
Spatial and Temporal Variations in Fish Assemblage in Feitsui Reservoir, in Northern Taiwan, from 2006–2020


Abstract
:1. Introduction
2. Materials and Methods
2.1. Environmental Parameters
2.2. Fish Sampling
2.3. Data Analysis
3. Results
3.1. Seasonal Environmental Variability
3.2. Fish Species Composition
3.3. Long-Term Variation in the Fish Assemblage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pool, T.K.; Olden, J.D. Taxonomic and functional homogenization of an endemic desert fish fauna. Divers. Distrib. 2012, 18, 366–376. [Google Scholar] [CrossRef]
- Clavero, M.; Hermoso, V. Reservoirs promote the taxonomic homogenization of fish communities within river basins. Biodivers. Conserv. 2011, 20, 41–57. [Google Scholar] [CrossRef]
- Pelicice, F.M.; Vitule, J.R.S.; Lima, D.P., Jr.; Orsi, M.L.; Agostinho, A.A. A Serious New Threat to Brazilian Freshwater Ecosystems: The Naturalization of Nonnative Fish by Decree. Conserv. Lett. 2014, 7, 55–60. [Google Scholar] [CrossRef]
- Nobile, A.B.; Freitas-Souza, D.; Lima, F.P.; Queiroz, J.; Bayona-Perez, I.L.; Carvalho, E.D.; Ramos, I.P. Damming and seasonality as modulators of fish community structure in a small tributary. Ecol. Freshw. Fish 2019, 28, 563–572. [Google Scholar] [CrossRef]
- Sá-Oliveira, J.C.; Angelini, R.; Isaac-Nahum, V.J. Population parameters of the fish fauna in a long-established Amazonian reservoir (Amapá, Brazil). J. Appl. Ichthyol. 2015, 31, 290–295. [Google Scholar] [CrossRef]
- Yang, Z.; Pan, X.; Hu, L.; Xu, W.; Jin, Y.; Zhao, N.; Yang, Q.; Chen, X.; Liu, H. Effects of upstream cascade dams and longitudinal environmental gradients on variations in fish assemblages of the Three Gorges Reservoir. Ecol. Freshw. Fish 2021, 30, 503–518. [Google Scholar] [CrossRef]
- Mol, J.H.; de Mérona, B.; Ouboter, P.E.; Sahdew, S. The fish fauna of Brokopondo Reservoir, Suriname, during 40 years of impoundment. Neotrop. Ichthyol. 2007, 5, 351–368. [Google Scholar] [CrossRef]
- Orsi, M.L.; Britton, J.R. Long-term changes in the fish assemblage of a neotropical hydroelectric reservoir. J. Fish Biol. 2014, 84, 1964–1970. [Google Scholar] [CrossRef]
- Schork, G.; Zaniboni-Filho, E. Structure dynamics of a fish community over ten years of formation in the reservoir of the hydroelectric power plant in upper Uruguay River. Brazilian J. Biol. 2017, 77, 710–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loures, R.C.; Pompeu, P.S. Long-term study of reservoir cascade in south-eastern Brazil reveals spatio-temporal gradient in fish assemblages. Mar. Freshw. Res. 2018, 69, 1983. [Google Scholar] [CrossRef] [Green Version]
- Perônico, P.B.; Agostinho, C.S.; Fernandes, R.; Pelicice, F.M. Community reassembly after river regulation: Rapid loss of fish diversity and the emergence of a new state. Hydrobiologia 2020, 847, 519–533. [Google Scholar] [CrossRef]
- Magurran, A.E.; Baillie, S.R.; Buckland, S.T.; Dick, J.M.; Elston, D.A.; Scott, E.M.; Smith, R.I.; Somerfield, P.J.; Watt, A.D. Long-term datasets in biodiversity research and monitoring: Assessing change in ecological communities through time. Trends Ecol. Evol. 2010, 25, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.M. The need for long-term investigations in ecology and the contribution of the Freshwater Biological Association. Freshw. Biol. 1990, 23, 1–5. [Google Scholar] [CrossRef]
