
Windows into the Recent Past: Simple Biotic Indices to Assess Hydrological Stability in Small, Isolated Ponds

Abstract
:
1. Introduction
Objectives
- quantify the hydrological stability of the ponds in lowland wetlands using simple hydrological parameters derived from the analysis of freely available maps and satellite images.
- describe and statistically confirm the relationships between hydrological stability over several decades and the taxonomic composition of the freshwater macroinvertebrates inhabiting them.
- determine the importance of hydrological stability in relation to the other abiotic parameters in the studied water bodies for the taxonomic composition of the macrofauna.
- develop and test protocols for the multimetric biotic indices to reconstruct the hydrological stability of small reservoirs over several decades. It was assumed that the indices will be formatted to allow for their improvement and adaptation to other similar environments.
- an additional aim of the study was to assess whether the non-lethal methods of obtaining information on the taxonomic composition of invertebrates could be sufficiently effective in assessing the hydrological stability of ponds.
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Testing the Significance of Abiotic Parameters
2.4. Data Preparation for Hydrological Stability Assessment
- The hydrological parameters were normalized.
- Taxa that were found to have a significant explanatory power in the multiple regression models at a level of p < 0.05 were included in the constructed index after normalization and then (Figure 3),
- multiplied by the R2 values in the multiple regression model. The resulting values were assigned a positive or negative value based on their relationship with the hydrological parameter.
- The index value was obtained by taking the weighted average (the weights were determined by the R2 values) of the standardized values from point c., which were then placed on a scale of 0–1.
3. Results
- The average pond surface area during the 10 years prior to sampling, expressed as a percentage of the maximum surface area (ASA10) on a scale of 0–1.
- The number of years in which episodes of complete drying of the pond were recorded during the 10 years prior to sampling (NCD10) on a scale of 0–10.
