1. Introduction
Most aquatic environments enjoy thermal stability due to the greater thermal carrying capacity of water when compared to air. As a result, many organisms living in these environments, including many fish species, have a narrow tolerance for rapid changes in ambient water temperature and respond [
1] with a cascade of physiological conditions when exposed to rapid fluctuations in water temperature [
2]. Most studies to date have investigated the effects of slowly raising water temperatures [
1,
2,
3], which often stress exposed fish through the rapidly decreasing oxygen carrying capacity of warmer waters coupled with the increased metabolic rate of the fish at a higher temperature. In contrast, cold shock, a condition in which ambient water temperatures drop rapidly, resulting in adverse physiological or behavioral consequences [
4], has received less attention (however, see the reviews by [
5,
6]) and the mechanisms by which fish are affected are less understood than those under heat-stress conditions.
The few existing studies [
4,
5,
7] and EPA [
8] guidelines for thermal effluents use death as an endpoint, thus providing little guidance on minimizing the impacts of cold shock on aquatic organisms. However, studies of heat shock and acclimation to increased environmental temperatures [
1,
9], which provide insight into the physiological responses of aquatic organisms to rapid temperature changes, indicate a wide range of physiological endpoints that may be utilized to guide the development of operational procedures to minimize the effects of cold shock [
6]. Among thermal induced stressors, changes in hematocrit, white blood cell counts, cortisol concentrations, and behavior have all been recognized as indicators of the adverse effects of rapid thermal change [
10,
11,
12].
Steam electric power generation is the nation’s largest surface water consumer [
13], and past data show that many fish species seek refuge in sustained heated effluents during winter [
4,
6,
9]. However, recent shifts in technologies are leading to increased employment of combined-cycle natural gas-fired facilities. This technology permits a rapid startup and shutdown process, with temperature stability only in the order of hours rather than weeks or months for traditional coal powered plants [
14,
15]. According to the U.S Energy Information Administration (
http://www.eia.gov/naturalgas/ (accessed on 7 October 2023)), from 1999 through 2010 natural gas-powered generation increased nearly 78% and is likely to continue to increase through 2050. However, the increased efficiency and ability to provide on-demand power generation is not without concern. Understanding the effects of cold shock and possibly developing mitigating protocols for the abatement of such phenomena have become more urgent as many base-load steam-electric facilities (i.e., coal) with sustained thermal effluents are retired and retrofitted with natural gas-fired combined-cycle facilities. Combined-cycle natural gas facilities have the capability to ramp up or shut down power generation within hours or even minutes. Effluent temperatures in the winter may drop from a high operational effluent temperature of 25 °C to 30 °C to an ambient temperature of 2 °C at a rate of greater than 10 °C/h. Thermal cycling at these rates and magnitudes can occur on a daily, weekly, or monthly basis depending on electrical demand [
14,
15]. In addition, the volume of river water will determine the extent of the aquatic habitat affected by the cycling thermal effluent and the ability of fish to possibly avoid the area subject to rapid heating and cooling cycles.
It is well established that long-term thermal effluent discharge attracts large fish populations into the effluent impacted mixing zone downstream of coal-fire and nuclear power plants [
4,
9,
16]. However, fish in these effluents acclimate to relatively stable conditions that may last for months—and even years if multiple units are available—with shutdowns and the associated reduction in the heat load only occurring because of emergency shutdowns or scheduled maintenance outages. When emergency shutdowns occur at these facilities, extensive fish deaths have been reported, indicating the adverse effects of cold shock events to resident fish populations. Interestingly, in a study by [
9], it was reported that fish had been residing in a thermal effluent for most of the winter, with half the fish not returning during the remainder of the study. However, it was beyond the scope of that study to determine whether fish experienced a fatal cold shock, did not return due to repulsion, or simply did not return to the previously inhabited area. It is currently unknown whether effluents with rapidly cycling temperatures will attract fish towards the effluents or may deter fish from areas in the vicinity of the thermal effluent, and whether the following cold shock will induce stress in the resident populations of fish in the receiving waters. These effects, and their directionality, may also vary based on the season, average ambient water temperature outside the thermal effluent plume, and intermittency of the thermal plume.
