
Taxonomic Structure of Planktonic Protist Communities in Saline and Hypersaline Continental Waters Revealed by Metabarcoding
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Collection
2.2. eDNA Extraction, Amplification and 18S rDNA Library Preparation
2.3. eDNA Analysis
2.4. Phylogenetic Analysis of Centrohelids’ ASVs
2.5. Statistical Analysis
3. Results
3.1. Sequence Data Overview
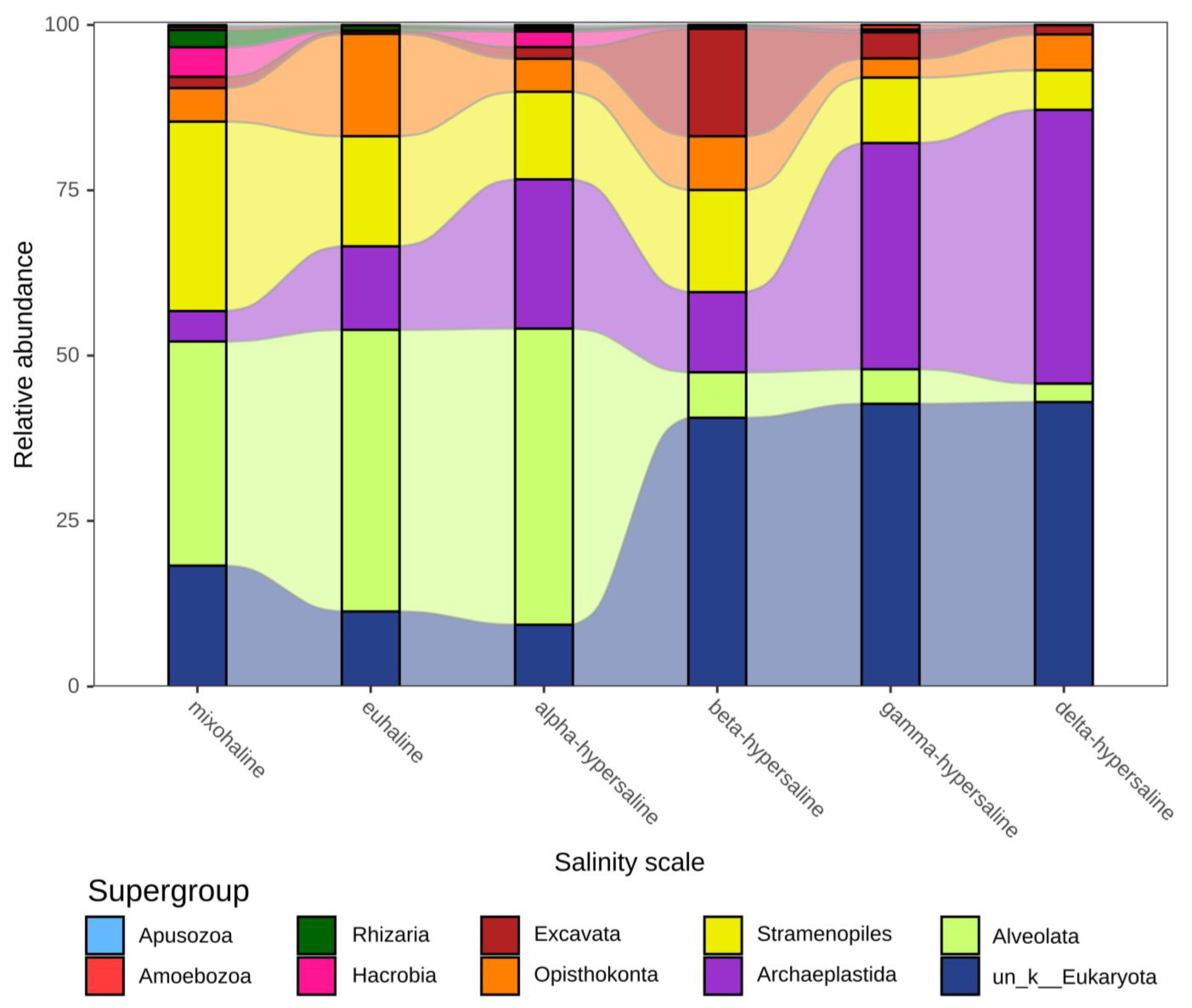
3.2. Taxonomic Composition and Distribution of Protists Communities
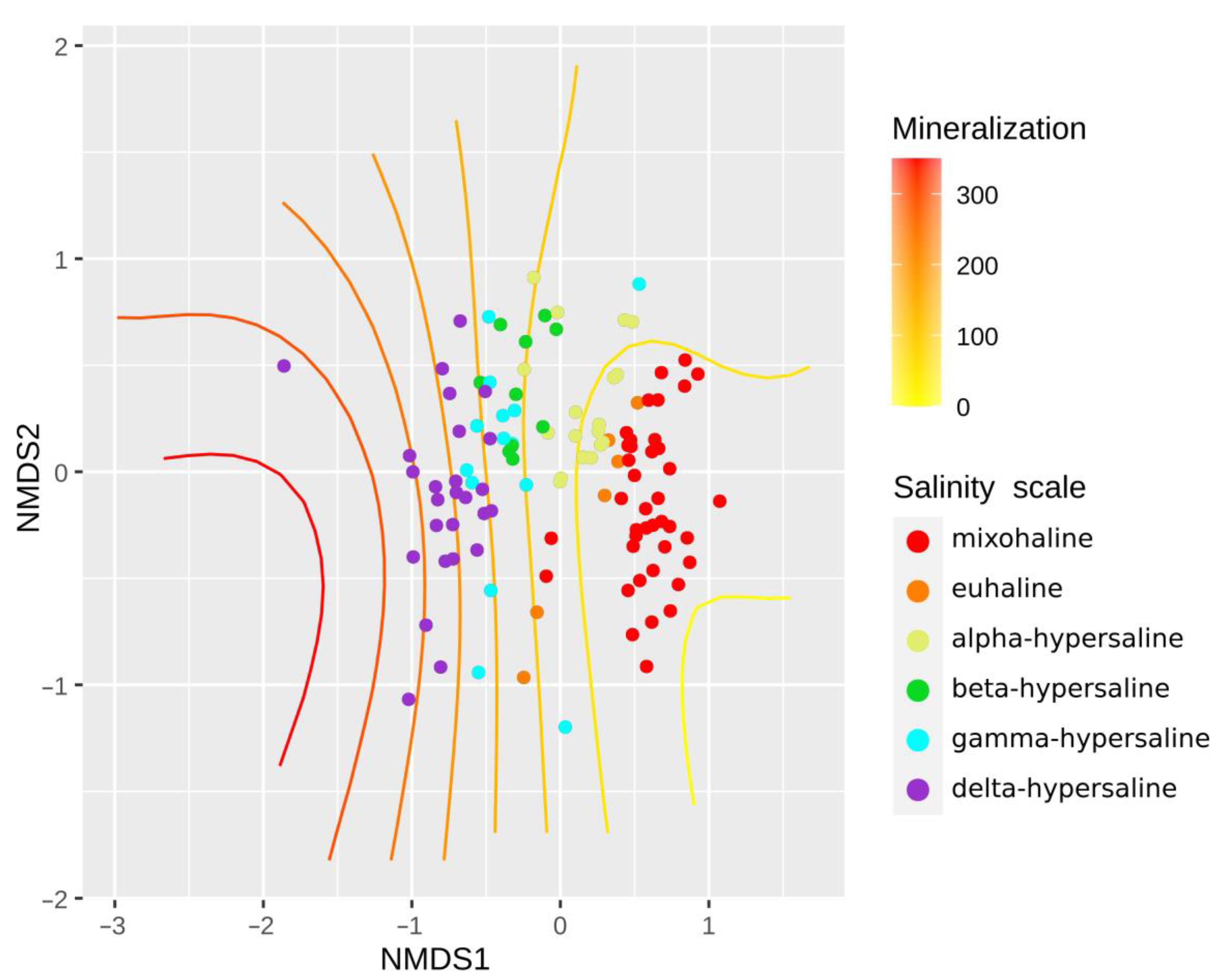
3.3. Protist Diversity and Salinity
3.4. Alpha- and Beta-Diversity Metrics
3.5. Centohelid Taxonomy and Diversity
3.6. Phylogenetic Position of Revealed ASVs and Marine/Freshwater Divergence of Centrohelids
4. Discussion
4.1. High-Throughput Sequencing and Promise in Biodiversity Studies
4.2. Protist Communities and Their Relationships with Salinity and Geographic Location
4.3. Salinity Tolerance and Marine/Freshwater Divergence in Centrohelids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rothschild, L.J.; Mancinelli, R.L. Life in Extreme Environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, A.K.; Allers, T.; DiRuggiero, J. Snap Shot: Microbial Extremophiles. Cell 2020, 180, 818–818.e1. [Google Scholar] [CrossRef] [PubMed]
- Post, F.J.; Borowitzka, L.J.; Borowitzka, M.A.; Mackay, B.; Moulton, T. The Protozoa of a Western Australian Hypersaline Lagoon. Hydrobiologia 1983, 105, 95–113. [Google Scholar] [CrossRef]
- Harding, T.; Simpson, A.G.B. Recent Advances in Halophilic Protozoa Research. J. Eukaryot. Microbiol. 2018, 65, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.-S.; Huang, L.-N. Microbial Diversity in Extreme Environments. Nat. Rev. Microbiol. 2022, 20, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Triadó-Margarit, X.; Casamayor, E.O. High Genetic Diversity and Novelty in Planktonic Protists Inhabiting Inland and Coastal High Salinity Water Bodies. FEMS Microbiol. Ecol. 2013, 85, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Menéndez-Serra, M.; Triadó-Margarit, X.; Casamayor, E.O. Ecological and Metabolic Thresholds in the Bacterial, Protist, and Fungal Microbiome of Ephemeral Saline Lakes (Monegros Desert, Spain). Microb. Ecol. 2021, 82, 885–896. [Google Scholar] [CrossRef]
- Wang, J.; Wang, F.; Chu, L.; Wang, H.; Zhong, Z.; Liu, Z.; Gao, J.; Duan, H. High Genetic Diversity and Novelty in Eukaryotic Plankton Assemblages Inhabiting Saline Lakes in the Qaidam Basin. PLoS ONE 2014, 9, e112812. [Google Scholar] [CrossRef] [Green Version]
