
Hillslope Geodiversity Impact on Biocrusts’ Biogeochemical Functions

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
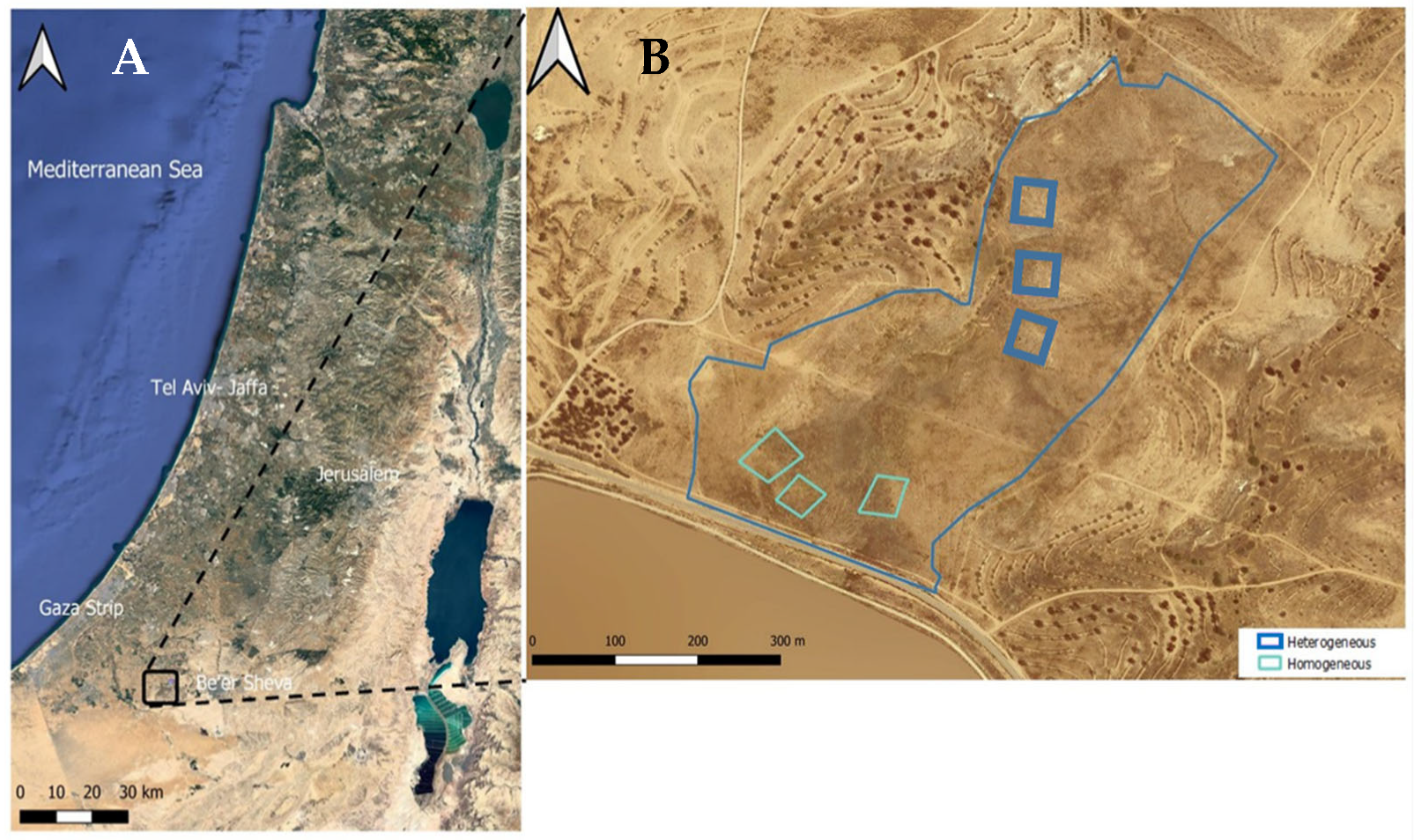
2.1. The Study Sites
2.2. Field and Laboratory Work
2.3. Statistical Analysis
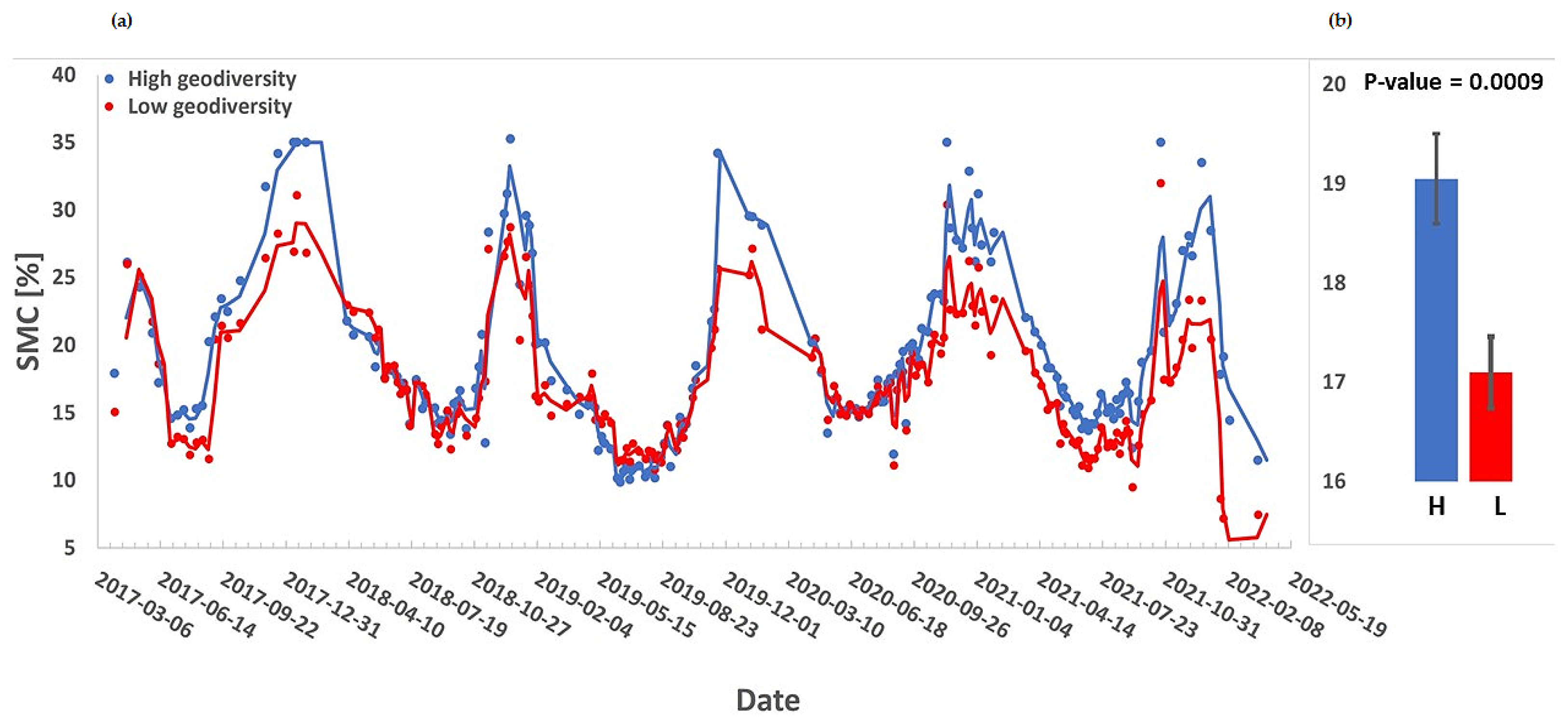
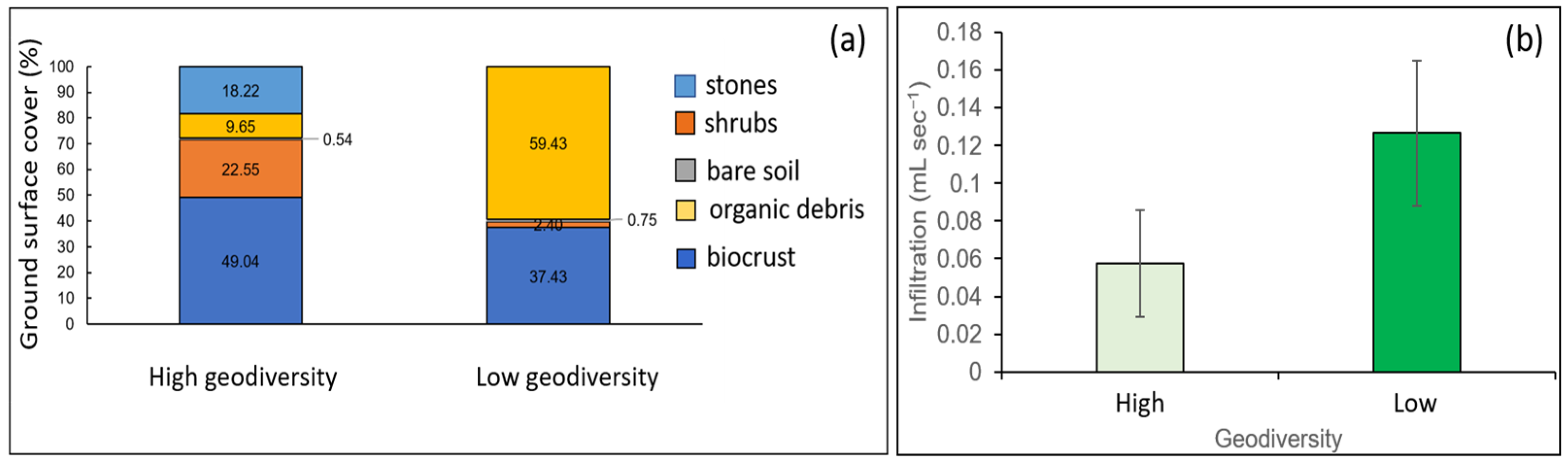
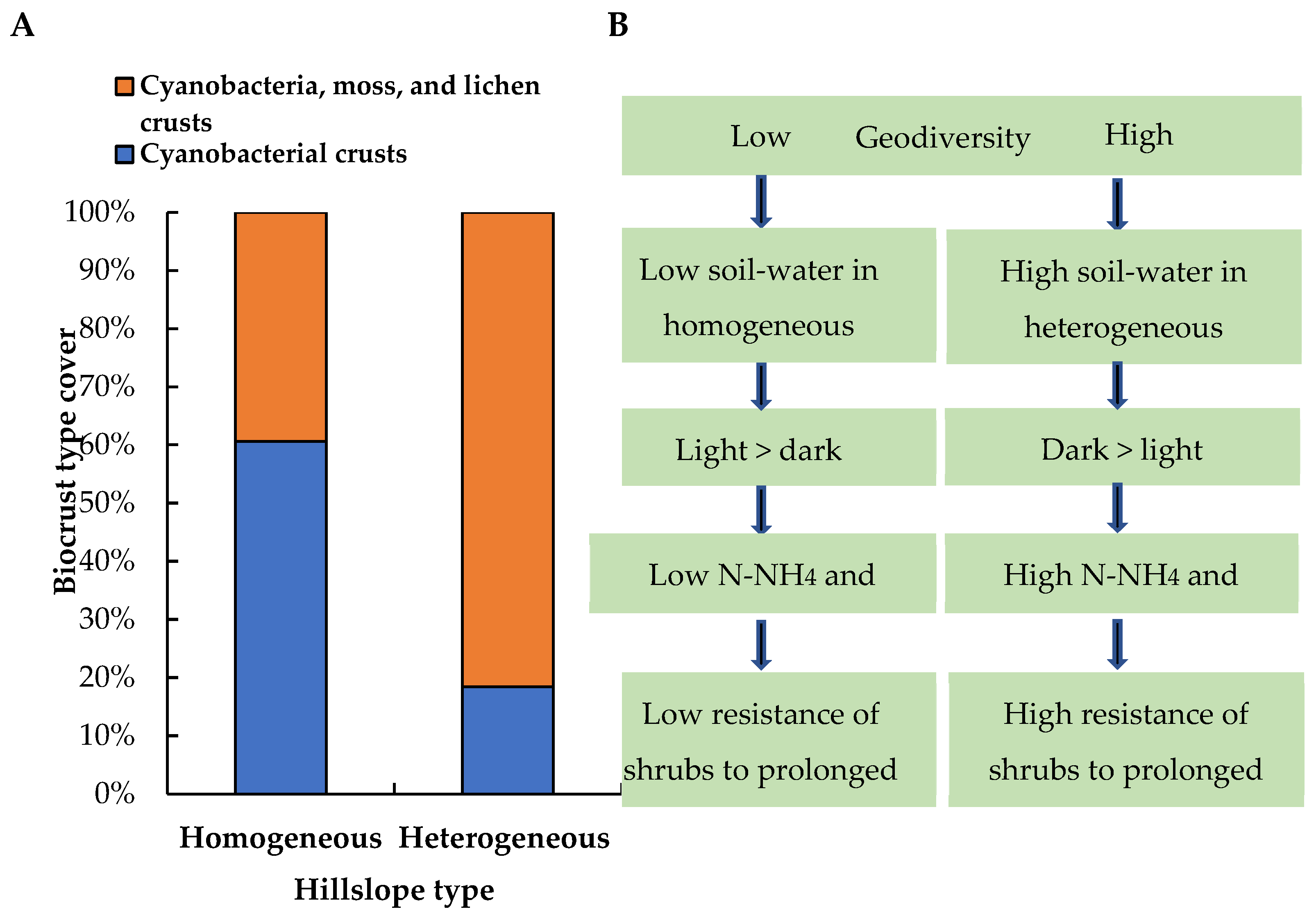
3. Results
4. Discussion



5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gray, M. Geodiversity and geo-conservation: What, why, and how? In Geodiversity and Geoconservation; Santucci, V.L., Ed.; The George Wright Forum: Hancock, MI, USA, 2005; Volume 22, pp. 4–12. Available online: http://www.georgewright.org/223.pdf (accessed on 1 January 2021).
- Bétard, F. Patch-scale relationships between geodiversity and biodiversity in hard rock quarries: A case study from a disused quartzite quarry in NW France. Geoheritage 2013, 5, 59–71. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.G.; Comer, P.J.; Beier, P.; Lawler, J.J.; Schloss, C.A.; Buttrick, S.; Albano, C.M.; Faith, D.P. Case studies of conservation plans that incorporate