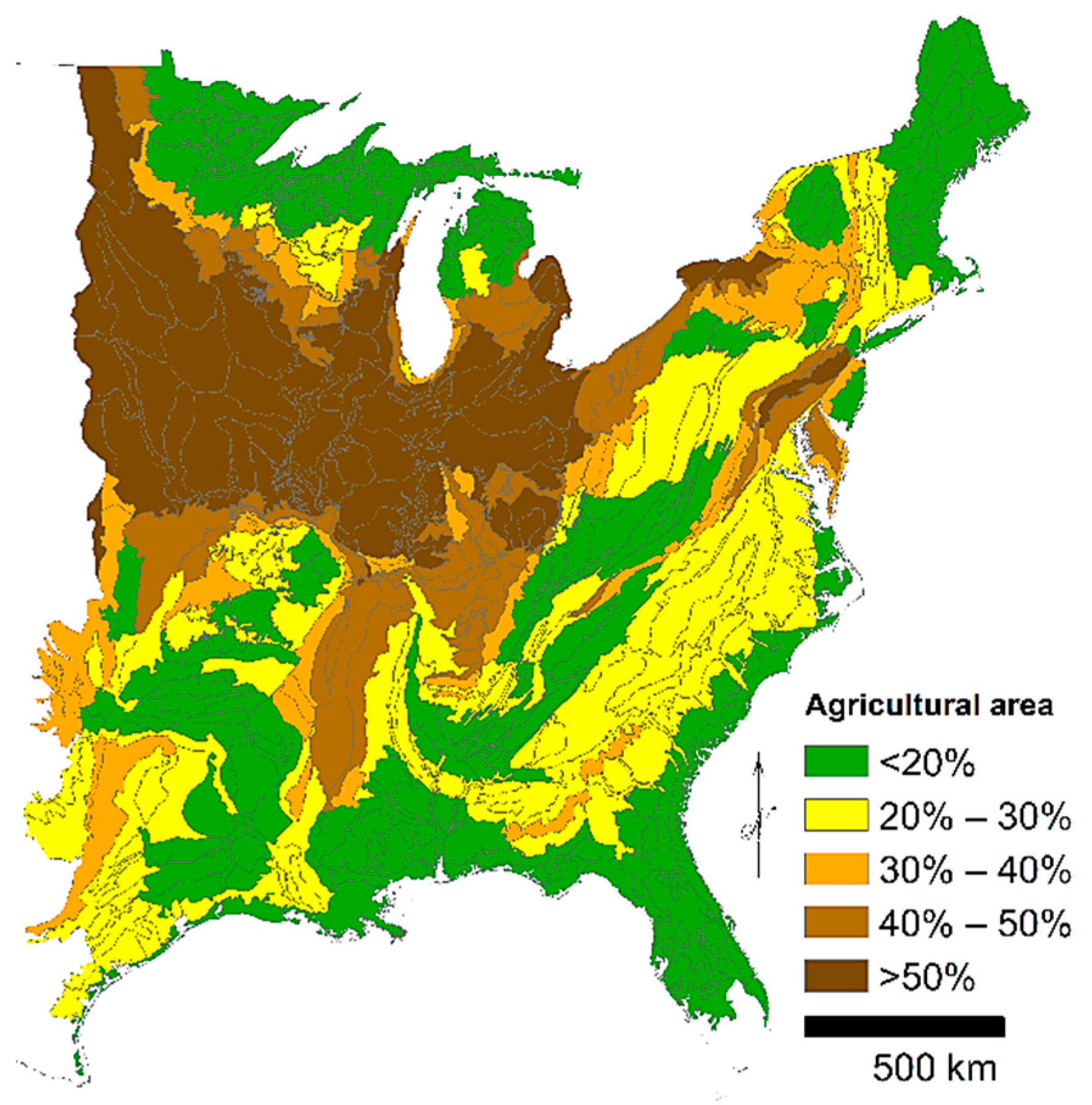
7]. Agricultural conversion was concentrated in the productive warm and flat locations of open oak and pine forests, beech forests, and grasslands rather than the cold, rugged northern region (
Figure 1). Surface fire exclusion occurred through the refutation of fire as a management tool and active fire suppression and additionally land cover changes of roads and agricultural fields and the development of dense, closed forests prevented frequent surface fire spread. Land use change has released a variety of early- to mid-successional tree species to increase in density and expand in range, creating abundant closed successional forests in replacement of the historical forests of either fire-tolerant oaks and pines or late-successional species [
1,
8].
