
Effects of Temperature and Food Concentration on the Population Recruitment of Acartia bifilosa (Copepoda, Calanoida): Implications for the Over-Summering Life History Strategy in Jiaozhou Bay
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
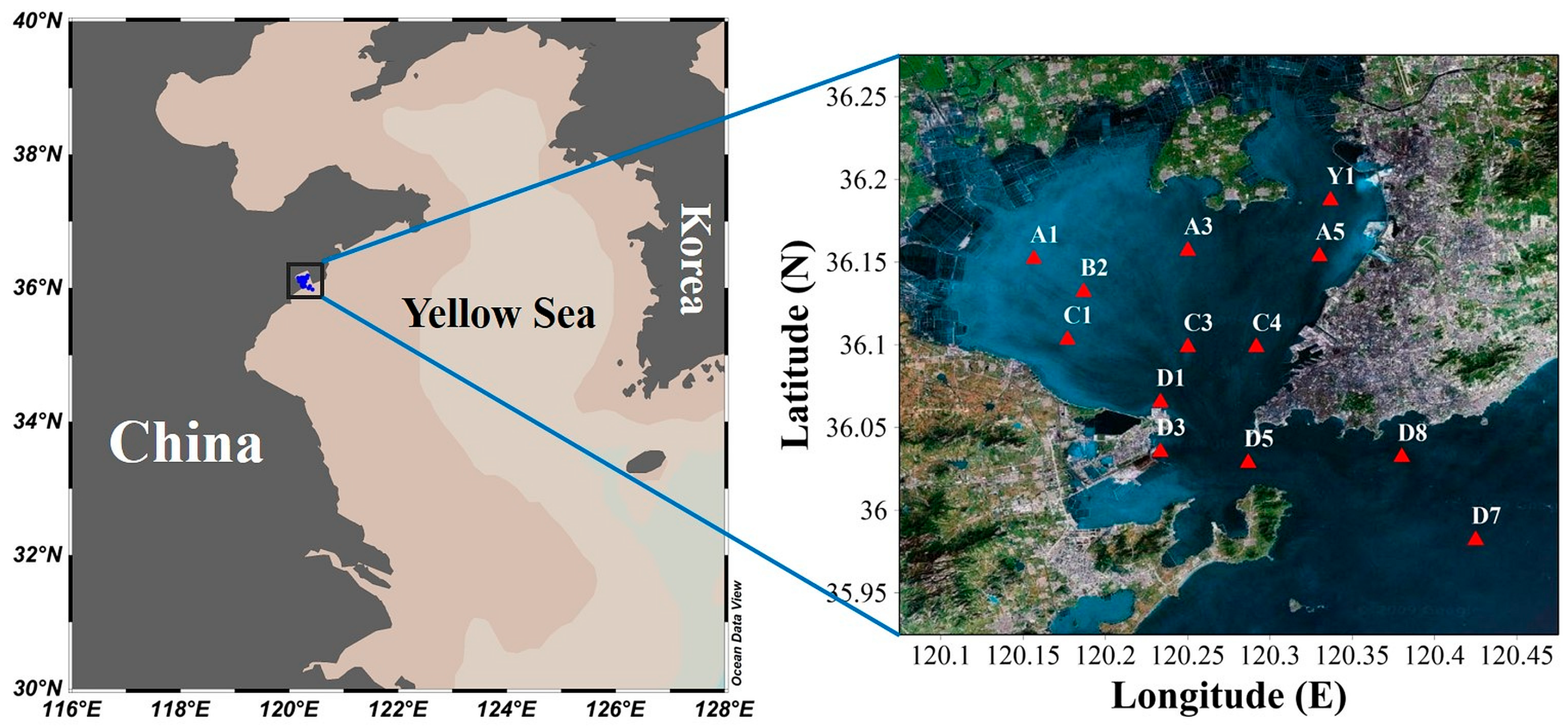
2.1. Field Investigation
2.2. Laboratory Experiments
2.3. Data Analysis
3. Results
3.1. Environmental Conditions and Distribution of Acartia bifilosa Abundance
3.2. Female Survival
3.3. Egg Production
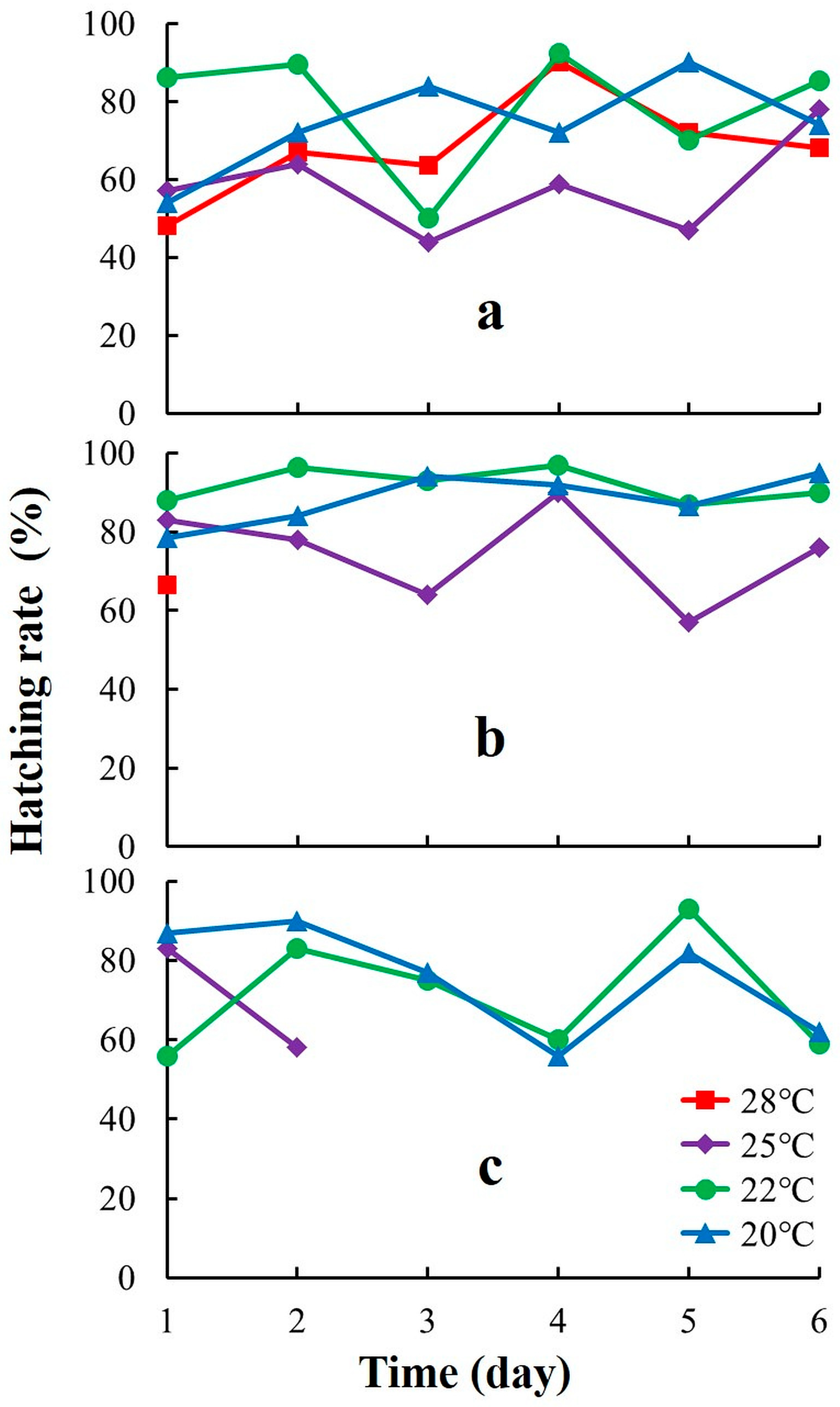
3.4. Hatching Success
3.5. Post-Embryonic Development
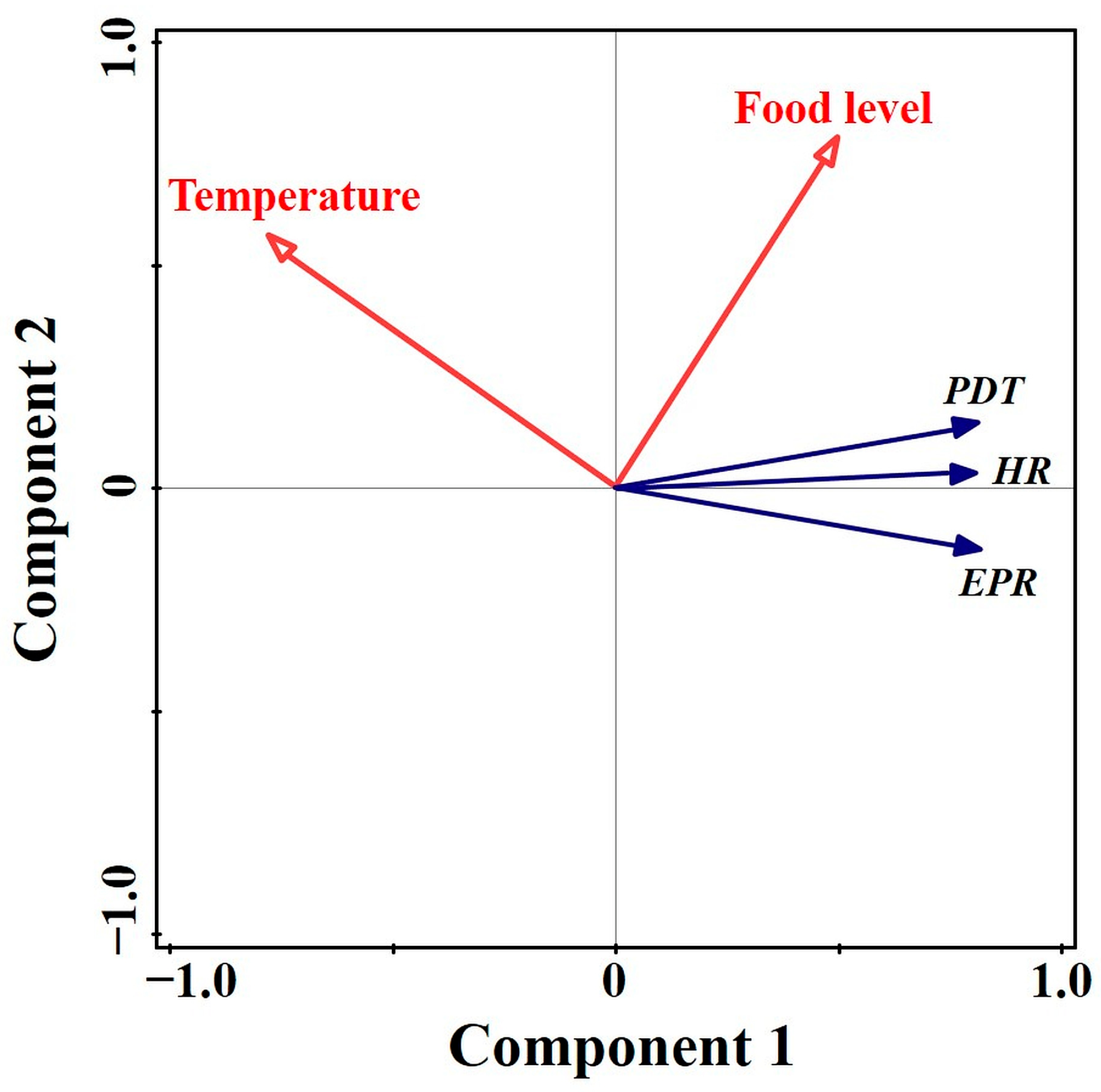
3.6. Relationship between Acartia bifilosa Population Recruitment and Environmental Factors
4. Discussion
4.1. Combination Effect of Temperature and Food Concentration
4.2. Implications for the Over-Summering Life History Strategy in the Jiaozhou Bay
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleppel, G.S. On the diets of calanoid copepods. Mar. Ecol. Prog. Ser. 1993, 99, 183–195. [Google Scholar] [CrossRef]
- Dubischar, C.D.; Lopes, R.M.; Bathmann, U.V. High summer abundances of small pelagic copepods at the Antarctic Polar Front-implications for ecosystem dynamics. Deep-Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 3871–3887. [Google Scholar] [CrossRef] [Green Version]
- Fiksen, O.; Giske, J. Vertical distribution and population dynamics of copepods by dynamic optimization. ICES J. Mar. Sci. 1995, 52, 483–503. [Google Scholar] [CrossRef]
- Grice, G.D.; Marcus, N.H. Dormant eggs of marine copepods. Oceanol. Mar. Biol. Annu. Rev. 1981, 19, 125–140. [Google Scholar]
- Dahms, H.U. Dormancy in the Copepoda—An Overview. Hydrobiologia 1995, 306, 199–211. [Google Scholar] [CrossRef]
- Roff, D.A. The Evolution of Life Histories: Theory and Analysis; Chapman and Hall: New York, NY, USA, 1992; p. 535. [Google Scholar]
- Halsband-Lenk, C.; Hirche, H.J.; Carlotti, F. Temperature impact on reproduction and development of congener copepod populations. J. Exp. Mar. Biol. Ecol. 2002, 271, 121–153. [Google Scholar] [CrossRef] [Green Version]