In this study, the hypotheses tested were as follows: (i) thermal effluents will attract fish quickly and (ii) the subsequent rapid fluctuation of the effluent temperature will induce stress and reduce survival in resident fish populations. To test the assumptions of these hypotheses, three experiments were conducted. First, three species of fish, ranging in size from 2 g to 1200 g, were exposed to a controlled cold shock in the laboratory to assess the effects of temperature differential, rate of temperature decline, and fish size. Then, the behavior of smallmouth bass Micropterus dolomieu (Lacepède 1802) was examined in a heterogeneous temperature environment to assess the behavioral preferences that may mediate the effects of rapid thermal cooling in natural environments. Lastly, acoustic telemetry was used to monitor native M. dolomieu residing within the vicinity of a recently constructed combine-cycle natural gas-fired power plant. Using information gleaned from these experiments, likely environmental scenarios that have the potential to impact resident fish populations were reviewed and protocols to mitigate these effects were developed. Together, these experiments provide a comprehensive assessment of the likelihood of adverse effects by rapid thermal cycling of power plants in northern latitudes.
2. Materials and Methods
Test Organisms. All experimental procedures were approved through the Institutional Animal Care and Use Committee (IACUC) at St. Cloud State University (permit # 0808) prior to any experiments taking place. The three fish species used in these experiments are native to the Upper Mississippi River and add environmental relevance to this study. Juvenile bluegill
Lepomis macrochirus (Rafinesque 1819) (50.7 ± 9.3 mm total length [mean ± standard deviation]; 2.1 ± 1.1 g wet weight) were captured by seine in the Minnesota lakes and acclimated for several weeks in the St. Cloud State University Aquatic Toxicology Laboratory in flow-through well water. Adult
L. macrochirus (174 ± 10 mm total length; 103 ± 17 g wet weight) and
M. dolomieu (341 ± 69 mm total length; 525 ± 198 g wet weight) were captured by electro-shocking several weeks prior to their use in any experiment and acclimated on-site in flow-through tanks supplied with ambient river water, as well water was not available at the field site. Mature male fathead minnows
Pimephales promelas (Rafinesque 1820) (6 months old; 59 ± 5 mm total length; 3 ± 0.1 g wet weight) were purchased from a laboratory fish supplier (Environmental Consulting and Testing;
www.ectesting.com) and acclimated in well water for several weeks prior to experimentation. All fish were fed frozen brine shrimp daily ad libitum (
Artemia ssp.; Brine Shrimp Direct,
www.brineshrimpdirect.com) and fish in all laboratory experiments were maintained at a constant 16:8 h light:dark photoperiod in 24 °C flow-through well water. If a group of fish needed to be acclimated to another temperature prior to a cold shock experiment, acclimation occurred very slowly (~1 °C/day), and fish were held at the new temperature for at least 48 h prior to the initiation of the experiment. The survival rate of the fish during the acclimation period was excellent (>95%).