geodiversity. Conserv. Biol. 2015, 29, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Hjort, J.; Gordon, J.E.; Gray, M.; Hunter, M.L., Jr. Why geodiversity matters in valuing nature’s stage? Conserv. Biol. 2015, 29, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Tukiainen, H.; Bailey, J.J.; Field, R.; Kangas, K.; Hjort, J. Combining geodiversity with climate and topography to account for threatened species richness. Conserv. Biol. 2017, 31, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Stavi, I.; Rachmilevitch, S.; Hjazin, A.; Yizhaq, H. Geodiversity decreases shrub mortality and increases ecosystem tolerance to droughts and climate change. Earth Surf. Process. Landf. 2018, 43, 2808–2817. [Google Scholar] [CrossRef]
- De Falco, N.; Tal-Berger, R.; Hjazin, A.; Yizhaq, H.; Stavi, I.; Rachmilevitch, S. Geodiversity impacts plant community structure in a semi-arid region. Sci. Rep. 2021, 11, 15259. [Google Scholar] [CrossRef] [PubMed]
- Comer, P.J.; Pressey, R.L.; Hunter, M.L.; Schloss, C.A.; Buttrick, S.C.; Heller, N.E.; Tirpak, J.M.; Faith, D.P.; Cross, M.S.; Shaffer, M.L. Incorporating geodiversity into conservation decisions. Conserv. Biol. 2015, 29, 692–701. [Google Scholar] [CrossRef]