- Gido, K.B.; Matthews, W.J.; Wolfinbarger, W.C. Long-term changes in a reservoir fish assemblage: Stability in an unpredictable environment. Ecol. Appl. 2000, 10, 1517–1529. [Google Scholar] [CrossRef]
- Philippi, T.E.; Dixon, P.M.; Taylor, B.E. Detecting trends in species composition. Ecol. Appl. 1998, 8, 300–308. [Google Scholar] [CrossRef]
- dosSantos, N.C.L.; García-Berthou, E.; Dias, J.D.; Lopes, T.M.; de Affonso, I.P.; Severi, W.; Gomes, L.C.; Agostinho, A.A. Cumulative ecological effects of a Neotropical reservoir cascade across multiple assemblages. Hydrobiologia 2018, 819, 77–91. [Google Scholar] [CrossRef]
- Morita, K.; Morita, S.H.; Yamamoto, S. Effects of habitat fragmentation by damming on salmonid fishes: Lessons from white-spotted charr in Japan. Ecol. Res. 2009, 24, 711–722. [Google Scholar] [CrossRef]
- Murchie, K.J.; Schwager, E.; Cooke, S.J.; Danylchuk, A.J.; Danylchuk, S.E.; Goldberg, T.L.; Suski, C.D.; Philipp, D.P. Spatial ecology of juvenile lemon sharks (Negaprion brevirostris) in tidal creeks and coastal waters of Eleuthera, The Bahamas. Environ. Biol. Fishes 2010, 89, 95–104. [Google Scholar] [CrossRef]
- Thornton, K.W.; Kimmel, B.L.; Payne, F.E. Reservoir Limnology: Ecological Perspectives; John Wiley & Sons: Hoboken, NJ, USA, 1990. [Google Scholar]
- Ferrareze, M.; Casatti, L.; Nogueira, M.G. Spatial heterogeneity affecting fish fauna in cascade reservoirs of the Upper Paraná Basin, Brazil. Hydrobiologia 2014, 738, 97–109. [Google Scholar] [CrossRef]
- Loures, R.C.; Pompeu, P.S. Temporal changes in fish diversity in lotic and lentic environments along a reservoir cascade. Freshw. Biol. 2019, 64, 1806–1820. [Google Scholar] [CrossRef]
- Vašek, M.; Kubečka, J.; Peterka, J.; Čech, M.; Draštík, V.; Hladík, M.; Prchalová, M.; Frouzová, J. Longitudinal and Vertical Spatial Gradients in the Distribution of Fish within a Canyon-shaped Reservoir. Int. Rev. Hydrobiol. 2004, 89, 352–362. [Google Scholar] [CrossRef]
- Agostinho, A.; Pelicice, F.; Gomes, L. Dams and the fish fauna of the Neotropical region: Impacts and management related to diversity and fisheries. Braz. J. Biol. 2008, 68, 1119–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agostinho, A.A.; Pelicice, F.M.; Petry, A.C.; Gomes, L.C.; Júlio, H.F. Fish diversity in the upper Paraná River basin: Habitats, fisheries, management and conservation. Aquat. Ecosyst. Health Manag. 2007, 10, 174–186. [Google Scholar] [CrossRef]
- Noble, R.L. Management of lakes, reservoirs, and ponds. In Fisheries Management; Lackey, R.T., Nielsen, L.A., Eds.; John Wiley & Sons: New York, NY, USA, 1980; pp. 265–295. [Google Scholar]
- Franssen, N.R.; Tobler, M. Upstream effects of a reservoir on fish assemblages 45 years following impoundment. J. Fish Biol. 2013, 82, 1659–1670. [Google Scholar] [CrossRef]
- Gao, X.; Zeng, Y.; Wang, J.; Liu, H. Immediate impacts of the second impoundment on fish communities in the Three Gorges Reservoir. Environ. Biol. Fishes 2010, 87, 163–173. [Google Scholar] [CrossRef]