- The smallest recorded pond surface area in the 5 years prior to sampling, expressed as a percentage of the maximum surface area during the past 20 years (SSA10) on a scale of 0–1.
3.1. The Indices
(%St ∙ −0.173 + 0.048) + (%Sn ∙ −0.013 + 0.167) + (%Et ∙ 0.040 − 0.032) + (%Hm ∙ 0.248 − 0.066)
(%St ∙ −0.148 + 0.041) + (%Sn ∙ −0.011 + 0.139) + (%Et ∙ 0.032 − 0.026) + (%Hm ∙ 0.186 − 0.050)
0.082) + (%No ∙ 0.033 − 0.0388) + (%Gl ∙ −0.043 + 0.038) + (%Dl ∙ 0.192 − 0.020) + (%Et ∙ 0.0413 − 0.0323)
(%Str ∙ −0.227 + 0.106) + (%No ∙ −0.025 − 0.029) + (%Gl ∙ −0.057 + 0.050) + (%Dl ∙ 0.168 − 0.018) + (%Et ∙ 0.033 − 0.026)
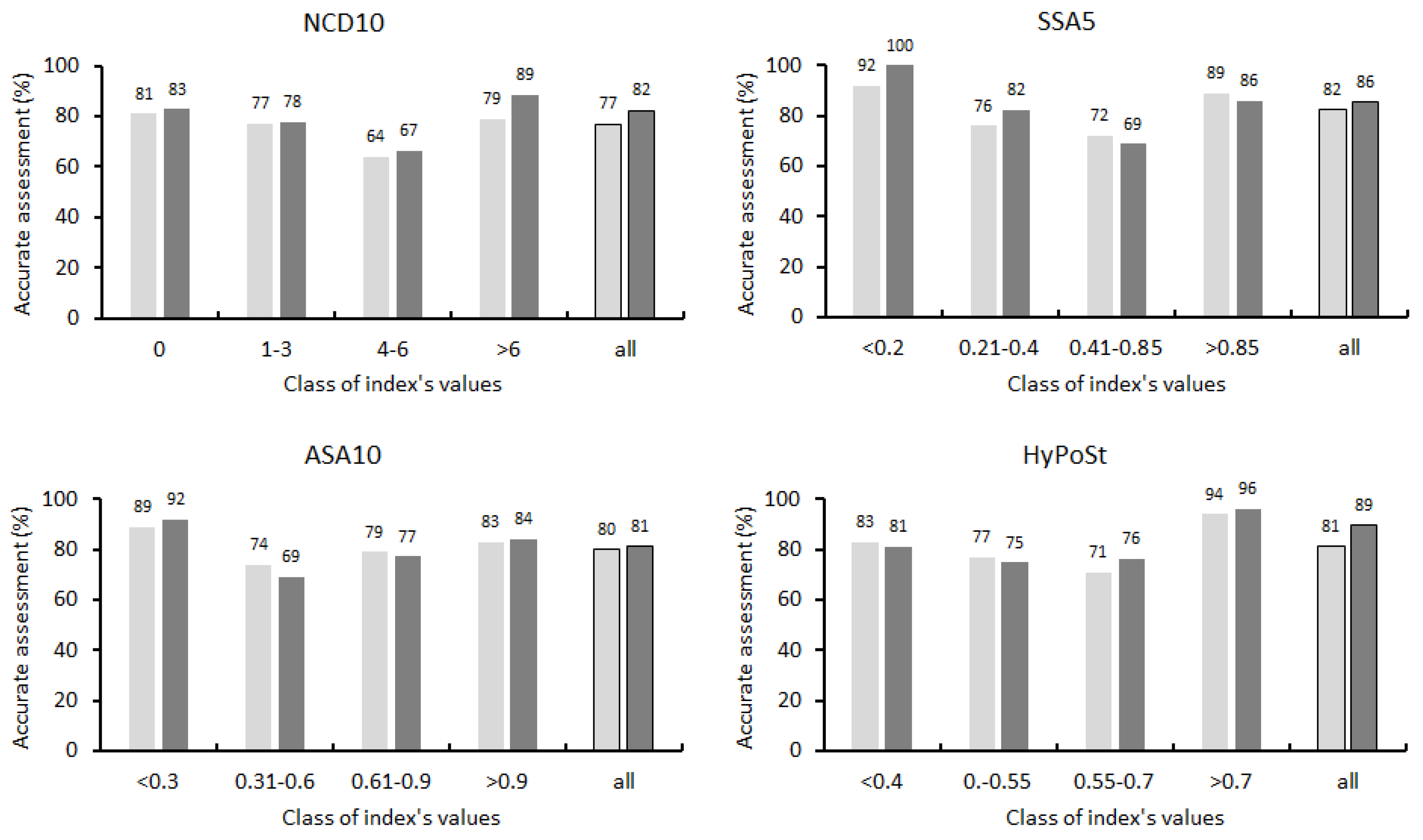
3.2. Testing the Indices Using Descriptive Statistics
4. Discussion
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Céréghino, R.; Biggs, J.; Oertli, B.; Declerck, S. The ecology of European ponds: Defining the characteristics of a neglected freshwater habitat. Hydrobiology 2008, 597, 1–6. [Google Scholar] [CrossRef]
- Gökçe, D. Wetland importance and management. In Wetlands Management—Assessing Risk and Sustainable Solutions; Gökçe, D., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Scheffer, M.; Van Geest, G.J.; Zimmer, K.; Jeppesen, E.; Søndergaard, M.; Butler, M.G.; De Meester, A.L. Small habitat size and isolation can promote species richness: Second-order effects on biodiversity in shallow lakes and ponds. Oikos 2006, 112, 227–231. [Google Scholar] [CrossRef]
- Oertli, B.; Parris, K.M. Toward management of urban ponds for freshwater biodiversity. Ecosphere 2019, 10, e02810. [Google Scholar] [CrossRef]
- Horváth, Z.; Ptacnik, R.; Vad, C.F.; Chase, J.M. Habitat loss over six decades accelerates regional and local biodiversity loss via changing landscape connectance. Ecol. Lett. 2019, 22, 1019–1027. [Google Scholar] [CrossRef]
- Jeffries, M.J.; Epele, L.B.; Studinski, J.M.; Vad, C.F. Invertebrates in Temporary Wetland Ponds of the Temperate Biomes. In Invertebrates in Freshwater Wetlands; Batzer, D., Boix, D., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Nieoczym, M.; Stryjecki, R.; Buczyński, P.; Płaska, W.; Kloskowski, J. Differential abundance, composition and mesohabitat use by aquatic macroinvertebrate taxa in ponds with and without fish. Aquat. Sci. 2023, 85, 25. [Google Scholar] [CrossRef]
- Harper, D.; Mekotova, J.; Hulme, S.; White, J.; Hall, J. Habitat heterogeneity and aquatic invertebrate diversity in floodplain forests. Glob. Ecol. Biogeogr. 1997, 6, 275–285. [Google Scholar] [CrossRef]