- Yizhaq, H.; Stavi, I.; Shachak, M.; Bel, G. Geodiversity increases ecosystem durability to prolonged droughts. Ecol. Complex. 2017, 31, 96–103. [Google Scholar] [CrossRef]
- Stavi, I.; Rachmilevitch, S.; Yizhaq, H. Geodiversity effects on soil quality and geo-ecosystem functioning in drylands. Catena 2019, 176, 372–380. [Google Scholar] [CrossRef]
- Evans, R.D.; Lange, O.L. Biological soil crusts and ecosystem nitrogen and carbon dynamics. In Biological Soil Crusts: Structure, Function, and Management, Ecological Studies; Belnap, J., Lange, O.L., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 263–279. [Google Scholar] [CrossRef]
- Whitford, W.; Duval, B. Ecology of Desert Systems, 2nd ed.; Academic Press: New York, NY, USA, 2002; pp. 259–260. [Google Scholar]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Budel, B.; Andreae, M.O.; Pöschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Sancho, L.G.; Belnap, J.; Colesie, C.; Raggio, J.; Weber, B. Carbon budgets of biological soil crusts at micro-, meso-, and global scales. In Biological Soil Crusts: An Organizing Principle in Drylands, Ecological Studies; Weber, B., Büdel, B., Belnap, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 226, pp. 287–304. [Google Scholar] [CrossRef]
- Belnap, J. Nitrogen fixation in biological soil crusts from southeast Utah, USA. Biol. Fertil. Soils 2002, 35, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Weber, B.; Wu, D.; Tamm, A.; Ruckteschler, N.; Rodríguez-Caballero, E.; Steinkamp, J.; Meusel, H.; Elbert, W.; Behrendt, T.; Sörgel, M.; et al. Biological soil crusts emit NO and HONO. Proc. Natl. Acad. Sci. USA 2015, 112, 15384–15389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barger, N.N.; Belnap, J.; Garcia-Pichel, F.; Zaady, E. Nitrogen cycling of biological soil crusts at micro-macro- and global scales. In Biological Soil Crusts: An Organizing Principle in Drylands. Ecological Studies; Weber, B., Büdel, B., Belnap, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 226, pp. 257–285. [Google Scholar]