- Lin, P.; Gao, X.; Liu, F.; Li, M.; Liu, H. Long-term monitoring revealed fish assemblage zonation in the Three Gorges Reservoir. J. Oceanol. Limnol. 2019, 37, 1258–1267. [Google Scholar] [CrossRef]
- Wu, H.L.; Shao, K.T.; Lai, C.F. Latin-Chenise Dictionary of Fishes Names; Wu, H.L., Shao, K.T., Lai, C.F., Eds.; The Sueichan Press: Taiwan, China, 1999; ISBN 9578596596. [Google Scholar]
- Adams, M.J. 12—The principles of multivariate data analysis. In Analytical Methods of Food Authentication; Ashurst, P.R., Dennis, M.J., Eds.; Blackie Academic & Professional: London, UK, 1998; p. 308. [Google Scholar]
- Otto, M. Multivariate methods. In Analytical Chemistry; Kellner, R., Mermet, J.M., Otto, M., Widmer, H.M., Eds.; Wiley: Weinheim, Germany, 1998; p. 916. [Google Scholar]
- Vega, M.; Pardo, R.; Barrado, E.; Debán, L. Assessment of seasonal and polluting effects on the quality of river water by exploratory data analysis. Water Res. 1998, 32, 3581–3592. [Google Scholar] [CrossRef]
- Helena, B. Temporal evolution of groundwater composition in an alluvial aquifer (Pisuerga River, Spain) by principal component analysis. Water Res. 2000, 34, 807–816. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in marine communities. An Approach to Statistical Analysis Interpretation, 2nd ed.; Clarke, K.R., Warwick, R.M., Eds.; Primer-E Ltd.: Plymouth, UK, 2001; Volume 2, pp. 1–168. [Google Scholar]
- Clarke, K.R.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Gomes, L.C.; Santos, N.C.L.; Ortega, J.C.G.; Pelicice, F.M. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish. Res. 2016, 173, 26–36. [Google Scholar] [CrossRef]
- Guy, C.S.; Treanor, H.B.; Kappenman, K.M.; Scholl, E.A.; Ilgen, J.E.; Webb, M.A.H. Broadening the Regulated-River Management Paradigm: A Case Study of the Forgotten Dead Zone Hindering Pallid Sturgeon Recovery. Fisheries 2015, 40, 6–14. [Google Scholar] [CrossRef]
- Pelicice, F.M.; Pompeu, P.S.; Agostinho, A.A. Large reservoirs as ecological barriers to downstream movements of Neotropical migratory fish. Fish Fish. 2015, 16, 697–715. [Google Scholar] [CrossRef]
- Matthews, W.J. Influence of Turbid Inflows on Vertical Distribution of Larval Shad and Freshwater Drum. Trans. Am. Fish. Soc. 1984, 113, 192–198. [Google Scholar] [CrossRef]
- Albrecht, B.A.; Holden, P.B.; Kegerries, R.B.; Golden, M.E. Razorback sucker recruitment in Lake Mead, Nevada–Arizona, why here? Lake Reserv. Manag. 2010, 26, 336–344. [Google Scholar] [CrossRef]
- Buckmeier, D.L.; Smith, N.G.; Fleming, B.P.; Bodine, K.A. Intra-annual variation in river-reservoir interface fish assemblages: Implications for fish conservation and management in regulated rivers. River Res. Appl. 2014, 30, 780–790. [Google Scholar] [CrossRef]
- Jennings, D.P. Bighead Carp (Hypophthalmichthys nobilis): A Biological Synopsis; Defence Technical Information Centre: Fort Belvoir, VA, USA, 1988. [Google Scholar]
- Robison, H.W.; Buchanan, T.M. Fishes of Arkansas; University of Arkansas Press: Fayetteville, AR, USA, 2020. [Google Scholar]