- Rich, V.I.; Maier, R.M. Aquatic Environments. In Environmental Microbiology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 111–138. ISBN 978-0-12-394626-3. [Google Scholar]
- Williams, W.D. Environmental Threats to Salt Lakes and the Likely Status of Inland Saline Ecosystems in 2025. Environ. Conserv. 2002, 29, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Shadrin, N.V.; Anufriieva, E.V.; Belyakov, V.P.; Bazhora, A.I. Chironomidae Larvae in Hypersaline Waters of the Crimea: Diversity, Distribution, Abundance and Production. Eur. Zool. J. 2017, 84, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Por, F.D. A Classification of Hypersaline Waters, Based on Trophic Criteria. Mar. Ecol. 1980, 1, 121–131. [Google Scholar] [CrossRef]
- Venice System (1958) Symposium on the Classification of Brackish Waters, Venice. Arch. Oceanogr. Limnol. 1958, 11, 1–248.
- Williams, W.D. The Largest, Highest and Lowest Lakes of the World: Saline Lakes. SIL Proc. 1922–2010 1996, 26, 61–79. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Anufrieva, E.V.; Shadrina, S.N. Brief Review of Phototrophs of Hypersaline Lakes and Lagoons of Crimea. Diversity, Ecological Role, Possibilities of Use. Mar. Biol. J. 2017, 2, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Waiser, M.J.; Robarts, R.D. Saline Inland Waters. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Oxford, UK, 2009; pp. 634–644. ISBN 978-0-12-370626-3. [Google Scholar]
- Saccò, M.; White, N.E.; Harrod, C.; Salazar, G.; Aguilar, P.; Cubillos, C.F.; Meredith, K.; Baxter, B.K.; Oren, A.; Anufriieva, E.; et al. Salt to Conserve: A Review on the Ecology and Preservation of Hypersaline Ecosystems. Biol. Rev. 2021, 96, 2828–2850. [Google Scholar] [CrossRef]
- Casamayor, E.O.; Triadó-Margarit, X.; Castañeda, C. Microbial Biodiversity in Saline Shallow Lakes of the Monegros Desert, Spain. FEMS Microbiol. Ecol. 2013, 85, 503–518. [Google Scholar] [CrossRef] [Green Version]
- Dillon, J.G.; McMath, L.M.; Trout, A.L. Seasonal Changes in Bacterial Diversity in the Salton Sea. Hydrobiologia 2009, 632, 49–64. [Google Scholar] [CrossRef]
- Parnell, J.J.; Rompato, G.; Latta, L.C.I.V.; Pfrender, M.E.; Van Nostrand, J.D.; He, Z.; Zhou, J.; Andersen, G.; Champine, P.; Ganesan, B.; et al. Functional Biogeography as Evidence of Gene Transfer in Hypersaline Microbial Communities. PLoS ONE 2010, 5, e12919. [Google Scholar] [CrossRef] [Green Version]
- Pagaling, E.; Wang, H.; Venables, M.; Wallace, A.; Grant, W.D.; Cowan, D.A.; Jones, B.E.; Ma, Y.; Ventosa, A.; Heaphy, S. Microbial Biogeography of Six Salt Lakes in Inner Mongolia, China, and a Salt Lake in Argentina. Appl. Environ. Microbiol. 2009, 75, 5750–5760. [Google Scholar] [CrossRef] [Green Version]