- Fernandes, V.M.C.; Rudgers, J.A.; Collins, S.L.; Garcia-Pichel, F. Rainfall pulse regime drives biomass and community composition in biological soil crusts. Ecology 2022, 103, e3744. [Google Scholar] [CrossRef]
- Pointing, S.; Belnap, J. Microbial colonization and controls in dryland systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef]
- Rodríguez-Caballero, E.; Cantón, Y.; Chamizo, S.; Afana, A.; Solé-Benet, A. Effects of biological soil crusts on surface roughness and implications for runoff and erosion. Geomorphology 2012, 145, 81–89. [Google Scholar] [CrossRef]
- Chamizo, S.; Belnap, J.; Eldridge, D.J.; Cantón, Y.; Malam-Issa, O. The role of biocrusts in arid land hydrology. In Biological Soil Crusts: An Organizing Principle in Drylands. Ecological Studies; Weber, B., Büdel, B., Belnap, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 226, pp. 321–346. [Google Scholar] [CrossRef]
- Zaady, E.; Stavi, I.; Yizhaq, H. Hillslope geodiversity in drylands: Effects on composition and properties of biological soil crusts. Eur. J. Soil Sci. 2021, 72, 2308–2315. [Google Scholar] [CrossRef]
- Israel Meteorological Service. 2021. Available online: http://www.ims.gov.il/en (accessed on 1 January 2021).