- Cremer, M.C.; Smitherman, R.O. Food habits and growth of silver and bighead carp in cages and ponds. Aquaculture 1980, 20, 57–64. [Google Scholar] [CrossRef]
- Xie, P.; Liu, J. Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms: A synthesis of decades of research and application in a subtropical hypereutrophic lake. Sci. World J. 2001, 1, 337–356. [Google Scholar] [CrossRef] [Green Version]
- Xie, P. Gut contents of silver carp, Hypophthalmichthys molitrix, and the disruption of a centric diatom, Cyclotella, on passage through the esophagus and intestine. Aquaculture 1999, 180, 295–305. [Google Scholar] [CrossRef]
- Liu, Q.-G.; Chen, Y.; Li, J.-L.; Chen, L.-Q. The food web structure and ecosystem properties of a filter-feeding carps dominated deep reservoir ecosystem. Ecol. Modell. 2007, 203, 279–289. [Google Scholar] [CrossRef]
- Nakamura, M. Cyprinid Fishes of Japan. Studies on the Life History of Cyprinid Fishes of Japan; Research Institute for Natural Resources: Tokyo, Japan, 1969; Volume 4, pp. 1–455. [Google Scholar]
- Zhao, L.; Chenoweth, E.L.; Liu, J.; Liu, Q. Effects of dam structures on genetic diversity of freshwater fish Sinibrama macrops in Min River, China. Biochem. Syst. Ecol. 2016, 68, 216–222. [Google Scholar] [CrossRef]
- Zhang, D.; Heng, W.; Chu, L.; Xu, D.; Kang, B.; Yan, Y. Taxonomic and functional diversity in a subtropical stream: A longitudinal pattern analysis. Ecol. Freshw. Fish 2020, 29, 752–763. [Google Scholar] [CrossRef]
- Howes, G.J.; Winfield, I.J.; Nelson, J.S. Cyprinid Fishes: Systematics, Biology and Exploitation; Springer Science: Berlin, Germany, 1991. [Google Scholar]
- Koehn, J.D. Carp (Cyprinus carpio) as a powerful invader in Australian waterways. Freshw. Biol. 2004, 49, 882–894. [Google Scholar] [CrossRef]
- Smith, B.B.; Walker, K.F. Spawning dynamics of common carp in the River Murray, South Australia, shown by macroscopic and histological staging of gonads. J. Fish Biol. 2004, 64, 336–354. [Google Scholar] [CrossRef]
- Fletcher, A.R.; Morison, A.K.; Hume, D.J. Effects of carp, Cyprinus carpio L., on communities of aquatic vegetation and turbidity of waterbodies in the lower Goulburn River basin. Mar. Freshw. Res. 1985, 36, 311–327. [Google Scholar] [CrossRef]
- Roberts, J.; Chick, A.; Oswald, L.; Thompson, P. Effect of carp, Cyprinus carpio L., an exotic benthivorous fish, on aquatic plants and water quality in experimental ponds. Mar. Freshw. Res. 1995, 46, 1171. [Google Scholar] [CrossRef]
- Miller, S.A.; Crowl, T.A. Effects of common carp (Cyprinus carpio) on macrophytes and invertebrate communities in a shallow lake. Freshw. Biol. 2006, 51, 85–94. [Google Scholar] [CrossRef]
- King, A.J.; Robertson, A.I.; Healey, M.R. Experimental manipulations of the biomass of introduced carp (Cyprinus carpio) in billabongs. I. Impacts on water-column properties. Mar. Freshw. Res. 1997, 48, 435–443. [Google Scholar] [CrossRef]
- Driver, P.D. The effects of size and density of carp (Cyprinus carpio L.) on water quality in an experimental pond. Arch. Hydrobiol. 2005, 163, 117–131. [Google Scholar] [CrossRef]
- Fishbase. Search Fishbase. Available online: http://www.fishbase.org (accessed on 1 January 2022).