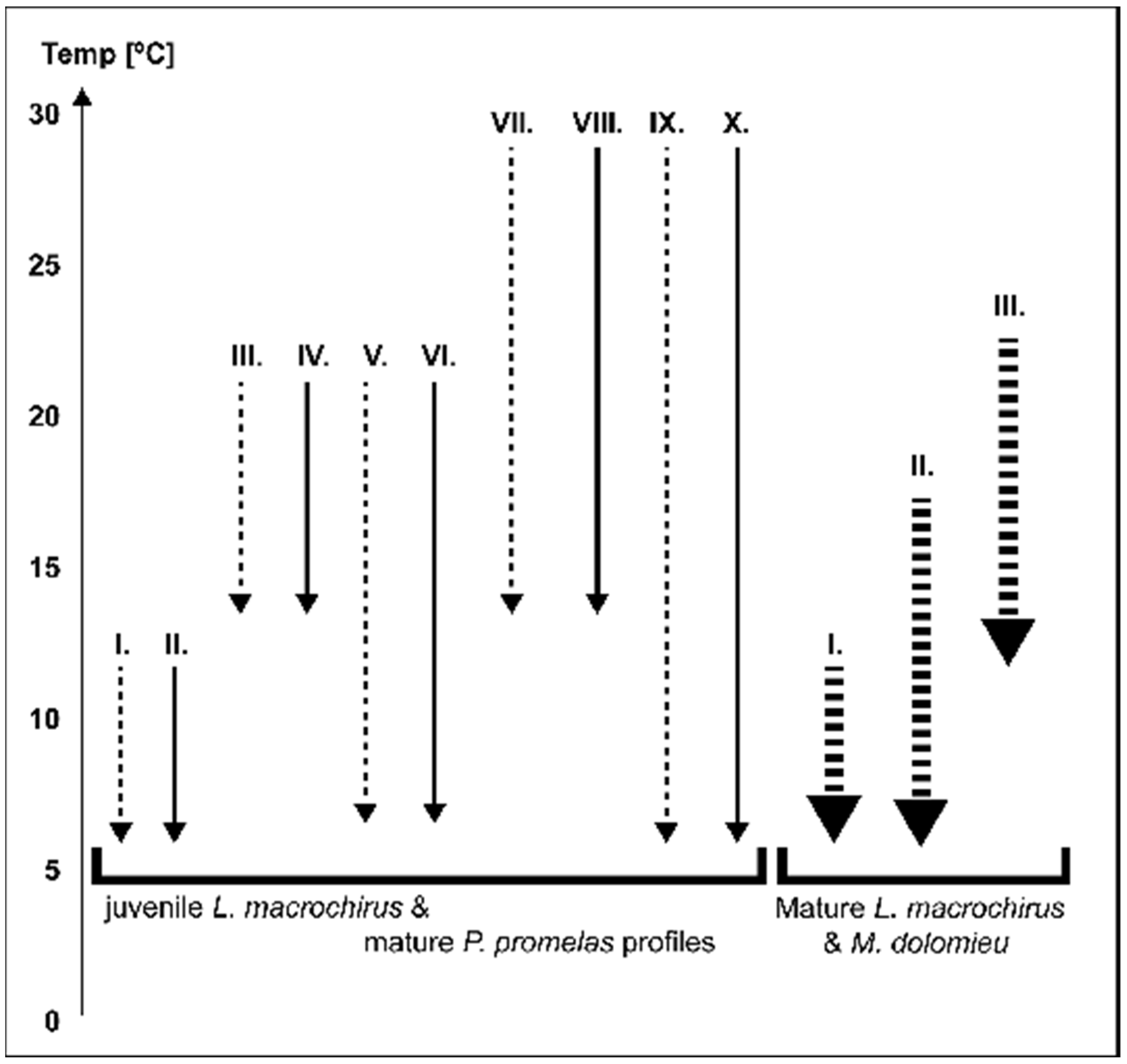
Laboratory cold shock experiment. Cold shock experiments were conducted in a controlled laboratory setting and designed to simulate environmental conditions as measured downstream of a gas-fired power plant on the Upper Mississippi River in Minneapolis, MN (
Figure 1). The experimental designs for juvenile
L. macrochirus and
P. promelas were identical and were conducted in sequence (
Table 1). Cold shock experiments for mature
L. macrochirus and
M. dolomieu were adjusted for the larger mass of these fish and each thermal profile was applied concurrently to both species (i.e., tanks holding mature
L. macrochirus and mature
M. dolomieu were adjacent to each other and received chilled water from the same source through split lines). All cold shock experiments were initiated in the morning by supplying pre-chilled water into the experimental aquarium (100 L for
P. promelas and juvenile
L. macrochirus; 400 L for mature
L. macrochirus and
M. dolomieu) to lower the water temperature at the desired rate of decline until the lower holding temperature was achieved (
Table 1,
Figure 1). Fish were then held for 48 h at the lower temperature to monitor the survival and health of the fish. In the case of the
M. dolomieu, their large size allowed for repeated sampling of blood from the caudal vasculature during the 48-h holding period.
For all species and experiments, survival throughout the cold shock and the ensuing 48-h holding period was monitored. Juvenile L. macrochirus were too small to collect sufficient blood for additional analysis. For P. promelas, mature L. macrochirus, and M. dolomieu, the ratio of blood cells to plasma (hematocrit) was assessed. In mature L. macrochirus and M. dolomieu, the abundance of white blood cells (leukocyte) and serum cortisol concentrations were also quantified. All endpoints were assessed in all surviving fish 48 h after the lower holding temperature was achieved. In addition, in M. dolomieu, endpoints were also assessed immediately following the cold shock (0 h of holding temperature) and at 3 h and 6 h after the lower holding temperature was achieved.
Hematological parameters included hematocrit and leukocyte differentials. Hematocrit determinations were made using 70 uL glass micro hematocrit tubes spun at 5000×
g for 5 min in a hematocrit centrifuge. Packed red blood cell levels were determined as a percentage of the whole blood volume using a hematocrit ruler (Critocaps, Thomas Scientific;
www.thomassci.com). Mean hematocrit values with standard deviation for each fish species per experiment were calculated [
2]. White blood cell differential analyses were performed using a manual count of the blood smears stained with modified Wright’s stain (Accustain
®, Sigma-Aldrich;
www.sigmaaldrich.com). Leukocytes were counted in multiple, successive fields of view until a total of 100 cells was reached. Leukocytes were recorded as lymphocytes, monocytes, or granulocytes, following basic identification parameters such as those described by [
17].