- Zaady, E.; Arbel, S.; Barkai, D.; Sarig, S. Long-term impact of agricultural practices on biological soil crusts and their hydrological processes in a semiarid landscape. J. Arid. Environ. 2013, 90, 5–11. [Google Scholar] [CrossRef]
- Shachak, M.; Sachs, M.; Moshe, I. Ecosystem management of desertified shrublands in Israel. Ecosystems 1998, 1, 475–483. [Google Scholar] [CrossRef]
- Sher, Y.; Zaady, E.; Nejidat, A. Spatial and temporal diversity and abundance of ammonia oxidizers in semiarid and arid ecosystems: Indications for a differential seasonal effect on archaeal and bacterial ammonia oxidizers. FEMS Microbiol. Ecol. 2013, 86, 544–556. [Google Scholar] [CrossRef]
- Zaady, E.; Ben-David, E.A.; Sher, Y.; Tzirkin, R.; Nejidat, A. Inferring biological soil crust successional stage using combined PLFA, DGGE, physical and biophysiological analyses. Soil Biol. Biochem. 2010, 42, 842–849. [Google Scholar] [CrossRef]
- Nejidat, A.; Potrafka, R.M.; Zaady, E. Successional biocrust stages on dead shrub soil mounds after severe drought: Effect of micro-geomorphology on microbial community structure and ecosystem recovery. Soil Biol. Biochem. 2016, 103, 213–220. [Google Scholar] [CrossRef]
- Sadeghi, M.; Jones, S.B.; Philpot, W.D. A linear physically-based model for remote sensing of soil moisture using short wave infrared bands. Remote Sens. Environ. 2015, 164, 66–76. [Google Scholar] [CrossRef]
- Dubinin, V.; Svoray, T.; Stavi, I.; Yizhaq, H. Using LANDSAT 8 and VENµS data to study the effect of geodiversity on soil moisture dynamics in a semiarid shrubland. Remote Sens. 2020, 12, 3377. [Google Scholar] [CrossRef]
- Clesceri, L.S.; Greenberg, A.E.; Trussell, R.R. Standard Methods for the Examination of Water and Wastewater, 17th ed.; American Public Health Association: Washington, DC, USA, 1989; p. 164. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association/American Water Works Association/Water Environment Federation: Washington, DC, USA, 2005. [Google Scholar]
- Ben-Dor, E.; Banin, A. Determination of organic matter content in arid-zone soils using simple “Loss-on-ignition” method. Commun. Soil Plant Anal. 1989, 20, 1675–1695. [Google Scholar] [CrossRef]
- Tirol-Padre, A.; Ladha, J.K. Assessing the reliability of permanganate-oxidizable carbon as an index for soil labile carbon. Soil Sci. Plant Anal. 2004, 67, 969–978. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of Protein. Methods Enzymol. 1990, 182, 50–68. [Google Scholar] [CrossRef]
- Faist, A.M.; Herrick, J.E.; Belnap, J.; Van Zee, J.W.; Barger, N.N. Biological soil crust and disturbance controls on surface hydrology in a semiarid ecosystem. Ecosphere 2017, 8, e01691. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS User’s Guide: Statistics. 2016, Cary, NC. Available online: https://support.sas.com/documentation/onlinedoc/stat/142/stathpug.pdf (accessed on 1 November 2016).