- Makhdoumi-Kakhki, A.; Amoozegar, M.A.; Kazemi, B.; Pašić, L.; Ventosa, A. Prokaryotic Diversity in Aran-Bidgol Salt Lake, the Largest Hypersaline Playa in Iran. Microbes Environ. 2012, 27, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Narasingarao, P.; Podell, S.; Ugalde, J.A.; Brochier-Armanet, C.; Emerson, J.B.; Brocks, J.J.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. De novo Metagenomic Assembly Reveals Abundant Novel Major Lineage of Archaea in Hypersaline Microbial Communities. ISME J. 2012, 6, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filker, S.; Gimmler, A.; Dunthorn, M.; Mahé, F.; Stoeck, T. Deep Sequencing Uncovers Protistan Plankton Diversity in the Portuguese Ria Formosa Solar Saltern Ponds. Extremophiles 2015, 19, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Demergasso, C.; Escudero, L.; Casamayor, E.O.; Chong, G.; Balagué, V.; Pedrós-Alió, C. Novelty and Spatio-Temporal Heterogeneity in the Bacterial Diversity of Hypersaline Lake Tebenquiche (Salar de Atacama). Extremophiles 2008, 12, 491–504. [Google Scholar] [CrossRef]
- Guglielmin, M.; Azzaro, M.; Buzzini, P.; Battistel, D.; Roman, M.; Ponti, S.; Turchetti, B.; Sannino, C.; Borruso, L.; Papale, M.; et al. A Possible Unique Ecosystem in the Endoglacial Hypersaline Brines in Antarctica. Sci. Rep. 2023, 13, 177. [Google Scholar] [CrossRef] [PubMed]
- Podell, S.; Ugalde, J.A.; Narasingarao, P.; Banfield, J.F.; Heidelberg, K.B.; Allen, E.E. Assembly-Driven Community Genomics of a Hypersaline Microbial Ecosystem. PLoS ONE 2013, 8, e61692. [Google Scholar] [CrossRef] [Green Version]
- Podell, S.; Emerson, J.B.; Jones, C.M.; Ugalde, J.A.; Welch, S.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. Seasonal Fluctuations in Ionic Concentrations Drive Microbial Succession in a Hypersaline Lake Community. ISME J. 2014, 8, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Naghoni, A.; Emtiazi, G.; Amoozegar, M.A.; Cretoiu, M.S.; Stal, L.J.; Etemadifar, Z.; Shahzadeh Fazeli, S.A.; Bolhuis, H. Microbial Diversity in the Hypersaline Lake Meyghan, Iran. Sci. Rep. 2017, 7, 11522. [Google Scholar] [CrossRef] [Green Version]
