
Ecological Corridor Construction Based on Least-Cost Modeling Using Visible Infrared Imaging Radiometer Suite (VIIRS) Nighttime Light Data and Normalized Difference Vegetation Index

Abstract
:1. Introduction
2. Materials and Methods
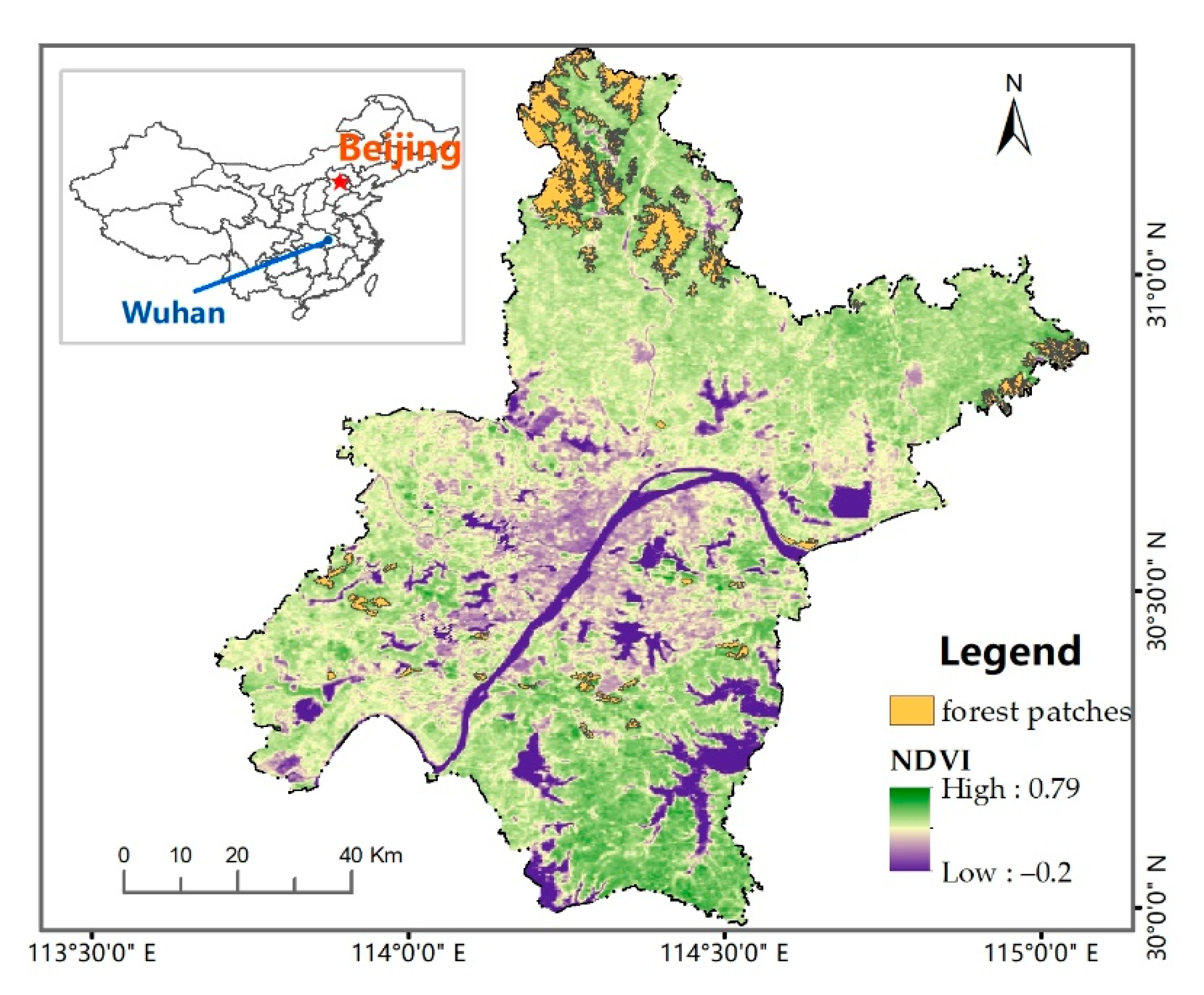
2.1. Study Area
2.2. Data Sources and Processing
2.3. Analysis Methods
2.3.1. Matrix Resistance from the Nighttime Light Adjusted Vegetation Index (NAVI)
2.3.2. Least-Cost Path
2.3.3. Analysis of Nightscape Change and Comparison of Connectivity Results
3. Results
3.1. Analysis of Nightscape Change
3.2. Comparison of Ecological Network Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Butchart, S.H.M. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, M.K.; Matthews, T.J.; Whittaker, R.J. The general dynamic model: Towards a unified theory of island biogeography? Glob. Ecol. Biogeogr. 2016, 25, 805–816. [Google Scholar] [CrossRef]
- Hilty, J.; Worboys, G.L.; Keeley, A.; Woodley, S.; Lausche, B.J.; Locke, H.; Carr, M.; Pulsford, I.; Pittock, J.; White, J.W.; et al. Guidelines for Conserving Connectivity through Ecological Networks and Corridors; Groves, C., Ed.; International Union for Conservation of Nature: Gland, Switzerland, 2020. [Google Scholar] [CrossRef]
- Bunn, A.G.; Urban, D.L.; Keitt, T.H. Landscape connectivity: A conservation application of graph theory. J. Environ. Manag. 2000, 59, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Avon, C.; Bergès, L. Prioritization of habitat patches for landscape connectivity conservation differs between least-cost and resistance distances. Landsc. Ecol. 2016, 31, 1551–1565. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Song, W. Identifying Ecological Corridors and Networks in Mountainous Areas. Int. J. Environ. Res. Public Health 2021, 18, 4797. [Google Scholar] [CrossRef] [PubMed]
- Green, A.L.; Fernandes, L.; Almany, G.; Abesamis, R.; McLeod, E.; Alino, P.M.; White, A.T.; Salm, R.; Tanzer, J.; Pressey, R.L. Designing Marine Reserves for Fisheries Management, Biodiversity Conservation, and Climate Change Adaptation. Coast. Manag. 2014, 42, 143–159. [Google Scholar] [CrossRef]
- Shi, F.N.; Liu, S.L.; An, Y.; Sun, Y.X.; Zhao, S.; Liu, Y.X.; Li, M.Q. Spatio-Temporal Dynamics of Landscape Connectivity and Ecological Network Construction in Long Yangxia Basin at the Upper Yellow River. Land 2020, 9, 265. [Google Scholar] [CrossRef]
- Jongman, R.H.G. Nature Conservation Planning in Europe—Developing Ecological Networks. Landsc. Urban Plan. 1995, 32, 169–183. [Google Scholar] [CrossRef]
- Sorvig, K. Ecological networks and greenways: Concept, design, implementation. Landsc. Archit. 2005, 95, 137. [Google Scholar]
- Tabor, G.; Bankova-Todorova, M.; Correa Ayram, C.; Garcia, L.; Kapos, V.; Olds, A.; Stupariu, I. Ecological Connectivity: A Bridge to Preserving Biodiversity; United Nations Environment Programme: Nairobi, Kenya, 2019; pp. 24–37. [Google Scholar]