Greater loss in unique ecosystems overall probably occurred with a greater agricultural area, which is a proxy for land use and disturbance changes, resulting in ecosystem replacement by opportunistic native tree species that were uncommon under historical disturbances. Datasets are available to examine the response of tree species to agriculture in the eastern United States, although tree diversity indices may be misinterpreted as a desirable outcome in regions where tree diversity increases in conjunction with agricultural use, which signifies a loss of historical ecosystems [
9]. That is, increased biodiversity typically occurs at the expense of formerly dominant species via increases in the dominance of non-native and formerly non-dominant native species. Instead of using diversity indices, I first tested correspondence with the agricultural area of historical and current tree genera (i.e., measured as a percentage of all trees) and their change over time, through ecological dissimilarity (i.e., Bray–Curtis) and differences between historical and current percentages of each genus, across the northeastern and part of the central United States (
Figure 1). Based on that association, I then comprehensively modeled the relationship between the current percentages of tree species and agricultural area for the eastern United States and regionally to isolate species that may have benefitted from land use change.
2. Materials and Methods
Mean percent area in agriculture from agricultural censuses by county during 1850 to 1997 was used as an index of agricultural area, a proxy for land use change [
10]. Definition of farmland is very broad and includes both cropland and pastureland; areas with cold climates, steep slopes, and unproductive soils were either never or briefly farmed. Mean percent agricultural area was determined by ecological subsections, which are ecological spatial units classified by climate, soils, topography, and vegetation (
Figure 2) [
11]. Ecological subsections are likely to share a common land use history compared to county units based on administrative boundaries.
Percent composition of historical tree genera (tree diameters ≥12.7 cm in diameter at 1.37 m height) was extracted for 231 ecological subsections, the area where similarly modeled data for historical (circa 1620 to 1900) genera were available (
Figure 1) [
12]. Due to species name ambiguity in historical tree surveys, a few genera were combined (i.e., “cedar” could be
Thuja or
Juniperus; “poplar” could be
Populus or
Liriodendron; gum could be
Nyssa or
Liquidambar; ironwood could be
Carpinus or
Ostrya). Unknown species were categorized as “other”; species outside of the genera classes are currently about 2.5% of all species, representing about 30 genera and 50 species. Following the same genera groupings for percent composition of current tree surveys (circa 2010 to 2020; tree diameters ≥12.7 cm in diameter at 1.37 m height) [
13], I quantified the Bray–Curtis dissimilarity between historical and current composition of ecological subsection pairs and applied nonmetric multidimensional scaling ordination of the dissimilarities. Although current tree surveys occurred during the years 2010 to 2020, most recorded naturally regenerated trees have a mean age of 40 to 80 years [
7] and, therefore, reflect new land use conditions of agricultural use surveys conducted during the years 1850 to 1997.
Instead of statistical, parametric models, machine learning regressors were applied to model relationships with greater accuracy. Because agricultural use extends beyond the limits of any one tree species, and also affects more than one tree species, identifying the strength of associations with land use involves modeling multiple variables; that is, correlational models are about strength of relationship and not causation. To relate agricultural area to historical and current genera percentages, their Bray–Curtis dissimilarity, and the difference between historical and current percentage of each genus, regression models with cubist and random forests incorporated 10-fold cross-validation and prediction for of 25% data that were witheld [
14,
15]. I iteratively modeled based on importance of variables until identifying the critical variables.
To establish current relationships between tree genera or species and agricultural area, the same modeling process with cubist and random forest regressors was employed without location restrictions. For 460 ecological subsections of the eastern United States (
Figure 1), I modeled current percentages of 43 genera and one “other” group comprised of 40 rare genera; the other group averaged 1.2% of all trees by subsection. To isolate the species most associated with agricultural area, I modeled current percentages of all but rare tree species of the eastern United States and regionally (
Figure 1). Due to fewer ecological subsection samples in the prairie region, the prairie and central regions were combined.