- Kheiri, R.; Mehrshad, M.; Pourbabaee, A.A.; Ventosa, A.; Amoozegar, M.A. Hypersaline Lake Urmia: A Potential Hotspot for Microbial Genomic Variation. Sci. Rep. 2023, 13, 374. [Google Scholar] [CrossRef]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.-B.; Sánchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. Prokaryotic Taxonomic and Metabolic Diversity of an Intermediate Salinity Hypersaline Habitat Assessed by Metagenomics. FEMS Microbiol. Ecol. 2014, 88, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Ghai, R.; Pašić, L.; Fernández, A.B.; Martin-Cuadrado, A.-B.; Mizuno, C.M.; McMahon, K.D.; Papke, R.T.; Stepanauskas, R.; Rodriguez-Brito, B.; Rohwer, F.; et al. New Abundant Microbial Groups in Aquatic Hypersaline Environments. Sci. Rep. 2011, 1, 135. [Google Scholar] [CrossRef] [Green Version]
- DeMaere, M.Z.; Williams, T.J.; Allen, M.A.; Brown, M.V.; Gibson, J.A.E.; Rich, J.; Lauro, F.M.; Dyall-Smith, M.; Davenport, K.W.; Woyke, T.; et al. High Level of Intergenera Gene Exchange Shapes the Evolution of Haloarchaea in an Isolated Antarctic Lake. Proc. Natl. Acad. Sci. USA 2013, 110, 16939–16944. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Knight, R. Global Patterns in Bacterial Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [Green Version]
- Auguet, J.-C.; Barberan, A.; Casamayor, E.O. Global Ecological Patterns in Uncultured Archaea. ISME J. 2010, 4, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Nolte, V.; Pandey, R.V.; Jost, S.; Medinger, R.; Ottenwälder, B.; Boenigk, J.; Schlötterer, C. Contrasting Seasonal Niche Separation Between Rare and Abundant Taxa Conceals the Extent of Protist Diversity: High Seasonal Protist Abundance Turnover. Mol. Ecol. 2010, 19, 2908–2915. [Google Scholar] [CrossRef] [Green Version]
- Scheckenbach, F.; Hausmann, K.; Wylezich, C.; Weitere, M.; Arndt, H. Large-Scale Patterns in Biodiversity of Microbial Eukaryotes from the Abyssal Sea Floor. Proc. Natl. Acad. Sci. USA 2010, 107, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Grey, E.K.; Bernatchez, L.; Cassey, P.; Deiner, K.; Deveney, M.; Howland, K.L.; Lacoursière-Roussel, A.; Leong, S.C.Y.; Li, Y.; Olds, B.; et al. Effects of Sampling Effort on Biodiversity Patterns Estimated from Environmental DNA Metabarcoding Surveys. Sci. Rep. 2018, 8, 8843. [Google Scholar] [CrossRef] [Green Version]
- Medinger, R.; Nolte, V.; Pandey, R.V.; Jost, S.; Ottenwälder, B.; Schlötterer, C.; Boenigk, J. Diversity in a Hidden World: Potential and Limitation of Next-Generation Sequencing for Surveys of Molecular Diversity of Eukaryotic Microorganisms. Mol. Ecol. 2010, 19, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Pawlowski, J.; Christen, R.; Lecroq, B.; Bachar, D.; Shahbazkia, H.R.; Amaral-Zettler, L.; Guillou, L. Eukaryotic Richness in the Abyss: Insights from Pyrotag Sequencing. PLoS ONE 2011, 6, e18169. [Google Scholar] [CrossRef]