- Collins, N.R.; Williams, R. Zooplankton Communities in the Bristol Channel and Severn Estuary. Mar. Ecol. Prog. Ser. 1982, 9, 1–11. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, H.Y.; Wang, K.; Zuo, T. Function performed by small copepods in marine ecosystem. Oceanol. Limnol. Sin. 2002, 33, 453–460. [Google Scholar]
- Sun, X.H.; Sun, S.; Li, C.L.; Zhang, G.T. Seasonal and spatial variability in egg production, abundance and production of small copepods in and near Jiaozhou Bay, China. J. Plankton Res. 2011, 33, 741–750. [Google Scholar] [CrossRef]
- Ge, R.P.; Chen, H.J.; Liu, G.X.; Zhu, Y.Z.; Jiang, Q. Diel vertical migration of mesozooplankton in the northern Yellow Sea. J. Oceanol. Limnol. 2021, 39, 1373–1386. [Google Scholar] [CrossRef]
- Hansson, S.; Larsson, U.; Johansson, S. Selective Predation by Herring and Mysids, and Zooplankton Community Structure in a Baltic Sea Coastal Area. J. Plankton Res. 1990, 12, 1099–1116. [Google Scholar] [CrossRef]
- Viitasalo, M. Calanoid Resting Eggs in the Baltic Sea—Implications for the Population-Dynamics of Acartia Bifilosa (Copepoda). Mar. Biol. 1992, 114, 397–405. [Google Scholar] [CrossRef]
- Sun, S.; Li, C.L.; Zhang, G.T.; Sun, X.X.; Yang, B. Long-term changes in the zooplankton community in the Jiaozhou bay. Oceanol. Limnol. Sin. 2011, 42, 625–631. [Google Scholar]
- Uriate, I.; Cotano, U.; Villate, F. Egg production of Acartia bifilosa in the small temperate estuary of Mundaka, Spain, in relation to environmental variables and population development. Mar. Ecol. Prog. Ser. 1998, 166, 197–205. [Google Scholar] [CrossRef]
- Burdloff, D.; Gasparini, S.; Villate, F.; Uriarte, I.; Cotano, U.; Sautour, B.; Etcheber, H. Egg production of copepod Acartia bifilosa in two contrasting European estuaries in relation to seston composition. J. Exp. Mar. Biol. Ecol. 2002, 274, 1–17. [Google Scholar] [CrossRef]
