
Ice Melt-Induced Variations of Structural and Functional Traits of the Aquatic Microbial Community along an Arctic River (Pasvik River, Norway)

 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Collection
2.3. Environmental Chemical Variables
2.3.1. Particulate Material
2.3.2. Dissolved Nutrients
2.3.3. Photosynthetic Pigment Contents
2.4. Microbial Abundanc
2.4.1. Total Microbial Cell Abundance by Flow Cytometry (MA-FC)
2.4.2. Estimation of Total Prokaryotic Biomass, Cell Volume and Morphotypes by Image Analysis
2.5. Microbial Activity Measurements
2.5.1. Extracellular Enzymatic Activity
2.5.2. Respiratory Activity
2.6. Phylogenetic Composition of the Bacterial Community
2.7. Statistical Analyses
3. Results
3.1. Physical-Chemical Parameters at Sampling Time
3.2. Environmental Chemical Variables
3.2.1. Particulate Material
3.2.2. Dissolved Nutrients
3.2.3. Photosynthetic Pigments
3.3. Estimation of Microbial Abundance and Morphometric Traits
3.3.1. Total Microbial Cell Abundance by Flow Cytometry (MA-FC)
3.3.2. Cell Volume, Prokaryotic Biomass, and Morphotypes by Image Analysis
3.4. Microbial Activity Measurements
3.4.1. Extracellular Enzymatic Activity
3.4.2. Respiratory Activity
3.5. Phylogenetic Composition of the Bacterial Community
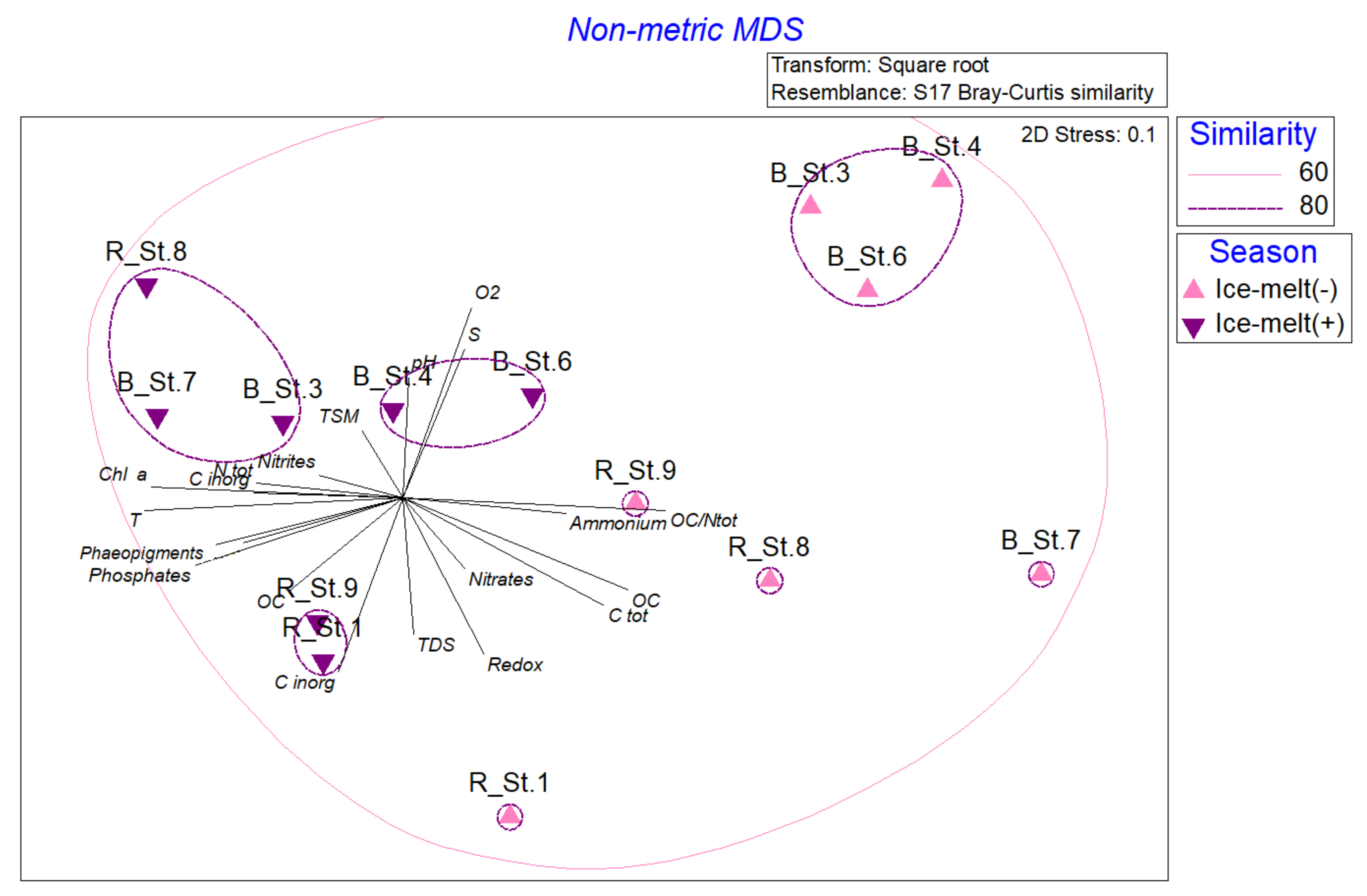
3.6. Statistical Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bring, A.; Fedorova, I.; Dibike, Y.; Hinzman, L.; Mård, J.; Mernild, S.H.; Prowse, T.; Semenova, O.; Stuefer, S.L.; Wooet, M.-K. Arctic terrestrial hydrology: A synthesis of processes, regional effects and research challenges. J. Geophys. Res. Biogeosci. 2016, 121, 621–649. [Google Scholar] [CrossRef]
- Hauptmann, A.L.; Markussen, T.N.; Stibal, M.; Olsen, N.S.; Elberling, B.; Baelum, J.; Sicheritz-Pontén, T.; Jacobsen, C.S. Upstream freshwater and terrestrial sources are differentially reflected in the bacterial community structure along a small Arctic river and its estuary. Front. Microbiol. 2016, 7, 1474. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.E.; Saros, J.E.; Vincent, W.F.; Smol, J.P. Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol. Oceanogr. 2009, 54, 2273–2282. [Google Scholar] [CrossRef]
- Colby, G.A.; Ruuskanen, M.O.; St. Pierre, K.A.; St. Louis, V.L.; Poulain, A.J.; Aris-Brosou, S. Climate change lowers diversity and functional potential of microbes in Canada’s high Arctic. bioRxiv 2019, 705178. [Google Scholar] [CrossRef] [Green Version]
- Dinasquet, J.; Ortega-Retuerta, E.; Lovejoy, C.; Obernosterer, I. Editorial: Microbiology of the Rapidly Changing Polar Environments. Front. Mar. Sci. 2018, 5, 154. [Google Scholar] [CrossRef]