- Tischendorf, L.; Fahrig, L. On the usage and measurement of landscape connectivity. Oikos 2000, 90, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, T.; Sharma, L.K.; Kumar, V.; Sharief, A.; Dutta, R.; Kumar, M.; Joshi, B.D.; Thakur, M.; Venkatraman, C.; Chandra, K. Adaptive spatial planning of protected area network for conserving the Himalayan brown bear. ScTEn 2021, 754, 11. [Google Scholar] [CrossRef]
- Allen, C.H.; Parrott, L.; Kyle, C. An individual-based modelling approach to estimate landscape connectivity for bighorn sheep (Ovis canadensis). PeerJ 2016, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Xie, P.; Yang, J.; Wang, H.Y.; Liu, Y.F.; Liu, Y.L. A New method of simulating urban ventilation corridors using circuit theory. Sustain. Cities Soc. 2020, 59, 10. [Google Scholar] [CrossRef]
- Li, H.; Li, D.; Li, T.; Qiao, Q.; Yang, J.; Zhang, H. Application of least-cost path model to identify a giant panda dispersal corridor network after the Wenchuan earthquake—Case study of Wolong Nature Reserve in China. Ecol. Model. 2010, 221, 944–952. [Google Scholar] [CrossRef]
- Ziółkowska, E.; Ostapowicz, K.; Radeloff, V.C.; Kuemmerle, T. Effects of different matrix representations and connectivity measures on habitat network assessments. Landsc. Ecol. 2014, 29, 1551–1570. [Google Scholar] [CrossRef] [Green Version]
- Zeller, K.A.; McGarigal, K.; Whiteley, A.R. Estimating landscape resistance to movement: A review. Landsc. Ecol. 2012, 27, 777–797. [Google Scholar] [CrossRef]
- Adriaensen, F.; Chardon, J.P.; De Blust, G.; Swinnen, E.; Villalba, S.; Gulinck, H.; Matthysen, E. The application of ‘least-cost’ modelling as a functional landscape model. Landsc. Urban Plan. 2003, 64, 233–247. [Google Scholar] [CrossRef]
- Kindlmann, P.; Burel, F. Connectivity measures: A review. Landsc. Ecol. 2008, 23, 879–890. [Google Scholar] [CrossRef] [Green Version]
- Unfried, T.M.; Hauser, L.; Marzluff, J.M. Effects of urbanization on Song Sparrow (Melospiza melodia) population connectivity. Conserv. Genet. 2012, 14, 41–53. [Google Scholar] [CrossRef]
- Gurrutxaga, M.; Rubio, L.; Saura, S. Key connectors in protected forest area networks and the impact of highways: A transnational case study from the Cantabrian Range to the Western Alps (SW Europe). Landsc. Urban Plan. 2011, 101, 310–320. [Google Scholar] [CrossRef]
- Beier, P.; Spencer, W.; Baldwin, R.F.; McRae, B.H. Toward best practices for developing regional connectivity maps. Conserv. Biol. 2011, 25, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Krosby, M.; Breckheimer, I.; Pierce, D.J.; Singleton, P.H.; Hall, S.A.; Halupka, K.C.; Gaines, W.L.; Long, R.A.; McRae, B.H.; Cosentino, B.L.; et al. Focal species and landscape “naturalness” corridor models offer complementary approaches for connectivity conservation planning. Landsc. Ecol. 2015, 30, 2121–2132. [Google Scholar] [CrossRef]
- Baldwin, R.F.; Perkl, R.M.; Trombulak, S.C.; Burwell, W.B. Modeling Ecoregional Connectivity. In Landscape-Scale Conservation Planning; Trombulak, S.C., Baldwin, R.F., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 349–367. [Google Scholar]