- Edgcomb, V.; Bernhard, J. Heterotrophic Protists in Hypersaline Microbial Mats and Deep Hypersaline Basin Water Columns. Life 2013, 3, 346–362. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.B.; Jeong, D.H.; Cho, B.C.; Park, J.S. The Diversity Patterns of Rare to Abundant Microbial Eukaryotes Across a Broad Range of Salinities in a Solar Saltern. Microb. Ecol. 2022, 84, 1103–1121. [Google Scholar] [CrossRef]
- Alexander, E.; Stock, A.; Breiner, H.-W.; Behnke, A.; Bunge, J.; Yakimov, M.M.; Stoeck, T. Microbial Eukaryotes in the Hypersaline Anoxic L’Atalante Deep-Sea Basin. Environ. Microbiol. 2009, 11, 360–381. [Google Scholar] [CrossRef] [PubMed]
- Filker, S.; Forster, D.; Weinisch, L.; Mora-Ruiz, M.; González, B.; Farías, M.E.; Rosselló-Móra, R.; Stoeck, T. Transition Boundaries for Protistan Species Turnover in Hypersaline Waters of Different Biogeographic Regions. Environ. Microbiol. 2017, 19, 3186–3200. [Google Scholar] [CrossRef] [PubMed]
- Gerasimova, E.A.; Plotnikov, A.O.; Khlopko, Y.A.; Zlatogursky, V.V. Multiple Euryhaline Lineages of Centrohelids (Haptista: Centroplasthelida) in Inland Saline Waters Revealed with Metabarcoding. J. Eukaryot. Microbiol. 2020, 67, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; del Campo, J.; Wylezich, C.; Massana, R.; Jürgens, K. Unveiling Trophic Functions of Uncultured Protist Taxa by Incubation Experiments in the Brackish Baltic Sea. PLoS ONE 2012, 7, e41970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrischev, V.P. Salt dome morphostructures in the Southern Priuralye. Geomorfologiya 2010, 1, 86–94. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference Database (PR2): A Catalog of Unicellular Eukaryote Small Sub-Unit RRNA Sequences with Curated Taxonomy. Nucleic Acids Res. 2012, 41, D597–D604. [Google Scholar] [CrossRef] [Green Version]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukes, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [Green Version]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A.G.B. The New Tree of Eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Use R; Springer: Cham, Switzerland, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Xu, S.; Zhan, L.; Tang, W.; Wang, Q.; Dai, Z.; Zhou, L.; Feng, T.; Chen, M.; Wu, T.; Hu, E.; et al. Microbiota Process: A Comprehensive R Package for Deep Mining Microbiome. Innovation 2023, 4, 100388. [Google Scholar] [CrossRef]
- Lahti, L.; Shetty, S. Microbiome R Package. Available online: http://microbiome.github.io (accessed on 18 May 2023).