- Buczyńska, E.; Buczyński, P.; Lechowski, L.; Stryjecki, R. Fish pond complexes as refugia of aquatic invertebrates (Odonata, Coleoptera, Heteroptera, Trichoptera, Hydrachnidia): A case study of the pond complex in Zalesie Kańskie (Central-East Poland). Nat. Conserv. 2007, 64, 39–55. [Google Scholar]
- Biggs, J.; Bilton, D.; Williams, P. Temporary pond of eastern Poland: An initial assessment of their importance for nature conservation. Arch. Sci. 2004, 57, 73–84. [Google Scholar] [CrossRef]
- Bloechl, A.; Koenemann, S.; Philippi, B.; Melber, A. Abundance, diversity and succession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in the North German Lowlands. Limnologica 2010, 40, 215–225. [Google Scholar] [CrossRef]
- Briers, R.A.; Biggs, J. Indicator taxa for the conservation of pond invertebrate diversity. Aquat. Conserv. 2003, 13, 323–330. [Google Scholar] [CrossRef]
- Céréghino, R.; Boix, D.; Cauchie, H.M.; Martens, K.; Oertli, B. The ecological role of ponds in a changing world. Hydrobiologia 2014, 723, 1–6. [Google Scholar] [CrossRef]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.; Lachavanne, J.B. Does size matter? The relationship between pond area and biodiversity. Biol. Cons. 2002, 104, 59–70. [Google Scholar] [CrossRef]
- Koperski, P.; Narożniak, E.; Mętrak, M. Composition of mollusk communities as a proxy in studies on seasonal dynamics of astatic water bodies. Monit. Środ Przyr. 2014, 15, 23–31. [Google Scholar]
- Meutter, F.V.D.; Meester, L.D.; Stoks, R. Metacommunity structure of pond macroinvertebrates: Effects of dispersal mode and generation time. Ecology 2007, 88, 1687–1695. [Google Scholar] [CrossRef]
- Jurkiewicz-Karnkowska, E. Effects of habitat conditions on the diversity and abundance of molluscs in floodplain water bodies of different permanence of flooding. Pol. J. Ecol. 2011, 59, 165–178. [Google Scholar]
- Ostad-Ali-Askari, K. Review of the effects of the anthropogenic on the wetland environment. Appl. Water Sci. 2022, 12, 260. [Google Scholar] [CrossRef]
- Koperski, P. Hydrological instability of ponds reduces functional diversity of freshwater molluscs in protected wetlands. Wetlands 2022, 42, 42. [Google Scholar] [CrossRef]
- CLIMATE-ADAPT. Available online: https://climate-adapt.eea.europa.eu/en/countries-regions/countries/poland (accessed on 15 March 2021).