- Chu, X.L.; Chen, Y.R. The Fishes of Yunnan, China. Part 1. Cyprinidae; Science Press: Beijing, China, 1989. [Google Scholar]
- Schlosser, I.J. Fish Community Structure and Function along Two Habitat Gradients in a Headwater Stream. Ecol. Monogr. 1982, 52, 395–414. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Pease, A.A.; González-Díaz, A.A.; Rodiles-Hernández, R.; Winemiller, K.O. Functional diversity and trait-environment relationships of stream fish assemblages in a large tropical catchment. Freshw. Biol. 2012, 57, 1060–1075. [Google Scholar] [CrossRef]
- Agasen, E.V.; Clemente, J.P., Jr.; Rosana, M.R.; Kawit, N.S. Biological Investigation of Jaguar guapote Parachromis managuensis (Gunther, 1867) in Taal Lake, Southern Luzon. In Proceedings of the Conference-Workshop on Alien Invasive Species in the Philippines and their Impact on Biodiversity, Marikina City, Philippines, 26–28 July 2006; pp. 26–28. [Google Scholar]
- Ortega, J.C.G.; Júlio, H.F.; Gomes, L.C.; Agostinho, A.A. Fish farming as the main driver of fish introductions in Neotropical reservoirs. Hydrobiologia 2015, 746, 147–158. [Google Scholar] [CrossRef]
- Garcia, D.A.Z.; Britton, J.R.; Vidotto-Magnoni, A.P.; Orsi, M.L. Introductions of non-native fishes into a heavily modified river: Rates, patterns and management issues in the Paranapanema River (Upper Paraná ecoregion, Brazil). Biol. Invasions 2018, 20, 1229–1241. [Google Scholar] [CrossRef] [Green Version]
- Poff, N.L.; Olden, J.D.; Merritt, D.M.; Pepin, D.M. Homogenization of regional river dynamics by dams and global biodiversity implications. Proc. Natl. Acad. Sci. USA 2007, 104, 5732–5737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Effendie, I.; Moch, H. Biologi Perikanan; Yogyakarta Archipelago Library Foundation: Yogyakarta, Indonesia, 2002. [Google Scholar]
- Buynak, G.L.; Hale, R.S.; Mitchell, B. Differential Growth of Young-of-Year Gizzard Shad in Several Kentucky Reservoirs. N. Am. J. Fish. Manag. 1992, 12, 656–662. [Google Scholar] [CrossRef]
- Olson, M.H.; Mittelbach, G.G.; Osenberg, C.W. Competition between Predator and Prey: Resource-Based Mechanisms and Implications for Stage-Structured Dynamics. Ecology 1995, 76, 1758–1771. [Google Scholar] [CrossRef]