- Oren, A. Life in Hypersaline Environments. In Their World: A Diversity of Microbial Environments; Hurst, C.J., Ed.; Advances in Environmental Microbiology; Springer: Cham, Switzerland, 2016; Volume 1, pp. 301–339. ISBN 978-3-319-28069-1. [Google Scholar]
- Mylnikov, A.P. A New Flagellate Percolomonas laustris sp. n. (Excavata, Percolozoa) from an Inland Saline Lake (Southeastern European Russia). Biol. Bull. Russ. Acad. Sci. 2016, 43, 587–594. [Google Scholar] [CrossRef]
- Dovgal, I.V.; Shadrin, N.V.; Gaponova, L.P. New Findings of Halobiont Ciliates (Ciliophora). Vestn. Zool. 2006, 40, 462. (In Russian) [Google Scholar]
- Pavlovskya, T.V.; Prazukin, A.V.; Shadrin, N.V. Seasonal Phenomena in Infusoria Community in Hypersaline Lake Khersonesskoye (Crimea). Mar. Ecol. J. 2009, 2, 53–63. (In Russian) [Google Scholar]
- Nevrova, E.L.; Shadrin, N.V. Bottom Diatom Algae of the Saline Lakes of Crimea. Mar. Ecol. J. 2005, 4, 61–71. (In Russian) [Google Scholar]
- Plotnikov, A.O.; Selivanova, E.A.; Nemtseva, N.V. Species Diversity of Heterotrophic Flagellates in Saline Sol’-Iletsk Lakes. Izv. PGU Im. V.G. Belinsky 2011, 25, 548–557. (In Russian) [Google Scholar]
- Senicheva, M.I.; Gubelit, Y.I.; Prazukhin, A.V.; Shadrin, N.V. Phytoplankton of Crimean Hypersaline Lakes. In Black Sea Microalgae: Problems of Biodiversity Conservation and Biotechnological Use; Tokarev, Y.N., Finenko, Z.Z., Shadrin, N.V., Eds.; EKOSI-Gidrofizika: Sevastopol, Russia, 2008; pp. 93–100. ISBN 978-966-02-5118-2. (In Russian) [Google Scholar]
- Zagorodnaya, Y.A.; Batogoeva, E.A.; Shadrin, N.V. Long-term Transformation of Zooplankton in the Hypersaline Lake Bakalskoe (Crimea) Under Salinity Fluctuations. Mar. Ecol. J. 2008, 7, 41–50. [Google Scholar]
- Prokina, K.I. Heterotrophic Flagellates from the Saline Lakes of the Crimean Peninsula. Ecosyst. Transform. 2020, 3, 3–10. [Google Scholar] [CrossRef]
- Plotnikov, A.O.; Ermolenko, E.A.; Nikiyan, H.N.; Vasilchenko, A.S. Study of centrohelid Heliozoa morphology by atomic force microscopy. Zool. J. 2013, 92, 955–961. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; Shadrin, N.V.; Shadrina, S.N. History of Research on Biodiversity in Crimean Hypersaline Waters. Arid. Ecosyst. 2017, 7, 52–58. [Google Scholar] [CrossRef]
- Nemtseva, N.V.; Selivanova, E.A.; Yatsenko-Stepanova, T.N.; Ignatenko, M.E. The structure of algoplankton in the Sol-Iletsk lakes with different levels of salinity. Izv. PGU Im. V.G. Belinsky 2011, 25, 535–541. [Google Scholar]
- Nemtseva, N.V.; Selivanova, E.A.; Ignatenko, M.E.; Sharapova, N.V. Characterization of a new strain of Dunaliella salina (Chlorophyta) and assessment of its cultivation parameters. Physiol. Rast. 2013, 60, 561–568. [Google Scholar] [CrossRef]
- Plotnikov, A.O.; Ermolenko, E.A. Centrohelid Heliozoa (Chromista, Hacrobia) of Southern Cis-Ural Region. Biol. Bull. Russ. Acad. Sci. 2015, 42, 683–695. [Google Scholar] [CrossRef]
- Plotnikov, A.O.; Gerasimova, E.A. Heliozoa (Centrohelea, Haptista, Hacrobia) of saline and brackish water bodies and watercourses of Russia. Inland Water Biol. 2017, 10, 121–129. [Google Scholar] [CrossRef]
- El-Shabrawy, G.; Anufriieva, E.; Shadrin, N. Tintinnina (Ciliophora) and Foraminifera in plankton of hypersaline Lagoon Bardawil (Egypt): Spatial and temporal variability. Turk. J. Zool. 2018, 42, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Zhao, Y.; Wang, Q.; Zheng, M.; Wei, J.; Wang, S. The community structure and seasonal dynamics of plankton in Bange Lake, northern Tibet, China. Chin. J. Ocean. Limnol. 2016, 34, 1143–1157. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, Y.Y.; Wang, Q.H.; Zheng, M.P.; Zhang, P.; He, Z.H. Spatial and temporal patterns of plankton assemblage structure in a high altitude saline lake, Namuka Co in Northern Tibet, China. Clean Soil Air Water 2010, 38, 960–968. [Google Scholar] [CrossRef]
- Sonntag, B.; Posch, T.; Klammer, S.; Teubner, K.; Psenner, R. Phagotrophic ciliates and flagellates in an oligotrophic, deep, alpine lake: Contrasting variability with seasons and depths. Aquat. Microb. Ecol. 2006, 43, 193–207. [Google Scholar] [CrossRef]