- Matthews, J. Anthropogenic climate change impacts on ponds: A thermal mass perspective. BioRisk 2010, 5, 193–209. [Google Scholar] [CrossRef]
- Lund, J.O.; Wissinger, S.A.; Peckarsky, B.L. Caddisfly behavioral responses to drying cues in temporary ponds: Implications for effects of climate change. Freshw. Sci. 2016, 35, 619–630. [Google Scholar] [CrossRef]
- Collinson, N.H.; Biggs, J.; Corfield, A.H.M.J.; Hodson, M.J.; Walker, D.; Whitfield, M.; Williams, P.J. Temporary and permanent ponds: An assessment of the effects of drying out on the conservation value of aquatic macroinvertebrate communities. Biol. Conserv. 1995, 74, 125–133. [Google Scholar] [CrossRef]
- Marcisz, K.; Kołaczek, P.; Gałka, M.; Diaconu, A.C.; Lamentowicz, M. Exceptional hydrological stability of a Sphagnum-dominated peatland over the late Holocene. Quat. Sci. Rev. 2020, 231, 106180. [Google Scholar] [CrossRef]
- Brendonck, L.; Jocqué, M.; Tuytens, K.; Timms, B.V.; Vanschoenwinkel, B. Hydrological stability drives both local and regional diversity patterns in rock pool metacommunities. Oikos 2015, 124, 741–749. [Google Scholar] [CrossRef]
- Wissinger, S.A.; Greig, H.; McIntosh, A. Absence of species replacements between permanent and temporary lentic communities in New Zealand. J. N. Am. Benthol. Soc. 2009, 28, 12–23. [Google Scholar] [CrossRef]
- Carvalho, L.; Mackay, E.B.; Cardoso, A.C.; Baattrup-Pedersen, A.; Birk, S.; Blackstock, K.L.; Solheim, A.L. Protecting and restoring Europe’s waters: An analysis of the future development needs of the Water Framework Directive. Sci. Total Environ. 2019, 658, 1228–1238. [Google Scholar] [CrossRef]
- McLean, K.I.; Mushet, D.M.; Newton, W.E.; Sweetman, J.N. Long-term multidecadal data from a prairie-pothole wetland complex reveal controls on aquatic-macroinvertebrate communities. Ecol. Ind. 2021, 126, 107678. [Google Scholar] [CrossRef]
- Stamenković, O.; Stojković Piperac, M.; Milošević, D.; Buzhdygan, O.Y.; Petrović, A.; Jenačković, D.; Simić, V. Anthropogenic pressure explains variations in the biodiversity of pond communities along environmental gradients: A case study in south-eastern Serbia. Hydrobiologia 2019, 838, 65–83. [Google Scholar] [CrossRef]
- Stamenković, O.; Stojković Piperac, M.; Milošević, D.; Čerba, D.; Cvijanović, D.; Gronau, A.; Buzhdygan, O. Multiple anthropogenic pressures and local environmental gradients in ponds governing the taxonomic and functional diversity of epiphytic macroinvertebrates. Hydrobiologia 2024, 851, 45–65. [Google Scholar] [CrossRef]
- Sebastián-González, E.; Sánchez-Zapata, J.A.; Botella, F. Agricultural ponds as alternative habitat for waterbirds: Spatial and temporal patterns of abundance and management strategies. Eur. J. Wild Res. 2010, 56, 11–20. [Google Scholar] [CrossRef]
- Sauermann, H.; Vohland, K.; Antoniou, V.; Balázs, B.; Göbel, C.; Karatzas, K.; Winter, S. Citizen science and sustainability transitions. Res. Policy 2020, 49, 103978. [Google Scholar] [CrossRef]
- Metcalfe, A.N.; Kennedy, T.A.; Mendez, G.A.; Muehlbauer, J.D. Applied citizen science in freshwater research. WIRE Water 2020, 9, e1578. [Google Scholar] [CrossRef]