- Sipler, R.E.; Kellogg, C.T.E.; Connelly, T.L.; Roberts, Q.N.; Yager, P.L.; Bronk, D.A. Microbial community response to terrestrially derived dissolved organic matter in the coastal Arctic. Front. Microbiol. 2017, 8, 1018. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, M.L.; Nielsen, S.E.B.; Müller, O.; Møller, E.F.; Stedmon, C.A.; Juul-Pedersen, T.; Markager, S.; Sejr, M.K.; Delgado Huertas, A.; Larsen, A.; et al. Carbon bioavailability in a high Arctic fjord influenced by glacial meltwater, NE Greenland. Front. Mar. Sci. 2017, 4, 176. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-González, C.; Niño-García, J.P.; Del Giorgio, P.A. Terrestrial origin of bacterial communities in complex boreal freshwater networks. Ecol. Lett. 2015, 18, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Niño-García, J.P.; Ruiz-González, C.; del Giorgio, P.A. Interactions between hydrology and water chemistry shape bacterioplankton biogeography across boreal freshwater networks. ISME J. 2016, 10, 1755–1766. [Google Scholar] [CrossRef] [Green Version]
- Caputo, S.; Papale, M.; Rizzo, C.; Giannarelli, S.; Conte, A.; Moscheo, F.; Graziano, M.; Aspholm, P.E.; Onor, M.; De Domenico, E.; et al. Heavy metal resistance in bacteria from contaminated Arctic sediment is driven by heavy metal local inputs. Arch. Environ. Contam. Toxicol. 2019, 77, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, M.A.; St. Louis, V.L.; Engel, K.; St. Pierre, K.A.; Schiff, S.L.; Stibal, M.; Neufeld, J.D. Freshwater microbial community diversity in a rapidly changing High Arctic watershed. FEMS Microbiol. Ecol. 2019, 95, fiz161. [Google Scholar] [CrossRef]
- Rappazzo, A.C.; Papale, M.; Rizzo, C.; Conte, A.; Giannarelli, S.; Onor, M.; Abete, C.; Cefali, P.; De Domenico, E.; Michaud, L.; et al. Heavy metal tolerance and polychlorinated biphenyl oxidation in bacterial communities inhabiting the Pasvik River and the Varanger Fjord area (Arctic Norway). Mar. Pollut. Bull. 2019, 141, 535–549. [Google Scholar] [CrossRef]
- Papale, M.; Rappazzo, A.C.; Mikkonen, A.; Rizzo, C.; Moscheo, F.; Conte, A.; Michaud, L.; Lo Giudice, A. Bacterial diversity in a dynamic and extreme sub-Arctic watercourse (Pasvik River, Norwegian Arctic). Water 2020, 12, 3098. [Google Scholar] [CrossRef]
- Laganà, P.; Votano, L.; Caruso, G.; Azzaro, M.; Lo Giudice, A.; Delia, S. Bacterial isolates from the Arctic region (Pasvik River, Norway): Assessment of biofilm production and antibiotic susceptibility profiles. Environ. Sci. Pollut. Res. 2018, 25, 1089–1102. [Google Scholar] [CrossRef] [PubMed]
- Tesi, T.; Miserocchi, S.; Acri, F.; Langone, L.; Boldrin, A.; Hatten, J.A.; Albertazzi, S. Flood-driven transport of sediment, particulate organic matter, and nutrients from the Po River watershed to the Mediterranean Sea. J. Hydrol. 2013, 498, 144–152. [Google Scholar] [CrossRef]