- Garcia-Pichel, F.; Pringault, O. Cyanobacteria track water in desert soils. Nature 2001, 413, 380–381. [Google Scholar] [CrossRef]
- Zaady, E.; Katra, I.; Yizhaq, H.; Kinast, S.; Ashkenazy, Y. Inferring the impact of rainfall gradient on biocrusts’ developmental stage and thus on soil physical structures in sand dunes. Aeolian Res. 2014, 13, 81–89. [Google Scholar] [CrossRef]
- Bowker, M.A.; Belnap, J.; Budel, B.; Sannier, C.; Pietrasiak, N.; Eldridge, D.J.; Rivera-Aguilar, V. Control on distribution patterns of biological soil crusts at micro-to global scales. In Biological Soil Crusts: An Organizing Principle in Drylands. Ecological Studies; Weber, B., Budel, B., Belnap, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 226, Chapter 10; pp. 173–198. [Google Scholar] [CrossRef]
- Zaady, E.; Kuhn, U.; Wilske, B.; Sandoval-Soto, L.; Kesselmeier, J. Patterns of CO2 exchange in biological soil crusts of successional age. Soil Biol. Biochem. 2000, 32, 959–966. [Google Scholar] [CrossRef]
- Housman, D.C.; Powers, H.H.; Collins, A.D.; Belnap, J. Carbon and nitrogen fixation differ between successional stages of biological soil crustsin the Colorado Plateau and Chihuahuan desert. J. Arid. Environ. 2006, 66, 620–634. [Google Scholar] [CrossRef] [Green Version]
- Dubinin, V.; Stavi, I.; Sovray, T.; Dorman, M.; Yizhaq, H. Hillslope geodiversity improves the resistance of shrubs to prolonged droughts in semiarid ecosystems. J. Arid. Environ. 2021, 188, 104462. [Google Scholar] [CrossRef]
- Zaady, E.; Groffman, P.; Shachak, M. Nitrogen fixation in macro- and microphytic patches in the Negev desert. Soil Biol. Biochem. 1998, 30, 449–454. [Google Scholar] [CrossRef]
- Szitenberg, A.; Alexander-Shani, R.; Yizhak, H.; Stavi, I. Hillslope geodiversity shapes ammonia-oxidizing communities and other microbial regulators in a semiarid shrubland. bioRxiv 2021. [Google Scholar] [CrossRef]
- Couradeau, E.; Giraldo-Silva, A.; De Martini, F.; Garcia-Pichel, F. Spatial segregation of the biological soil crust microbiome around its foundational cyanobacterium, Microcoleus vaginatus, and the formation of a nitrogen-fixing cyanosphere. Microbiome 2019, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Grez, B.; Tam, K.; Cross, A.T.; Yong, J.W.H.; Kumaresan, D.; Nevill, P.; Farrell, M.; Whiteley, A.S. The bacterial microbiome associated with arid biocrusts and the biogeochemical influence of biocrusts upon the underlying soil. Front. Microbiol. 2019, 10, 2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stavi, I.; Yizhaq, H.; Szitenberg, A.; Zaady, E. Patch- to hillslope-scale geodiversity alleviates susceptibility of dryland ecosystems to climatic change: Insights from the Israeli Negev. Curr. Opin. Environ. Sustain. 2021, 50, 129–137. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hillslope Types | SOM (g kg−1) | LOC (mg g−1) | N-NH4 (mg kg soil−1) | N-NO2 (mg kg soil−1) | Protein (mg kg−1 soil−1) |
|---|---|---|---|---|---|
| Heterogeneous | 4.632 (±0.756 *) a | 0.00715 (±0.0030) a | 2.536 (±1.685) a | 0.112 (±0.079) a | 0.0917 (±0.041) a |
| Homogeneous | 3.949 (±0.778) b | 0.00626 (±0.0022) b | 1.659 (±0.882) b | 0.102 (±0.069) a | 0.0912 (±0.034) a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaady, E.; Stavi, I.; Dubinin, V.; Kamennaya, N.; Abu-Glion, H.; Shuker, S.; Yizhaq, H. Hillslope Geodiversity Impact on Biocrusts’ Biogeochemical Functions. Land 2022, 11, 1983. https://doi.org/10.3390/land11111983
Zaady E, Stavi I, Dubinin V, Kamennaya N, Abu-Glion H, Shuker S, Yizhaq H. Hillslope Geodiversity Impact on Biocrusts’ Biogeochemical Functions. Land. 2022; 11(11):1983. https://doi.org/10.3390/land11111983
Chicago/Turabian StyleZaady, Eli, Ilan Stavi, Vladislav Dubinin, Nina Kamennaya, Hiam Abu-Glion, Shimshon Shuker, and Hezi Yizhaq. 2022. "Hillslope Geodiversity Impact on Biocrusts’ Biogeochemical Functions" Land 11, no. 11: 1983. https://doi.org/10.3390/land11111983