Plasma cortisol levels were measured using a competitive antibody-capture enzyme immunoassay purchased from Cayman Chemicals (
www.caymanchem.com). The assay utilized microtiter plates coated with anti-rabbit IgG. Plasma cortisol in the fish and an acetylcholinesterase (AChE)-labeled cortisol tracer competed for binding to the anti-cortisol rabbit antiserum. A standard curve was prepared with cortisol standards in place of plasma. After sufficient time for the binding of the rabbit antiserum to the plate-coated anti-rabbit IgG, Ellman’s reagent was added as the substrate for AChE. The absorbance of the ensuing yellow product was measured at 405 nm using a Multiskan EX plate reader (Thermo Scientific;
www.thermoscientific.com) and data were plotted using the associated software. A four-parameter logistic regression was used to determine cortisol concentrations in the fish, with the standard curves generating R
2 values over 0.97. The detection range for the assay as performed was 780 pg/mL to 300 ng/mL.
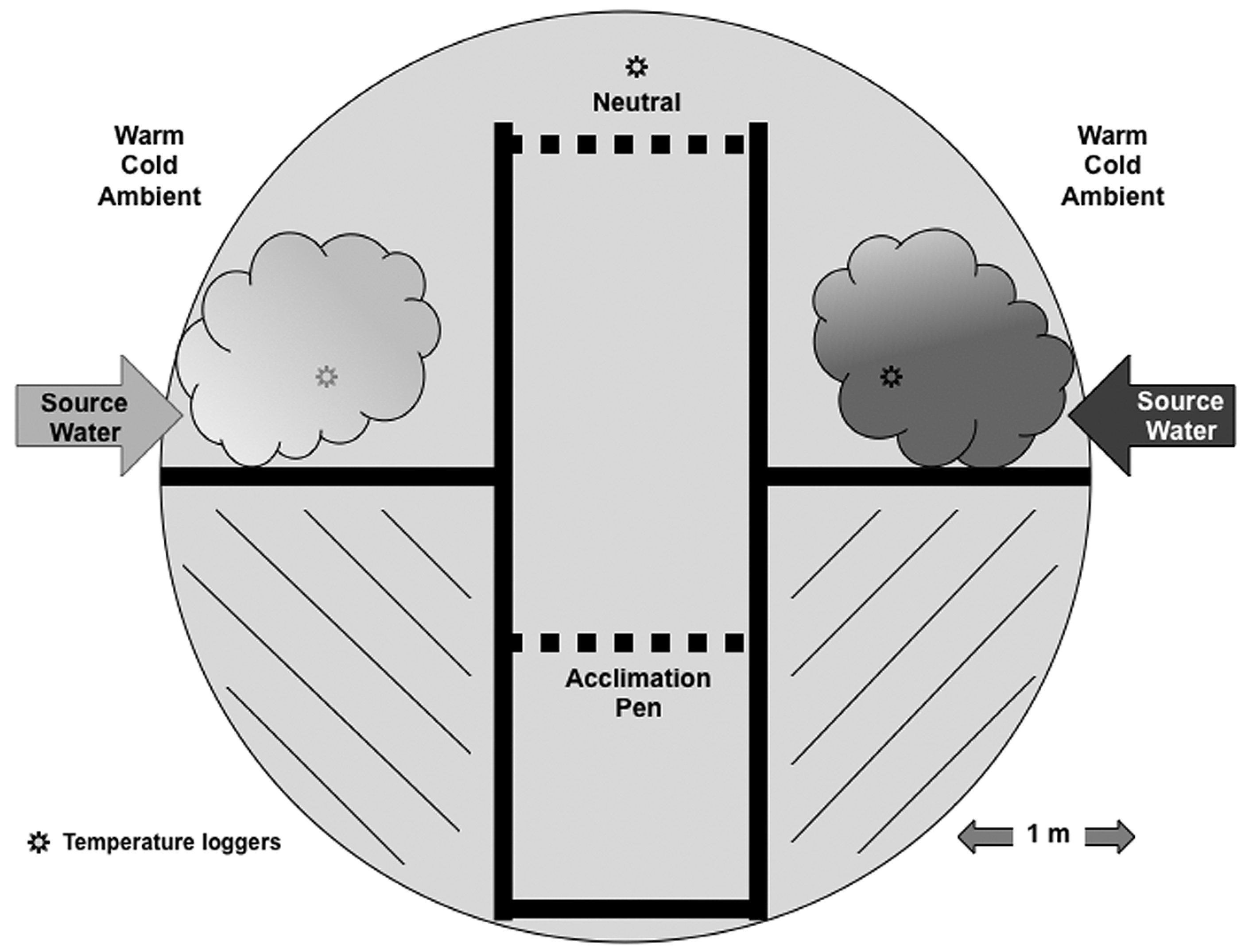
Choice behavior experiment. To assess whether
M. dolomieu behavior may mediate adverse effects of thermal cooling, a 20 m
2 thermal choice mace (
Figure 2) was constructed. For each of the 27 trials, a single mature
M. dolomieu (328 ± 68 mm total length; 554 ± 331 g wet weight) was held overnight in an acclimation pen (
Figure 2) at an ambient water temperature of approximately 17 ± 4 °C. The following morning, water inflow into the two side chambers (randomized daily to avoid a directional bias) was established with outflows near the neutral zone. Water inflow consisted of either cold (2 ± 1 °C) or warm (34 ± 2 °C) water. After mixing inside the side chambers, chamber temperatures averaged 4 ± 2 °C for the cold chamber and 30 ± 3 °C for the warm chamber (temperature was monitored continuously with Hobo temperature loggers, Onset;
www.onsetcomp.com). The locations of warm and cold chambers for each trial were randomized to avoid locational bias. One hour after flow was established to allow for the temperature gradient to become established in the choice maze, both gates (