- Gorokhova, E. Relationships between nucleic acid levels and egg production rates in Acartia bifilosa: Implications for growth assessment of copepods in situ. Mar. Ecol. Prog. Ser. 2003, 262, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Castro-Longoria, E.; Williams, J.A. The production of subitaneous and diapause eggs: A reproductive strategy for Acartia bifilosa (Copepoda: Calanoida) in Southampton Water, UK. J. Plankton. Res. 1999, 21, 65–84. [Google Scholar] [CrossRef]
- Koski, M.; Kuosa, H. The effect of temperature, food concentration and female size on the egg production of the planktonic copepod Acartia bifilosa. J. Plankton Res. 1999, 21, 1779–1789. [Google Scholar] [CrossRef]
- Chinnery, F.E.; Williams, J.A. Photoperiod and temperature regulation of diapause egg production in Acartia bifilosa from Southampton Water. Mar. Ecol. Prog. Ser. 2003, 263, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Katajisto, T. Development of Acartia bifilosa (Copepoda: Calanoida) eggs in the northern Baltic Sea with special reference to dormancy. J. Plankton Res. 2003, 25, 357–364. [Google Scholar] [CrossRef]
- Choi, S.Y.; Jang, M.C.; Youn, S.H.; Seo, M.H.; Soh, H.Y. Egg production and hatching patterns of Acartia erythraea (Copepoda, Calanoida), with a note on its two egg types, in a eutrophic bay in Korea. J. Plankton Res. 2021, 43, 428–441. [Google Scholar] [CrossRef]
- Zhong, X.F.; Xiao, Y.C. Resting eggs of Acartia bifilosa Giesbrecht and A. pacifica Steuer in the Jiaozhou Bay. Mar. Sci. 1992, 5, 55–59. [Google Scholar]
- Zhong, X.F.; Xiao, Y.C. Seasonal cycles of abundance of three copepod species population in the Jiaozhou Bay. Mar. Sci. 1992, 1, 44–48. [Google Scholar]
- Liu, R.Y. Characteristics of Physical Environmental of the Jiaozhou Bay; Science Press: Beijing, China, 1992; pp. 2–3. [Google Scholar]
- Shen, Z.L.; Liu, Q.; Wu, Y.L.; Yao, Y. Nutrient structure of seawater and ecological responses in the Jiaozhou Bay, China. Estuar. Coast. Shelf Sci. 2006, 69, 299–307. [Google Scholar]
- Sun, S.; Sun, X.X.; Zhang, G.T.; Tang, H.B.; Liu, Q.; Li, G.M. Long-term changes in major meteorological and hydrological factors in the Jiaozhou bay. Oceanol. Limnol. Sin. 2011, 42, 632–638. [Google Scholar]