- Prout, M.W.; Mills, E.L.; Forney, J.L. Diet, Growth, and Potential Competitive Interactions between Age-0 White Perch and Yellow Perch in Oneida Lake, New York. Trans. Am. Fish. Soc. 1990, 119, 966–975. [Google Scholar] [CrossRef]
- Welker, M.T.; Pierce, C.L.; Wahl, D.H. Growth and Survival of Larval Fishes: Roles of Competition and Zooplankton Abundance. Trans. Am. Fish. Soc. 1994, 123, 703–717. [Google Scholar] [CrossRef]
- Fletcher, C.M.; Collins, S.F.; Nannini, M.A.; Wahl, D.H. Competition during early ontogeny: Effects of native and invasive planktivores on the growth, survival, and habitat use of bluegill. Freshw. Biol. 2019, 64, 697–707. [Google Scholar] [CrossRef]
- Chick, J.H.; Pegg, M.A. Invasive Carp in the Mississippi River Basin. Science 2001, 292, 2250–2251. [Google Scholar] [CrossRef]
- Taki, Y. An analytical study of the fish fauna of the Mekong basin as a biological production system in nature. Res. Inst. Evol. Biol. 1978, 1, 1–77. [Google Scholar]
- Rathod, S.R.; Khedkar, G.D. Impact of elevation, latitude and longitude on fish diversity in Godavari River. J. Res. Biol. 2011, 4, 269–275. [Google Scholar]




{kind=link}
{kind=link}
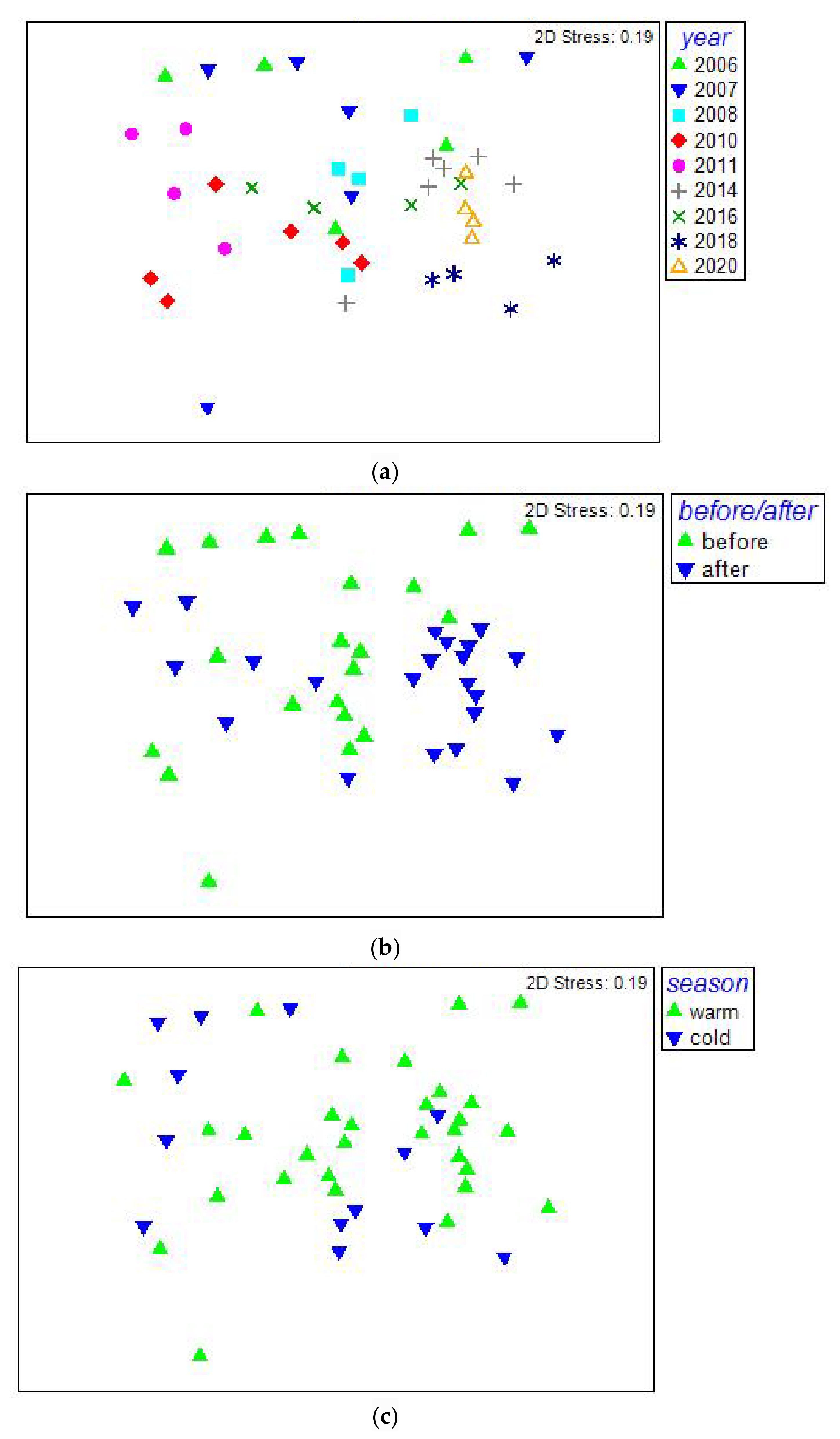{kind=link}
| PC1 | PC2 | PC3 | PC4 | PC5 | |
|---|---|---|---|---|---|
| Eigenvalue | 1.12 | 0.279 | 0.146 | 0.115 | 2.99 E-2 |
| Total variance (%) | 64.5 | 16.1 | 8.4 | 6.6 | 1.7 |
| Cumulative variance proportion (%) | 64.5 | 80.6 | 89.0 | 95.6 | 97.4 |
| Loading of Variables | |||||
|---|---|---|---|---|---|
| Variables | PC1 | PC2 | PC3 | PC4 | PC5 |
| SD | −0.076 | 0.285 | −0.200 | 0.251 | −0.261 |
| WT | 0.102 | −0.062 | 0.045 | −0.007 | 0.837 |
| Turbidity | 0.061 | −0.426 | 0.421 | −0.673 | −0.193 |
| pH | 0.019 | −0.012 | −0.010 | −0.011 | 0.163 |
| DO | 0.007 | 0.052 | −0.010 | −0.016 | −0.277 |
| TDS | 0.025 | −0.022 | −0.014 | 0.027 | 0.102 |
| EC | −0.004 | −0.058 | −0.050 | 0.000 | 0.244 |
| TP | 0.093 | −0.487 | 0.509 | 0.694 | −0.091 |
| Chl-a | 0.154 | −0.668 | −0.720 | 0.027 | −0.072 |
| Algal count | 0.973 | 0.208 | 0.019 | −0.008 | −0.079 |
| Pairwise Test | ||||||||
|---|---|---|---|---|---|---|---|---|
| p-Value | 2006 | 2007 | 2008 | 2010 | 2011 | 2014 | 2016 | 2018 |
| 2007 | 0.537 | |||||||
| 2008 | 0.476 | 0.8 | ||||||