- Gran-Stadniczeñko, S.; Egge, E.; Hostyeva, V.; Logares, R.; Eikrem, W.; Edvardsen, B. Protist diversity and seasonal dynamics in skagerrak plankton communities as revealed by metabarcoding and microscopy. J. Eukaryot. Microbiol. 2019, 66, 494–513. [Google Scholar] [CrossRef] [Green Version]
- Khromechek, E.B.; Barkhatov, Y.V.; Rogozin, D.Y. Community structure and vertical distribution of planktonic ciliates in the saline meromictic lake Shira during breakdown of meromixis. Ecohydrol. Hydrobiol. 2021, 21, 142–152. [Google Scholar] [CrossRef]
- Lepère, C.; Domaizon, I.; Hugoni, M.; Vellet, A.; Debroas, D. Diversity and Dynamics of Active Small Microbial Eukaryotes in the Anoxic Zone of a Freshwater Meromictic Lake (Pavin, France). Front. Microbiol. 2016, 7, 130. [Google Scholar] [CrossRef] [Green Version]
- Edgcomb, V.P.; Pachiadaki, M. Ciliates along oxyclines of permanently stratified marine water columns. J. Eukaryot. Microbiol. 2014, 61, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Fermani, P.; Metz, S.; Balagué, V.; Descy, J.P.; Morana, C.; Logares, R.; Massana, R.; Sarmento, H. Microbial eukaryote assemblages and potential novel diversity in four tropical East African Great Lakes. FEMS Microbiol. Ecol. 2021, 97, fiab114. [Google Scholar] [CrossRef]
- Khromechek, E.B.; Barkhatov, Y.V.; Rogozin, D.Y. Densities and distribution of flagellates and ciliates in the chemocline of saline, meromictic Lake Shunet (Siberia, Russia). Aquat. Ecol. 2010, 44, 497–511. [Google Scholar] [CrossRef]
- Zlatogursky, V.V.; Gerasimova, E.A.; Drachko, D.; Klimov, V.I.; Shɨshkin, Y.; Plotnikov, A.O. Pinjata ruminata gen. et sp. n.—A new member of centrohelid family Yogsothothidae (Haptista: Centroplasthelida) from the brackish river. J. Eukaryot. Microbiol. 2019, 66, 862–868. [Google Scholar] [CrossRef]
- Gerasimova, E.A. Morphology and species composition of centrohelid heliozoans (Centroplasthelida) from the continental lowland river with salinity gradient. Inland Water Biol. 2021, 14, 652–661. [Google Scholar] [CrossRef]
- Gerasimova, E.A. Two new brackish-water species of centrohelid heliozoans (Haptista: Centroplasthelida), Choanocystis mylnikovi sp. n. and C. punctata sp. n., from Russia. Protistology 2022, 16, 10–20. [Google Scholar] [CrossRef]
- Gerasimova, E.A.; Radaykina, L.V.; Zagumyonnyi, D.G.; Tikhonenkov, D.V.; Drachko, D.; Zlatogursky, V.V. Morphology and spicules elemental composition of Marophrys nikolaevi spec. nov. (Haptista: Centroplasthelida). Eur. J. Protistol. 2022, 84, 125888. [Google Scholar] [CrossRef] [PubMed]
- Heidelberg, K.B.; Nelson, W.C.; Holm, J.B.; Eisenkolb, N.; Andrade, K.; Emerson, J.B. Characterization of Eukaryotic Microbial Diversity in Hypersaline Lake Tyrrell, Australia. Front. Microbiol. 2013, 4, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casamayor, E.O.; Massana, R.; Benlloch, S.; Øvreås, L.; Díez, B.; Goddard, V.J.; Gasol, J.M.; Joint, I.; Rodríguez-Valera, F.; Pedrós-Alió, C. Changes in Archaeal, Bacterial and Eukaryal Assemblages Along a Salinity Gradient by Comparison of Genetic Fingerprinting Methods in a Multipond Solar Saltern. Environ. Microbiol. 2002, 4, 338–348. [Google Scholar] [CrossRef]
- Pedrós-Alió, C.; Calderón-Paz, J.I.; MacLean, M.H.; Medina, G.; Marrasé, C.; Gasol, J.M.; Guixa-Boixereu, N. The Microbial Food Web Along Salinity Gradients. FEMS Microbiol. Ecol. 2000, 32, 143–155. [Google Scholar] [CrossRef]
- Elloumi, J.; Carrias, J.-F.; Ayadi, H.; Sime-Ngando, T.; Bouaïn, A. Communities Structure of the Planktonic Halophiles in the Solar Saltern of Sfax, Tunisia. Estuar. Coast. Shelf Sci. 2009, 81, 19–26. [Google Scholar] [CrossRef]
- Oren, A. The Ecology of Dunaliella in High-Salt Environments. J. Biol. Res. Thessalon. 2014, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. A Hundred Years of Dunaliella Research: 1905–2005. Aquat. Biosyst. 2005, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Glycerol Metabolism in Hypersaline Environments: Glycerol Metabolism. Environ. Microbiol. 2017, 19, 851–863. [Google Scholar] [CrossRef]