Figure 2) were opened, and the fish was allowed to enter the choice maze (which usually occurred within a few minutes of the gate opening). Once the fish had passed the 2nd gate and entered the neutral area, the 2nd gate was lowered, preventing the fish from re-entering the channel. A digital video camera positioned above the choice maze recorded the swimming behavior of the fish and allowed for later quantification of the location of the fish throughout the trial. Each trial lasted approximately 180 min from the time the fish had entered the neutral area, and the 2nd gate was closed. In addition to 14 trials with a choice of cold or warm water, we also conducted six trials in which one chamber received cold water while the second chamber was maintained at ambient (acclimation) water temperature, and seven trials with an ambient and a warm chamber in the choice maze. Lastly, six trials were conducted without altering water temperature to ensure that no locational biases existed in the maze. Each fish was used only once in the experiment.
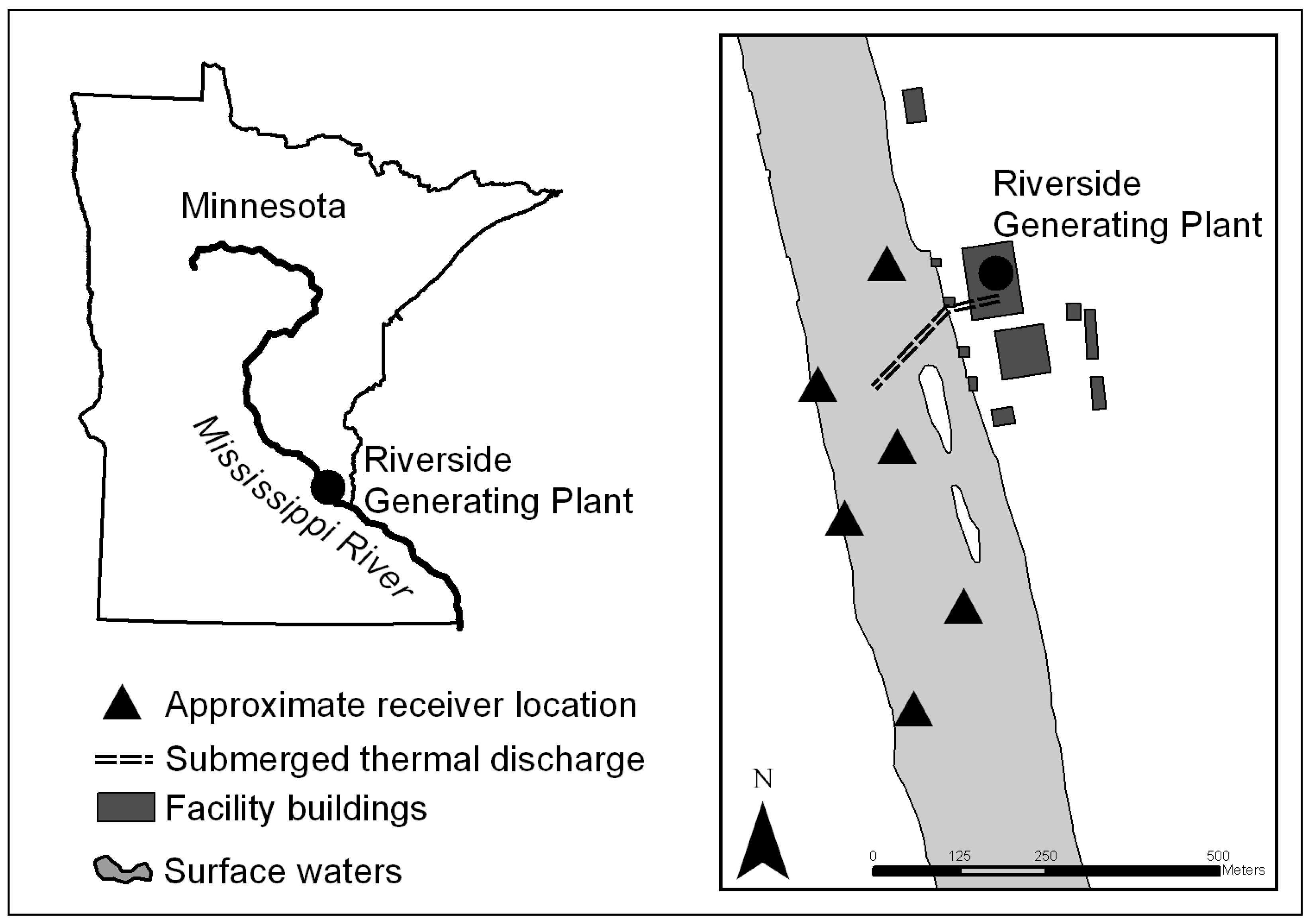
Telemetry. The physiological response of
M. dolomieu to an intermittent thermal discharge was assessed at the Riverside Generating Plant located on the Upper Mississippi River (
Figure 3). This facility underwent a complete repowering beginning in 2005, replacing the existing coal-fired units with combine-cycle natural gas-fired power generation in 2009. Cooling water is returned to the river through an approximately 183 m long, 2.1 m diameter submerged single port discharge to the center of the river. There are no tempering pumps at this facility and no permanent cooling towers; thus, there are no diversion measures available to reduce the temperature of the heated water before it reaches the river.
In November, 14
M. dolomieu (337 ± 19 mm total length; 522 ± 103 g body weight; Table 6) where captured by boat electro-shocking in the Mississippi River near the facility. After capture, fish were placed in an aerated induction tank containing water and 40 mg/L MS-222 until anesthesia and loss of equilibrium were induced. Fish were placed ventral side up on a foam surgical platform and a 20 mm incision was made slightly lateral to the ventral midline, just posterior to the pelvic girdle such that the acoustic transmitter (Vemco, V13T-1H;