- Muratet, A.; Lorrilliere, R.; Clergeau, P.; Fontaine, C. Evaluation of landscape connectivity at community level using satellite-derived NDVI. Landsc. Ecol. 2013, 28, 95–105. [Google Scholar] [CrossRef]
- Szabo, S.; Novak, T.; Elek, Z. Distance models in ecological network management: A case study of patch connectivity in a grassland network. J. Nat. Conserv. 2012, 20, 293–300. [Google Scholar] [CrossRef]
- Arellano, B.; Roca, J. Landscapes Impacted by Light. ISPRS Int. Arch. Photogramm. Remote. Sens. Spat. Inf. Sci. 2016, XLI–B8, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J.; Gaston, S.; Bennie, J.; Hopkins, J. Benefits and costs of artificial nighttime lighting of the environment. Environ. Rev. 2015, 23, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biol. Rev. Camb. Philos. Soc. 2013, 88, 912–927. [Google Scholar] [CrossRef]
- Marcantonio, M.; Pareeth, S.; Rocchini, D.; Metz, M.; Garzon-Lopez, C.X.; Neteler, M. The integration of Artificial Night-Time Lights in landscape ecology: A remote sensing approach. Ecol. Complex. 2015, 22, 109–120. [Google Scholar] [CrossRef]
- Hoelker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef]
- Davies, T.W.; Bennie, J.; Gaston, K.J. Street lighting changes the composition of invertebrate communities. Biol. Lett. 2012, 8, 764–767. [Google Scholar] [CrossRef]
- Gaston, K.J.; Bennie, J. Demographic effects of artificial nighttime lighting on animal populations. Environ. Rev. 2014, 22, 323–330. [Google Scholar] [CrossRef]
- Firebaugh, A.; Haynes, K.J. Experimental tests of light-pollution impacts on nocturnal insect courtship and dispersal. Oecologia 2016, 182, 1203–1211. [Google Scholar] [CrossRef]
- Rodrigues, P.; Aubrecht, C.; Gil, A.; Longcore, T.; Elvidge, C. Remote sensing to map influence of light pollution on Cory’s shearwater in São Miguel Island, Azores Archipelago. Eur. J. Wildl. Res. 2011, 58, 147–155. [Google Scholar] [CrossRef]
- Elvidge, C.D.; Baugh, K.E.; Zhizhin, M.; Hsu, F.-C. Why VIIRS data are superior to DMSP for mapping nighttime lights. Proc. Asia Pac. Adv. Netw. 2013, 35, 62. [Google Scholar] [CrossRef] [Green Version]
- Theobald, D.M.; Reed, S.E.; Fields, K.; Soulé, M. Connecting natural landscapes using a landscape permeability model to prioritize conservation activities in the United States. Conserv. Lett. 2012, 5, 123–133. [Google Scholar] [CrossRef]
- Miao, Z.; Pan, L.; Wang, Q.; Chen, P.; Yan, C.; Liu, L. Research on Urban Ecological Network Under the Threat of Road Networks—A Case Study of Wuhan. ISPRS Int. J. Geo-Inf. 2019, 8, 342. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Deng, L.; Dong, S.; Zhao, Q.; Yang, J.; Wang, C. Landscape connectivity dynamics based on network analysis in the Xishuangbanna Nature Reserve, China. Acta Oecol. 2014, 55, 66–77. [Google Scholar] [CrossRef]