- Li, C.L.; Zhang, F.; Shen, X.; Yang, B.; Shen, Z.L.; Sun, S. Concentration, distribution and annual fluctuation of chlorophyll-a in the Jiaozhou Bay. Oceanol. Limnol. Sin. 2005, 36, 499–506. [Google Scholar]
- Sun, X.X.; Sun, S.; Zhang, Y.S.; Zhang, F. Long-term changes of chlorophyll-a concentration and primary productivity in the Jiaozhou bay. Oceanol. Limnol. Sin. 2011, 42, 654–661. [Google Scholar]
- Paffenhöfer, G.A.; Stearns, D.E. Why is Acartia tonsa (Copepoda: Calanoida) restricted to nearshore environments? Mar. Ecol. Prog. Ser. 1988, 42, 33–38. [Google Scholar] [CrossRef]
- Durbin, E.G.; Durbin, A.G.; Smayda, T.J.; Verity, P.G. Food Limitation of Production by Adult Acartia tonsa in Narragansett Bay, Rhode-Island. Limnol. Oceanogr. 1983, 28, 1199–1213. [Google Scholar] [CrossRef]
- Irigoien, X.; Castel, J. Feeding rates and production of the copepod Acartia bifilosa in a highly turbid estuary; the Gironde (SW France). Hydrobiologia 1996, 311, 115–125. [Google Scholar] [CrossRef]
- Hairston, N.G.; Caceres, C.E. Distribution of crustacean diapause: Micro- and macroevolutionary pattern and process. Hydrobiologia 1996, 320, 27–44. [Google Scholar] [CrossRef]
- Li, J.; Sun, S.; Li, C.L.; Zhang, Z.; Pu, X.M. Effects of different diets on the reproduction and naupliar development of the copepod Acartia bifilosa. J. Exp. Mar. Biol. Ecol. 2008, 355, 95–102. [Google Scholar] [CrossRef]
- Tao, Z.C.; Wang, Y.Q.; Wang, J.J.; Liu, M.T.; Zhang, W.C. Photobehaviors of the calanoid copepod Calanus sinicus from the Yellow Sea to visible and UV-B radiation as a function of wavelength and intensity. J. Oceanol. Limnol. 2019, 37, 1289–1300. [Google Scholar] [CrossRef]
- Yoon, W.D.; Shim, M.B.; Choi, J.K. Description of the developmental stages in Acartia bifilosa Giesbrecht (Copepoda: Calanoida). J. Plankton Res. 1998, 20, 923–942. [Google Scholar] [CrossRef] [Green Version]
- Chinnery, F.E.; Williams, J.A. The influence of temperature and salinity on Acartia (Copepoda: Calanoida) nauplii survival. Mar. Biol. 2004, 145, 733–738. [Google Scholar] [CrossRef]