3. Results
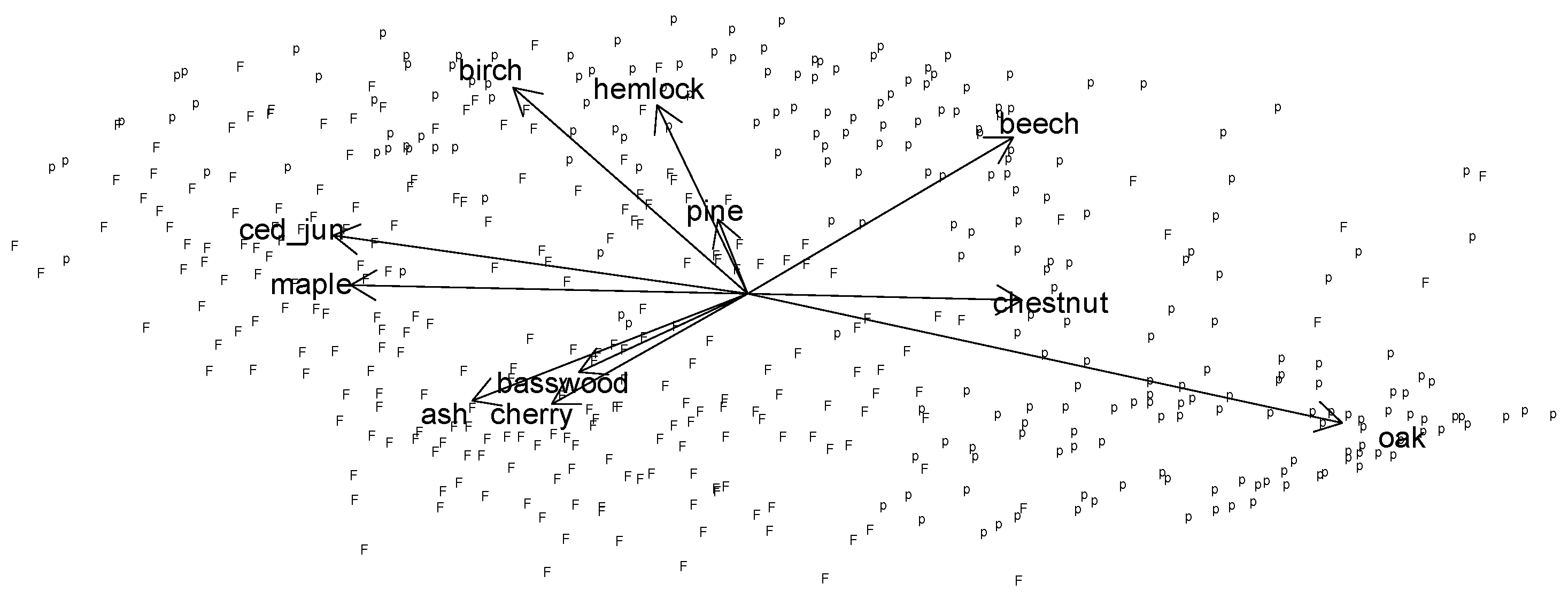
The magnitude of disturbance and forest change in landscape diversity was expressed by a major reversal from fire-dependent oak and pine forests or disturbance-independent beech forests to diverse eastern broadleaf forests of maple (i.e., red maple;
A. rubrum), ashes, and cherries (
Figure 3). Nonetheless, according to the regressors, neither the Bray–Curtis dissimilarity between the historical and current percentages of each genus nor the difference between the historical and current percentage of each genus was strongly related to agricultural area. Likewise, dominance by the historical genera was not influential. Instead, the current percentages of elm and the uncommon “other” genera were the model variables with the strongest relationship with agricultural area; the
R2 value was 78% (both regressors) for the withheld samples. The
R2 was 61% and 68% with only one of the two variables, and 21% and 31% with Bray–Curtis dissimilarity as the predictor.
For the relationship between the current genera and agricultural area in the eastern United States,
Ulmus,
Juglans, and
Prunus best predicted agricultural area (
R2 of 66% for both regressors;
Figure 4). Modeling of the six most influential genera, with the inclusion of
Acer,
Carya, and
Fraxinus, slightly increased the
R2 values to 70% (cubist regressor) and 75% (random forest regressor), which was close to the maximum
R2 value of about 78% for 17 genera.