www.vemco.com; 13 mm × 36 mm, 0° to 40 °C, 120 s nominal ping rate) could be placed intraperitoneally into the fish. Incisions were closed with 2 to 3 independent sutures made with nonabsorbable monofilament material [
18,
19,
20]. During the surgeries, individuals also received a maintenance dose of anesthetic to the gills supplied from the induction tank. Once completed, fish were returned to 756 L flow-through recovery tanks, held overnight for observation, and released at the terminus of the thermal mixing zone the following morning.
The monitoring system consisted of six omnidirectional acoustic hydrophones (VR2W, VEMCO) moored underwater in a grid-like pattern such that a “listening area” covered the regions above and below the thermal discharge (
Figure 3). Individual fish presence and core body temperature were monitored in the vicinity of the power plant’s thermal effluent from November to March when the hydrophones had to be removed due to impending spring flooding. When a single tag transmission was detected by three or more receivers, a position could be calculated by means of timed-difference-of-arrival via a proprietary post-processing method conducted by the manufacturer. However, following the field season, we assessed that the reliability of the system [
21] and proportional yield (percent of tag transmissions that result in a calculated position) was lower than 5% for the entire study (likely due to the sharp thermocline along the thermal effluent/river water edge). Therefore, this study focused strictly on the presence/absence and temperature of individuals when within the listening area.
Statistical Analysis. All statistical analyses were performed in Graphpad Prism for Macintosh (
www.graphpad.com). A Fisher’s exact test was used to assess differences in survival among fish cohorts following a laboratory-induced cold shock. The assumption of normality for cold shock and behavioral data sets were tested with the Kolmogorov–Smirnov test for normality prior to any additional analysis. As most data did not meet standards of homogeneity, data were analyzed using either a Mann–Whitney U test or a Kruskal–Wallis analysis followed by a Dunn’s post hoc test. A probability of
p < 0.05 was set as the level of significance for all comparisons.