| 2010 | 0.017 | 0.048 | 0.11 | |||||
| 2011 | 0.127 | 0.133 | 0.029 | 0.19 | ||||
| 2014 | 0.019 | 0.004 | 0.029 | 0.002 | 0.005 | |||
| 2016 | 0.167 | 0.305 | 0.029 | 0.024 | 0.029 | 0.019 | ||
| 2018 | 0.024 | 0.005 | 0.029 | 0.005 | 0.029 | 0.005 | 0.029 | |
| 2020 | 0.071 | 0.062 | 0.029 | 0.005 | 0.029 | 0.010 | 0.057 | 0.057 |
| Species | Contribution to Dissimilarity (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| G 2006 vs. G 2010 | G 2007 vs. G 2010 | G 2008 vs. G 2011 | G 2006 vs. G 2014 | G 2011 vs. G 2014 | G 2010 vs. G 2016 | G 2011 vs. G 2018 | G 2011 vs. G 2020 | |
| Carassius cuvieri | 10.34 | 10.16 | ||||||
| Chanodichthys erythropterus | 12.68 | |||||||
| Cyprinus carpio | 10.26 | |||||||
| Distoechodon tumirostris | 10.92 | 11.52 | ||||||
| Hemiculter leucisculus | 11.23 | 12.76 | ||||||
| Hypophthalmichthys nobilis | 13.18 | |||||||
| Mozambique tilapia | 10.22 | 10.83 | 13.68 | 10.25 | ||||
| Parachromis managuensis | 16.47 | 16.62 | ||||||
| Sinibrama macrops | 12.67 | 13.1 | 17.88 | |||||
| Feitsui Reservoir | Cold and Warm Season | Pre and Post 2010 Period | Years |
|---|---|---|---|
| Global R | 0.088 | 0.168 | 0.369 |
| p-value | 0.100 | 0.001 | 0.001 |
| Groups Before & After | ||
|---|---|---|
| Average Dissimilarity = 53.94 | ||
| Species | Contrib (%) | Cum. (%) |
| Sinibrama macrops | 10.8 | 10.8 |
| Oreochromis sp. | 8.62 | 19.42 |
| Culter erythropterus | 8.07 | 27.49 |
| Carassius auratus auratus | 7.56 | 35.05 |
| Hemiculter leucisculus | 7.29 | 42.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi-Ron, C.; Wen-Shang, H.; Chen-Kang, H.; Chu-Yang, C. Spatial and Temporal Variations in Fish Assemblage in Feitsui Reservoir, in Northern Taiwan, from 2006–2020. Water 2022, 14, 498. https://doi.org/10.3390/w14030498
Yi-Ron C, Wen-Shang H, Chen-Kang H, Chu-Yang C. Spatial and Temporal Variations in Fish Assemblage in Feitsui Reservoir, in Northern Taiwan, from 2006–2020. Water. 2022; 14(3):498. https://doi.org/10.3390/w14030498
Chicago/Turabian StyleYi-Ron, Chen, Hou Wen-Shang, Huang Chen-Kang, and Chou Chu-Yang. 2022. "Spatial and Temporal Variations in Fish Assemblage in Feitsui Reservoir, in Northern Taiwan, from 2006–2020" Water 14, no. 3: 498. https://doi.org/10.3390/w14030498