Ulmus alone had
R2 values of 33% and 38%.
For the eastern United States, the current percentage of boxelder, black cherry, and hackberry, and the total percentage of these species were the most related to agricultural area (
R2 of 66% and 68%;
Figure 4). Because the greatest agricultural area occurred in the central region extending into the prairie region, boxelder, black cherry, and hackberry were the most influential predictors, along with green ash (
Fraxinus pennsylvanica), black walnut, and American elm (
R2 of 57% and 68% for the central region, and
R2 of 62% and 68% for the central and prairie regions). For the southern region, slippery elm (
Ulmus rubra), American sycamore, red maple, and sweetgum best predicted agricultural area (
R2 of 60% and 62%). For the northern region, green ash, white ash (
Fraxinus americana), and mockernut hickory had the strongest relationship with agricultural area (
R2 of 74%).
4. Discussion
Due to land use changes, diverse eastern broadleaf species have increased and expanded, replacing historically dominant oak and pine forests throughout the eastern United States since Euro-American settlement. Elms, walnuts, and cherries as genera, and slippery elm, American elm, red maple and boxelder (
Acer negundo), green and white ashes, black cherry, hackberry, black walnut, mockernut hickory, sweetgum, and American sycamore in particular, were associated with agricultural area, a proxy for a cascade of land use effects on ecosystems. This study extended the work of Vellend et al. [
9], with similar findings that a diverse group of historically rare species aligned with agricultural area remarkably well, considering the spatial extent, unique species distributions, and complicated land use histories.
Of the species with the strongest relationships with agriculture, most have increased and expanded over time, notably, red maple, boxelder, and black cherry [
8,
16]. Even though specific dynamics of mockernut hickory are unknown, hickories as a genus have likely remained stable in relative abundance because hickories historically had some representation in uplands exposed to fire. Hickories and walnuts received assistance and protection, as these genera were agricultural crops for native humans [
17,
18]. Black walnut remains an agricultural crop, and mockernut hickory distribution may endure along historical agricultural pathways, perhaps concentrating over time [
19]. American elm has decreased due to Dutch elm disease (Ophiostomataceae), but American elm historically occurred along riparian networks, an ideal match for agriculture.
Indeed, all of the identified species have some affinity for lowlands (
Table 1) [
20], which may be an artifact of filtering by historical frequent surface fire regimes. Fire-sensitive species survived in protected firebreaks, which were the moisture extremes of wetlands and rocky outcrops with discontinuous vegetation. However, some herbaceous wetlands burn frequently in the eastern United States, and relatively frequent fires in less productive sites may produce equivalent tree control to frequent fires in sites where trees grow rapidly. In any event, diverse fire-sensitive eastern broadleaf and pine species navigated the space between upland frequent fires and lowland frequent flooding [
16,
21]. Therefore, although these species were native to eastern forests, they tended to be spatially partitioned rather than uniformly distributed across landscapes.
The tree species with the strongest relationships with agriculture expressed an exchange in competitive balance among species with different disturbance response traits. Most of the identified species are relatively short-lived, with longevity as a trade for the traits of fast growth during early life stages to reach abundant seed dispersal [
20]. Rapid establishment and reproduction after historically rare severe overstory disturbances, such as flooding in the past, make these species suited to recovery from current disturbance regimes. For example, even though the two maple species are moderately shade tolerant, which means that they are considered mid-successional, early growth is most rapid until seed production develops; furthermore, growth and reproduction are prioritized over survivorship, as they seldom live more than 100 to 150 years [
22,
23]. Therefore, the maple species have the traits of successional colonizing species under unstable conditions, which are generated by human activities, unlike fire-tolerant or shade-tolerant competitors that devote resources to survival under stable conditions [
8].