4. Discussion
In this study, controlled cold shocks, behavioral trials, and telemetry were used to test the hypotheses that (i) thermal effluents will attract fish quickly and (ii) that the subsequent rapid fluctuation of the effluent temperature will induce stress and reduce survival in resident fish populations. The cold shock experiments resulted in significant mortality only when the final holding temperatures were <5 °C at the end of the cold shock. In contrast, the rate of temperature decline was only of secondary importance in predicting mortality. This second finding is counter-intuitive and not supported by previous cold shock effect models [
4]. However, it is noteworthy that both rates of temperature decline that were used in the current study (~4 °C and ~8 °C) are rapid when compared to other studies (e.g., [
22,
23]) or natural conditions and should not be extrapolated to possible effects observed in a controlled multi-day decline in water temperature. Furthermore, fish size did not appear to be a mitigating factor in predicting the likelihood of mortality to occur in association with any cold shock thermal profile. Similarly, the age of the fish (juvenile
L. macrochirus vs. adult minnows,
L. macrochirus, and
M. dolomieu) did not modulate survival in association with any cold shock thermal profile. The hematological and biochemical parameters measured in the fish in this study did not provide a clear trend in the responses to cold shock, which was surprising given the intimate connection between metabolic rates, stress (as induced by a sudden and dramatic change in temperature), and immunological response which has been reported for fish exposed to rising water temperatures (e.g., [
2,
3,
24]). However, all three species used in this study could be considered “hardy” and acclimated to great seasonal temperature fluctuations while species already considered more vulnerable (i.e., endangered/threatened) may be more susceptible to mortality. This caveat should be explored especially in circumstances where listed species may be residing in a thermal mixing zone downstream of effluents that lack thermal stability.
When given the choice between areas of cold and warm water, fish moved frequently back and forth between thermal areas and spent a similar length of time and total amount of time at the extremes of the temperature gradient. The continuous movement between all three areas may indicate a temperature-regulating behavior (avoiding warming/cooling of core temp). Although core body temperature was not recorded for these fish (to avoid the stress associated with the implantation surgery) this behavior was seen consistently across all fish used in the behavioral experiments. Given the mean amount of time each fish spent continuously in any thermal area, it is unlikely that core body temperature would have warmed substantially. It is important to note that these behavioral responses relate to short term changes in local water temperature as seen in the cycling at our field site. In contrast, established thermal plumes that persist for weeks, months, or even years have been shown to attract fish which will reside nearly continuously or at least seasonally in these plumes [
5,
9,
16,
25].
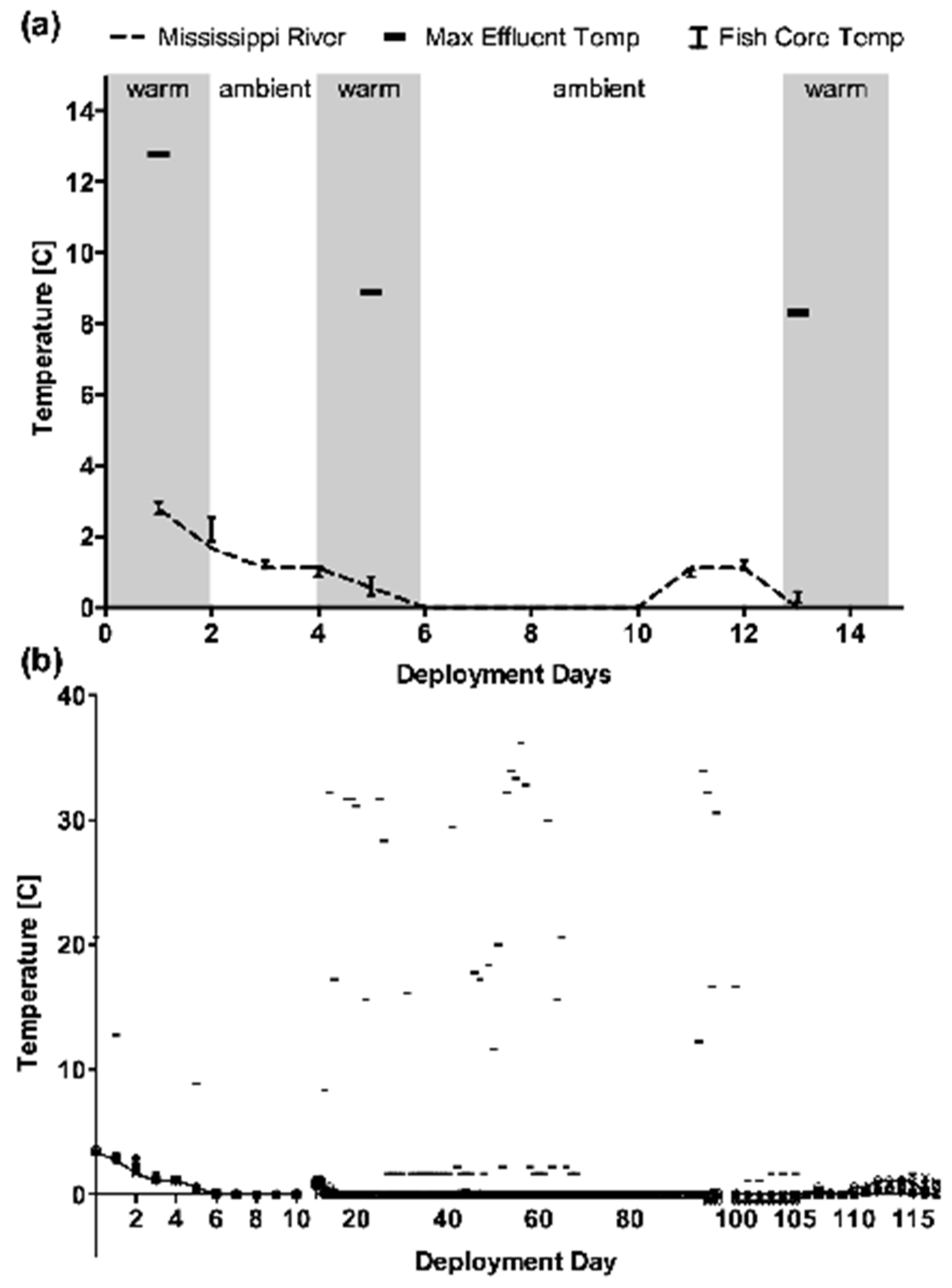