- Pu, X.M.; Sun, S.; Yang, B.; Ji, P.; Zhang, Y.S.; Zhang, F. The combined effects of temperature and food supply on Calanus sinicus in the southern Yellow Sea in summer. J. Plankton Res. 2004, 26, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Vehmaa, A.; Hogfors, H.; Gorokhova, E.; Brutemark, A.; Holmborn, T.; Engstrom-Ost, J. Projected marine climate change: Effects on copepod oxidative status and reproduction. Ecol. Evol. 2013, 3, 4548–4557. [Google Scholar] [CrossRef] [Green Version]
- David, D.; Souissi, S.; Seuront, L. Effects of Chlorophyll concentration and temperature variation on the reproduction and survival of Temora longicornis (Copepoda, Calanoida) in the Eastern English Channel. J. Exp. Mar. Biol. Ecol. 2005, 318, 145–162. [Google Scholar]
- Ianora, A.; Miralto, A.; Halsband-Lenk, C. Reproduction, hatching success, and early naupliar survival in Centropages typicus. Prog. Oceanogr. 2007, 72, 195–213. [Google Scholar] [CrossRef]
- Souissi, S.; Carlotti, F.; Nival, P. Food and temperature dependent function of moulting rate in copepods: An example of parameterization for population dynamics models. J. Plankton Res. 1997, 19, 1331–1346. [Google Scholar] [CrossRef]
- Uye, S. Resting egg production as a life history strategy of marine planktonic copepods. Bull. Mar. Sci. 1985, 37, 440–449. [Google Scholar]
- Marcus, N.H. Ecological and evolutionary significance of resting eggs in marine copepods: Past, present, and future studies. Hydrobiologia 1996, 320, 141–152. [Google Scholar] [CrossRef]
- Katajisto, T. Effects of anoxia and hypoxia on the dormancy and survival of subitaneous eggs of Acartia bifilosa (Copepda: Calanoida). Mar. Biol. 2004, 145, 751–757. [Google Scholar]
- Sun, X.H.; Sun, S.; Li, C.L.; Wang, M.X. Seasonal change in body length of important small copepods and relationship with environmental factors in Jiaozhou Bay, China. Chin. J. Oceanol. Limn. 2012, 30, 404–409. [Google Scholar] [CrossRef]
- Drillet, G.; Hansen, B.W.; Kiorboe, T. Resting egg production induced by food limitation in the calanoid copepod Acartia tonsa. Limnol. Oceanogr. 2011, 56, 2064–2070. [Google Scholar]