Land use change encompasses multiple components. Tree response may be proportionate to the land use directly in agriculture (e.g., walnuts), disturbance change indicated by agricultural area, such as clearcutting followed by frequent overstory tree removal [
7], and the disruption of frequent surface fires. Species have expanded both by colonization and deliberate introduction through planting in field borders, old fields after agricultural abandonment, and grasslands converted to agriculture, even in recent decades [
24,
25]. However, other species, such as slippery elm, may have increased particularly in forests [
26,
27,
28]. Species with traits for rapid response after disturbance established in second-growth forests that developed after clearing for forest products or agriculture with subsequent agricultural abandonment; relatively frequent overstory disturbance has continued along with surface fire exclusion [
7]. In the historical grasslands of the eastern United States, numerous historical accounts related the visible transition from surface fires, which controlled tree establishment, to fire exclusion as settlements, agricultural fields, and trees developed [
29,
30]. A dimension of fire exclusion is that land use change allows the development of trees, which also prevent fire spread by reducing or removing the herbaceous vegetation that provides fine fuels.
Current forests comprised of diverse eastern broadleaf species, such as those favored by agriculture, are characteristically dense, closed forests with small-diameter trees relative to historical forests [
1]. Without control by surface fire or large-diameter trees that capture the surrounding resources, small-diameter trees are free to establish and grow. Eventually, self-thinning does occur as trees of similar sizes compete for resources, but these trees are cleared within decades, before characteristics of disturbance-independent closed forests develop [
7]. Historical forests were old-growth closed and open forests, but not successional forests (
Figure 1). In addition to composition and structure, successional forests have different functions, conditions, and biodiversity than historical forests [
3,
31]. Indeed, some “successional” species are declining despite the presence of abundant successional forests, indicating the misattribution.
The application of invasive species terminology to native species is contentious. Analogous terms, such as encroachment, are instead applied when native species claim the growing space of historically abundant species due to the direct removal of established species by land use and disruption of the competitive balance after the transformed disturbance to more frequent overstory removal and less frequent understory removal. Logically, if invasive non-native species are specified, then invasive native species can also be specified as invasive. Invasive species tend to have high dispersal rates and rapid growth rates with short generation times, allowing tolerance for rapid changes, similar to the traits of the increasing native tree species. The increase in native tree populations has not only had a negative effect, but, moreover, it has replaced historical open forest and grassland ecosystems with eastern broadleaf forests. Indeed, the magnitude of invasion by native tree species is a regime shift to an alternative state away from fire-tolerant forests. Dense forests intrinsically prevent the return of frequent surface fires and re-establishment of fire-tolerant forests through replacement of herbaceous plants, which act as fine fuels for fire spread, with trees, which burn rarely but severely, as fire travels vertically up smaller trees to the canopies. Furthermore, closed forest conditions, including wind velocities, are less suitable for ignition and spread. This internal mechanism of dense, successional forests acting as a surface fire break reinforces active fire exclusion and land use changes, such as agriculture, which prevent surface fires.
Although the outcome of ecosystem alteration is similar, invasive native and non-native plant species may retain differences in their direct ability to supplant vegetation in the absence of disturbance change. Rather than driving change, native tree species facilitated disturbance, by either promoting open or closed conditions. Invasive native tree species are a symptom of land use change, controlled and spatially isolated before disturbance change, and uncontrolled and integrated after disturbance change, from infrequent severe disturbance with frequent surface fires to frequent severe disturbance with infrequent surface fires. If there were not so many native tree species in the eastern United States, historically dominant tree species may not regenerate well enough under severe disturbance to out-compete herbaceous vegetation in some locations; this could modify human land use to reduce successional cycles in an effort to promote reforestation. Additionally, interpretations of reconstructions in a few localized areas were that historical vegetation may operate independently of disturbance, albeit the surrounding vegetation matrix, which provides propagules, may impose conformity on stands in the absence of disturbance [
32]. Conversely, some invasive species may potentially have succeeded under historical conditions and directly influenced a change in conditions. Certainly, numerous invasive species were released in the eastern United States during the 1800s and earlier, but a base level of propagule pressure by continued introductions is required before the species can become invasive [
33]. Regardless, native and introduced plant species have assembled into novel communities in response to the human activities of devegetation and agricultural use cycles, which are unfavorable to the regeneration of historical ecosystems in the eastern United States and globally [
34].








{kind=link}
{kind=link}
{kind=link}
{kind=link}