The assumption that fish near intermittent thermal plumes engage in a temperature-regulating behavior was supported by results obtained through telemetry assessments of resident fish near an intermittent thermal plume in the Mississippi River. Micropterus dolomieu were tracked for over three months to assess their presence and core body temperature on days with and without thermal effluent release. None of the fish tagged exhibited core body temperatures more than 2 °C above ambient river temperatures (as defined by water temperatures measured upstream of the thermal effluent discharge) even when the effluent temperature reached 35 °C at its discharge point. Although a poor proportional yield (likely as a result of sharp thermal profiles and the reflection of signals from moving ice on the river) prevented the accurate positional tracking of the resident M. dolomieu, their continued presence in the vicinity of the effluent discharge site and lack of body core temperature response to thermal discharge strongly suggests that fish either avoid the discharge plume during the 10–20 h of thermal discharge or spent only brief periods of time in the thermal plume, thus self-regulating their body temperature similar to the behavior observed in the choice maze.
The laboratory studies corroborate the hypothesis that rapid cooling, especially below 5 °C, can harm fish that are physiologically adjusted to higher temperatures. However, the behavioral trials and telemetry assessment lead us to reject the hypothesis that fish are attracted quickly to thermal effluents and, therefore, will adjust physiologically to higher temperatures. The combination of these findings provides aquatic resource managers and power plant operators with multiple avenues to manage thermal effluent and mitigate any associated risks to resident fish populations. The laboratory cold shock experiments across species, sizes, and ontogenetic stages suggest that the cessation of thermal effluent is hazardous to resident fish populations when ambient water temperatures drop below 5 °C. Furthermore, in large riverine systems, where resident fish populations can self-regulate their body temperature through swimming behaviors, cold shock, as the result of the off cycling of thermal effluents, may be mitigated by fish behavior. However, in small riverine systems, lakes, or aquatic environments in which fish movement is restricted, including confined discharge canals, the rapid on-and-off cycling of thermal effluents may have detrimental effects on resident fish populations.
The conclusions reached through these experiments need to be evaluated in the context of the species and conditions we assessed. All three species, L. macrochirus, P. promelas, and M. dolomieu are considered “hardy” species and are found across a wide range of ambient water temperatures. Furthermore, these results apply only to rapid cycling between warm and cold water temperatures; if a warm water plume persists for longer periods of time, as observed in base-load nuclear or coal-fire thermal effluents, the physiological response of the fish to a rapid cool-down of effluent (an emergency shutdown for example) may be quite different than that observed in the current study. Consequently, further studies are needed to assess and mitigate the effects of cold shock from the rapid operational cycling process that is generally indicative of combined-cycle natural gas power generation stations.




 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}