- Kirschke, S.; Bennett, C.; Bigham Ghazani, A.; Franke, C.; Kirschke, D.; Lee, Y.; Loghmani Khouzani, S.T.; Nath, S. Citizen science projects in freshwater monitoring. From individual design to clusters? J. Environ. Manag. 2022, 309, 114714. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska-Barna, I. Macroinvertebrate benthic communities in the macrophyte-dominated Lake Łuknajno (Northeastern Poland). Oceanol. Hydrobiol. Stud. 2007, 36 (Suppl. 4), 29–38. [Google Scholar]
- Falarz, M.; Bielec-Bąkowska, Z.; Wypych, A.; Matuszko, D.; Niedźwiedź, T.; Pińskwar, I.; Wibig, J. Climate Change in Poland—Summary, Discussion and Conclusion. In Climate Change in Poland: Past, Present, Future; Falarz, M., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 561–581. [Google Scholar]
- Łupikasza, E.; Małarzewski, Ł. Precipitation change. In Climate Change in Poland: Past, Present, Future; Falarz, M., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 349–373. [Google Scholar]
- Williams, D.D. Temporary ponds and their invertebrate communities. Aquat. Cons. 1997, 7, 105–117. [Google Scholar] [CrossRef]
- Sayer, C.; Andrews, K.; Shilland, E.; Edmonds, N.; Edmonds-Brown, R.; Patmore, I.; Axmacher, J. The role of pond management for biodiversity conservation in an agricultural landscape. Aquat. Conserv. 2012, 22, 626–638. [Google Scholar] [CrossRef]
- Epele, L.B.; Grech, M.G.; Williams-Subiza, E.A.; Stenert, C.; McLean, K.; Greig, H.S.; Maltchik, L.; Pires, M.M.; Bird, M.S.; Boissezon, A.; et al. Perils of life on the edge: Climatic threats to global diversity patterns of wetland macroinvertebrates. Sci. Total Environ. 2022, 820, 153052. [Google Scholar] [CrossRef] [PubMed]
- Epele, L.B.; Williams-Subiza, E.A.; Bird, M.S.; Boissezon, A.; Boix, D.; Demierre, E.; Fair, C.G.; García, P.E.; Gascón, S.; Grech, M.G.; et al. A global assessment of environmental and climate influences on wetland macroinvertebrate community structure and function. Glob. Chang. Biol. 2024, 30, e17173. [Google Scholar] [CrossRef]
- Boix, D.; Sala, J.; Moreno-Amich, R. The faunal composition of Espolla pond (NE Iberian peninsula): The neglected biodiversity of temporary waters. Wetlands 2001, 21, 577–592. [Google Scholar] [CrossRef]
- Jurkiewicz-Karnkowska, E. Aquatic mollusc communities in riparian sites of different size, hydrological connectivity and succession stage. Pol. J. Ecol. 2008, 56, 99. [Google Scholar]
- Céréghino, R.; Ruggiero, A.; Marty, P.; Angélibert, S. Biodiversity and distribution patterns of freshwater invertebrates in farm ponds of a south-western French agricultural landscape. Hydrobiologia 2008, 597, 43–51. [Google Scholar] [CrossRef]
- Cockroft, R.J.; Jenkins, W.R.; Irwin, A.G.; Norman, S.; Brown, K.C. Emergence timing and voltinism of phantom midges, Chaoborus spp., in the UK. Web Ecol. 2022, 22, 101–108. [Google Scholar] [CrossRef]
- Lee, C.Y.; Kim, D.G.; Baek, M.J.; Choe, L.J.; Bae, Y.J. Life history and emergence pattern of Cloeon dipterum (Ephemeroptera: Baetidae) in Korea. Environ. Entomol. 2013, 42, 1149–1156. [Google Scholar] [CrossRef]
- Bilton, D.T. Dispersal in Dytiscidae. In Ecology, Systematics, and the Natural History of Predaceous Diving Beetles (Coleoptera: Dytiscidae); Yee, D.A., Ed.; Springer: Cham, Switzerland, 2023. [Google Scholar] [CrossRef]