- Mikrjukov, K.A. Heliozoa as a Component of Marine Microbenthos. In Proceedings of the 3rd The European Congress of Protistology, Helsingor, Denmark, 25–30 July 1999; p. 49. [Google Scholar]
- Mikrjukov, K.A. Heliozoa As a Component of Marine Microbenthos: A Study of Heliozoa of the White Sea. Ophelia 2001, 54, 51–73. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; von der Heyden, S. Molecular Phylogeny, Scale Evolution and Taxonomy of Centrohelid Heliozoa. Mol. Phylogenet. Evol. 2007, 44, 1186–1203. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; Chao, E.E.-Y. Oxnerella micra sp. n. (Oxnerellidae fam. n.), a Tiny Naked Centrohelid, and the Diversity and Evolution of Heliozoa. Protist 2012, 163, 574–601. [Google Scholar] [CrossRef]
- Shɨshkin, Y.; Drachko, D.; Klimov, V.I.; Zlatogursky, V.V. Yogsothoth knorrus gen. n., sp. n. and Y. carteri sp. n. (Yogsothothidae fam. n., Haptista, Centroplasthelida), with Notes on Evolution and Systematics of Centrohelids. Protist 2018, 169, 682–696. [Google Scholar] [CrossRef]
- Shɨshkin-Skarð, Y.; Drachko, D.; Zlatogursky, V.V. Shedding Light on the Origin of Acanthocystidae: Ricksol blepharistes gen. n., sp. n. (Ricksolidae fam. n., Panacanthocystida, Centroplasthelida), with Notes on the Evolution of the Genera Acanthocystis, Ozanamia gen. n. (Ozanamiidae fam. n.), and “Heterophrys-like Organisms”. Org. Diver. Evol. 2023, 23, 263–274. [Google Scholar] [CrossRef]
- Zlatogursky, V.V.; Shishkin, Y.; Drachko, D.; Burki, F. The Long-Time Orphan Protist Meringosphaera mediterranea Lohmann, 1902 [1903] is a Centrohelid Heliozoan. J. Eukaryot. Microbiol. 2021, 68, e12860. [Google Scholar] [CrossRef]
- Gerasimova, E.A.; Plotnikov, A.O. New Freshwater Species of Centrohelids Acanthocystis lyra sp. nov. and Acanthocystis siemensmae sp. nov. (Haptista, Heliozoa, Centrohelea) from the South Urals, Russia. Acta Protozool. 2016, 55, 231–237. [Google Scholar] [CrossRef]
- Zlatogursky, V.V.; Gerasimova, E.A.; Plotnikov, A.O. A New Species of Centrohelid Heliozoan Acanthocystis amura n. sp. Isolated from Two Remote Locations in Russia. J. Eukaryot. Microbiol. 2017, 64, 434–439. [Google Scholar] [CrossRef]
- Zagumyonnyi, D.G.; Radaykina, L.V.; Tikhonenkov, D.V. Triangulopteris lacunata gen. et sp. nov. (Centroplasthelida), a New Centrohelid Heliozoan from Soil. Diversity 2021, 13, 658. [Google Scholar] [CrossRef]
- Zagumyonnyi, D.G.; Radaykina, L.V.; Keeling, P.J.; Tikhonenkov, D.V. Centrohelid Heliozoans of Ukraine with a Description of a New Genus and Species (Haptista: Centroplasthelida). Eur. J. Protistol. 2022, 86, 125916. [Google Scholar] [CrossRef]
- Ponizovsky, A.M. Salt Resources of the Crimea; Krym: Simferopol, Crimea, 1965; 163p. [Google Scholar]
- Selivanova, E.A.; Poshvina, D.V.; Khlopko, Y.A.; Gogoleva, N.E.; Plotnikov, A.O. Diversity of Prokaryotes in Planktonic Communities of Saline Sol-Iletsk Lakes (Orenburg Oblast, Russia). Microbiology 2018, 87, 569–582. [Google Scholar] [CrossRef]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerasimova, E.A.; Balkin, A.S.; Filonchikova, E.S.; Mindolina, Y.V.; Zagumyonnyi, D.G.; Tikhonenkov, D.V. Taxonomic Structure of Planktonic Protist Communities in Saline and Hypersaline Continental Waters Revealed by Metabarcoding. Water 2023, 15, 2008. https://doi.org/10.3390/w15112008
Gerasimova EA, Balkin AS, Filonchikova ES, Mindolina YV, Zagumyonnyi DG, Tikhonenkov DV. Taxonomic Structure of Planktonic Protist Communities in Saline and Hypersaline Continental Waters Revealed by Metabarcoding. Water. 2023; 15(11):2008. https://doi.org/10.3390/w15112008
Chicago/Turabian StyleGerasimova, Elena A., Alexander S. Balkin, Ekaterina S. Filonchikova, Yulia V. Mindolina, Dmitry G. Zagumyonnyi, and Denis V. Tikhonenkov. 2023. "Taxonomic Structure of Planktonic Protist Communities in Saline and Hypersaline Continental Waters Revealed by Metabarcoding" Water 15, no. 11: 2008. https://doi.org/10.3390/w15112008