- Saura, S.; Estreguil, C.; Mouton, C.; Rodríguez-Freire, M. Network analysis to assess landscape connectivity trends: Application to European forests (1990–2000). Ecol. Indic. 2011, 11, 407–416. [Google Scholar] [CrossRef]
- Liu, S.; Dong, Y.; Deng, L.; Liu, Q.; Zhao, H.; Dong, S. Forest fragmentation and landscape connectivity change associatedwith road network extension and city expansion: A case study in the Lancang River Valley. Ecol. Indic. 2014, 36, 160–168. [Google Scholar] [CrossRef]
- GSFC; JPSS; CMO. Joint Polar Satellite System (JPSS) VIIRS Imagery Products Algorithm Theoretical Basis Document (ATBD); NASA: Washington, DC, USA, 2014. [Google Scholar]
- Kamel Didan, A.B.M.; Solano, R.; Huete, A. MODIS_VI_UsersGuide_June_2015_C6. 2015. Available online: https://vip.arizona.edu/documents/MODIS/MODIS_VI_UsersGuide_June_2015_C6.pdf (accessed on 23 November 2020).
- Borowik, T.; Pettorelli, N.; Sönnichsen, L.; Jędrzejewska, B. Normalized difference vegetation index (NDVI) as a predictor of forage availability for ungulates in forest and field habitats. Eur. J. Wildl. Res. 2013. [Google Scholar] [CrossRef]
- Kerr, J.T.; Ostrovsky, M. From space to species: Ecological applications for remote sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Zhang, Q.; Schaaf, C.; Seto, K.C. The Vegetation Adjusted NTL Urban Index: A new approach to reduce saturation and increase variation in nighttime luminosity. RSEnv 2013, 129, 32–41. [Google Scholar] [CrossRef]
- Yi, L.; Yu, Z.Y.; Qian, J.; Kobuliev, M.; Chen, C.L.; Xing, X.W. Evaluation of the heterogeneity in the intensity of human interference on urbanized coastal ecosystems: Shenzhen (China) as a case study. Ecol. Indic. 2021, 122, 10. [Google Scholar] [CrossRef]
- Geri, F.; Amici, V.; Rocchini, D. Human activity impact on the heterogeneity of a Mediterranean landscape. Appl. Geogr. 2010, 30, 370–379. [Google Scholar] [CrossRef]
- Svechkina, A.; Portnov, B.A.; Trop, T. The impact of artificial light at night on human and ecosystem health: A systematic literature review. Landsc. Ecol. 2020, 35, 1725–1742. [Google Scholar] [CrossRef]
- Pothukuchi, K. City Light or Star Bright: A Review of Urban Light Pollution, Impacts, and Planning Implications. J. Plan. Lit. 2021, 36, 155–169. [Google Scholar] [CrossRef]
- Bolliger, J.; Hennet, T.; Wermelinger, B.; Bosch, R.; Pazur, R.; Blum, S.; Haller, J.; Obrist, M.K. Effects of traffic-regulated street lighting on nocturnal insect abundance and bat activity. Basic Appl. Ecol. 2020, 47, 44–56. [Google Scholar] [CrossRef]
- Owens, A.C.S.; Lewis, S.M. Effects of artificial light on growth, development, and dispersal of two North American fireflies (Coleoptera: Lampyridae). J. Insect Physiol. 2021, 130, 9. [Google Scholar] [CrossRef]
- Levin, N. The impact of seasonal changes on observed nighttime brightness from 2014 to 2015 monthly VIIRS DNB composites. RSEnv 2017, 193, 150–164. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Yu, B.L.; Ta, N.; Shi, K.F.; Yang, C.S.; Wang, C.X.; Zhao, X.Z.; Deng, S.Q.; Wu, J.P. Delineating Seasonal Relationships Between Suomi NPP-VIIRS Nighttime Light and Human Activity Across Shanghai, China. IEEE J. Sel. Top. Appl. Earth Obs. Remote. Sens. 2019, 12, 4275–4283. [Google Scholar] [CrossRef]