- Unglaub, B.; Steinfartz, S.; Kühne, D.; Haas, A.; Schmidt, B.R. The relationships between habitat suitability, population size and body condition in a pond-breeding amphibian. Basic. Appl. Ecol. 2018, 27, 20–29. [Google Scholar] [CrossRef]
- Bănăduc, D.; Sas, A.; Cianfaglione, K.; Barinova, S.; Curtean-Bănăduc, A. The role of aquatic refuge habitats for fish, and threats in the context of climate change and human impact, during seasonal hydrological drought in the Saxon Villages area (Transylvania, Romania). Atmosphere 2021, 12, 1209. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Classes of Parameters | Kruskal–Wallis Test | ||||
|---|---|---|---|---|---|---|
| NCD10 | 0 | 1–3 | 4–6 | >6 | 1.3400 × 10−11 | |
| 0 | 0.020 | 1.3780 × 10−8 | <1 × 10−8 | |||
| 1–3 | 4.220 | 3.4580 × 10−4 | <1 × 10−8 | |||
| 4–6 | 9.691 | 6.039 | <1 × 10−8 | |||
| >6 | 25.210 | 20.410 | 11.550 | |||
| SSA 5 | <20 | 20–50 | 60–80 | >80 | 6.6590 × 10−13 | |
| <20 | 0.000 | <1 × 10−9 | <1 × 10−9 | |||
| 20–50 | 10.320 | 0.010 | <1 × 10−9 | |||
| 60–80 | 13.110 | 4.571 | 4.1220 × 10−6 | |||
| >80 | 21.120 | 13.050 | 7.714 | |||
| ASA 10 | <35 | 35–60 | 60–85 | >85 | 6.8630 × 10−12 | |
| <35 | <1 × 10−5 | <1 × 10−5 | <1 × 10−5 | |||
| 35–60 | 13.370 | 2.9470 × 10−4 | <1 × 10−5 | |||
| 60–85 | 19.380 | 6.108 | 1.1040 × 10−4 | |||
| >85 | 23.520 | 11.780 | 6.496 | |||
| HyPoSt | <0.2 | 20–60 | 60–80 | >80 | 2.3900 × 10−14 | |
| <0.2 | <1 × 10−11 | <1 × 10−11 | <1 × 10−11 | |||
| 20–60 | 15.630 | <1 × 10−11 | <1 × 10−11 | |||
| 60–80 | 25.580 | 14.610 | 4.9140 × 10−11 | |||
| >80 | 35.550 | 27.620 | 11.310 | |||
| Index | Classes of Parameters | Kruskal–Wallis Test (p) | ||||
|---|---|---|---|---|---|---|
| NCDMi | <0.2 | 0.2–0.35 | 0.35–0.55 | >0.55 | 2.25 × 10−10 | |
| <0.2 | 0.020 | 2.81 × 10−8 | <1 × 10−8 | |||
| 0.2–0.35 | 4.222 | 0.002 | <1 × 10−8 | |||
| 0.35–0.55 | 9.453 | 5.285 | 2.85 × 10−6 | |||
| >0.55 | 19.517 | 14.190 | 7.846 | |||
| SSAMi | <0.25 | 0.25–0.4 | 0.4–0.65 | >0.65 | 3.21 × 10−12 | |
| <0.25 | 1.38 × 10−7 | <1 × 10−7 | <1 × 10−7 | |||
| 0.25–0.4 | 8.905 | 3.22 × 10−6 | <1 × 10−7 | |||
| 0.4–0.65 | 15.341 | 7.802 | 6.27 × 10−4 | |||
| >0.65 | 19.784 | 13.204 | 5.796 | |||
| ASAMi | <0.2 | 0.2–0.4 | 0.4–0.8 | >0.8 | 7.16 × 10−11 | |
| <0.2 | 0.010 | <1 × 10−7 | <1 × 10−7 | |||
| 0.2–0.4 | 4.586 | 3.02 × 10−7 | <1 × 10−7 | |||
| 0.4–0.8 | 11.623 | 8.636 | 2.03 × 10−4 | |||
| >0.8 | 15.330 | 13.121 | 6.2520 | |||
| HyPoStMi | <0.2 | 20–60 | 60–80 | >80 | 8.27 × 10−13 | |
| <0.2 | 8.89 × 10−10 | <1 × 10−10 | <1 × 10−10 | |||
| 20–60 | 10.591 | 5.52 × 10−7 | <1 × 10−10 | |||
| 60–80 | 16.202 | 8.426 | 3.13 × 10−5 | |||
| >80 | 22.349 | 16.435 | 6.9700 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koperski, P. Windows into the Recent Past: Simple Biotic Indices to Assess Hydrological Stability in Small, Isolated Ponds. Water 2024, 16, 1206. https://doi.org/10.3390/w16091206
Koperski P. Windows into the Recent Past: Simple Biotic Indices to Assess Hydrological Stability in Small, Isolated Ponds. Water. 2024; 16(9):1206. https://doi.org/10.3390/w16091206
Chicago/Turabian StyleKoperski, Paweł. 2024. "Windows into the Recent Past: Simple Biotic Indices to Assess Hydrological Stability in Small, Isolated Ponds" Water 16, no. 9: 1206. https://doi.org/10.3390/w16091206