- Engstrom-Ost, J.; Holmborn, T.; Brutemark, A.; Hogfors, H.; Vehmaa, A.; Gorokhova, E. The effects of short-term pH decrease on the reproductive output of the copepod Acartia bifilosa—A laboratory study. Mar. Freshw. Behav. Phy. 2014, 47, 173–183. [Google Scholar] [CrossRef]
- Broman, E.; Brusinm, M.; Dopsonm, M.; Hylanderm, S. Oxygenation of anoxic sediments triggers hatching of zooplankton eggs. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152025. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Level (μg/L) | Temperature (°C) | avEPR | avHR (%) | PDT |
|---|---|---|---|---|
| 15 | 28 | 0.73 | 68 | 10 |
| 25 | 2.39 | 78 | 9 | |
| 22 | 10.35 | 85 | 11 | |
| 20 | 11.94 | 74 | 12 | |
| 10 | 28 | 0.02 | 67 | 11 |
| 25 | 0.74 | 76 | 10 | |
| 22 | 10.21 | 91 | 11.5 | |
| 20 | 10.41 | 88 | 12 | |
| 5 | 28 | 0 | * | ** |
| 25 | 0.11 | 71 | ** | |
| 22 | 1.89 | 77 | 12 | |
| 20 | 3.85 | 75 | 12 | |
| 0 | 28 | 0 | * | * |
| 25 | 0 | * | * | |
| 22 | 0 | * | * | |
| 20 | 0 | * | * |
| avEPR | avHR | PDT | |
|---|---|---|---|
| Temperature (°C) | −0.775 ** | −0.629 * | −0.722 * |
| Food level (μg/L) | 0.432 | 0.073 | −0.542 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Tao, Z.; Gao, X.; Wang, L.; Sun, S. Effects of Temperature and Food Concentration on the Population Recruitment of Acartia bifilosa (Copepoda, Calanoida): Implications for the Over-Summering Life History Strategy in Jiaozhou Bay. Water 2022, 14, 3541. https://doi.org/10.3390/w14213541
Zhang Z, Tao Z, Gao X, Wang L, Sun S. Effects of Temperature and Food Concentration on the Population Recruitment of Acartia bifilosa (Copepoda, Calanoida): Implications for the Over-Summering Life History Strategy in Jiaozhou Bay. Water. 2022; 14(21):3541. https://doi.org/10.3390/w14213541
Chicago/Turabian StyleZhang, Zhan, Zhencheng Tao, Xiaotong Gao, Lei Wang, and Song Sun. 2022. "Effects of Temperature and Food Concentration on the Population Recruitment of Acartia bifilosa (Copepoda, Calanoida): Implications for the Over-Summering Life History Strategy in Jiaozhou Bay" Water 14, no. 21: 3541. https://doi.org/10.3390/w14213541