- Mu, H.W.; Li, X.C.; Du, X.P.; Huang, J.X.; Su, W.; Hu, T.Y.; Wen, Y.A.; Yin, P.Y.; Han, Y.; Xue, F. Evaluation of Light Pollution in Global Protected Areas from 1992 to 2018. Remote Sens. 2021, 13, 1849. [Google Scholar] [CrossRef]
- Gaston, K.J.; Davies, T.W.; Bennie, J.; Hopkins, J. Review: Reducing the ecological consequences of night-time light pollution: Options and developments. J. Appl. Ecol. 2012, 49, 1256–1266. [Google Scholar] [CrossRef] [Green Version]
- Kamrowski, R.L.; Sutton, S.G.; Tobin, R.C.; Hamann, M. Balancing artificial light at night with turtle conservation? Coastal community engagement with light-glow reduction. Environ. Conserv. 2015, 42, 171–181. [Google Scholar] [CrossRef]
- Li, F.; Yan, Q.W.; Bian, Z.F.; Liu, B.L.; Wu, Z.H. A POI and LST Adjusted NTL Urban Index for Urban Built-Up Area Extraction. Sensors 2020, 20, 2918. [Google Scholar] [CrossRef] [PubMed]
- Tabor, R.A.; Perkin, E.K.; Beauchamp, D.A.; Britt, L.L.; Haehn, R.; Green, J.; Robinson, T.; Stolnack, S.; Lantz, D.W.; Moore, Z.J. Artificial lights with different spectra do not alter detrimental attraction of young Chinook salmon and sockeye salmon along lake shorelines. Lake Reserv. Manag. 2021, 1–12. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VIIRS Nighttime Brightness (nW/cm2/sr) | |||||||
|---|---|---|---|---|---|---|---|
| NDVI | ≤1 | 1–10 | 10–20 | 20–30 | 30–40 | >40 | Sum |
| ≤0 | 2.276% | 1.529% | 0.358% | 0.156% | 0.023% | 0.014% | 4.357% |
| 0–0.1 | 1.383% | 0.646% | 0.108% | 0.035% | 0.011% | 0.007% | 2.190% |
| 0.1–0.3 | 3.089% | 2.655% | 1.616% | 1.416% | 0.991% | 1.046% | 10.813% |
| 0.3–0.5 | 29.442% | 13.494% | 2.970% | 1.788% | 0.665% | 0.323% | 48.681% |
| 0.5–0.7 | 28.549% | 4.786% | 0.303% | 0.082% | 0.019% | 0.005% | 33.745% |
| >0.7 | 0.181% | 0.028% | 0.006% | 0.000% | 0.000% | 0.000% | 0.215% |
| sum | 64.920% | 23.139% | 5.361% | 3.477% | 1.709% | 1.395% | 100.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Liu, Y.; Fang, J. Ecological Corridor Construction Based on Least-Cost Modeling Using Visible Infrared Imaging Radiometer Suite (VIIRS) Nighttime Light Data and Normalized Difference Vegetation Index. Land 2021, 10, 782. https://doi.org/10.3390/land10080782
Hu J, Liu Y, Fang J. Ecological Corridor Construction Based on Least-Cost Modeling Using Visible Infrared Imaging Radiometer Suite (VIIRS) Nighttime Light Data and Normalized Difference Vegetation Index. Land. 2021; 10(8):782. https://doi.org/10.3390/land10080782
Chicago/Turabian StyleHu, Jiameng, Yanfang Liu, and Jian Fang. 2021. "Ecological Corridor Construction Based on Least-Cost Modeling Using Visible Infrared Imaging Radiometer Suite (VIIRS) Nighttime Light Data and Normalized Difference Vegetation Index" Land 10, no. 8: 782. https://doi.org/10.3390/land10080782