- Tesi, T.; Miserocchi, S.; Goñi, M.; Langone, L.; Boldrin, A.; Turchetto, M. Organic matter origin and distribution in suspended particulate materials and superficial sediments from the western Adriatic Sea (Italy). Estuar. Coast. Shelf Sci. 2007, 73, 431–446. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A practical hand book of seawater analysis. Fish. Res. Board Can. Bull. 1972, 157, 310. [Google Scholar]
- Aminot, A.; Chaussepied, M. Manuel des analyses chimiques en milieu marin. CNEXO Ed. Jouvre Paris 1983, 395. [Google Scholar]
- Yentsch, C.S.; Menzel, D.W. A method for the determination of phytoplankton chlorophyll and phaeophytin by fluorescence. Deep Sea Res. 1963, 7, 221–231. [Google Scholar] [CrossRef]
- Lorenzen, C.I. Determination of chlorophyll and phaeopigments spectro-photometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Zoppini, A.; Ademollo, N.; Bensi, M.; Berto, D.; Bongiorni, L.; Campanelli, A.; Casentini, B.; Patrolecco, L.; Amalfitano, S. Impact of a river flood on marine water quality and planktonic microbial communities. Estuar. Coast. Shelf Sci. 2019, 224, 62–72. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- La Ferla, R.; Maimone, G.; Azzaro, M.; Conversano, F.; Brunet, C.; Cabral, A.S.; Paranhos, R. Vertical distribution of the prokaryotic cell size in the Mediterranean Sea. Helgol. Mar. Res. 2012, 66, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, H.G. Use of fluorogenic model substrates for extracellular enzyme activity (eea) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publisher: Boca Raton, FL, USA, 1993; pp. 423–432. [Google Scholar]
- La Ferla, R.; Azzaro, M.; Maimone, G. Microbial respiration and trophic regimes in the Northern Adriatic Sea (Mediterranean Sea). Estuar. Coast. Shelf Sci. 2006, 69, 196–204. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M. Microbial respiration in the Levantine Sea: Evolution of the oxidative processes in relation to the main Mediterranean water masses. Deep Sea Res. Part I 2001, 48, 2147–2159. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M.; Chiodo, G. Microplankton respiratory activity and CO2 production rates in the Otranto Strait (Mediterranean Sea). Aquat. Ecol. 1999, 33, 157–165. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D. Microbial communities and processes in Arctic permafrost environments. In Microbiology of Extreme Soils. Soil Biology; Dion, P., Nautiyal, C.S., Eds.; Springer: Berlin, Heidelberg, 2008; Volume 13, pp. 133–154. [Google Scholar]
- IPCC. Summary for Policymakers, book section SPM, 1–30; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; Available online: http://www.climatechange2013.org (accessed on 19 August 2021).
- Melillo, J.M.; Steudler, P.A.; Aber, J.D.; Newkirk, K.; Lux, H.; Bowles, F.P.; Catricala, C.; Magill, A.; Ahrens, T.; Morrisseau, S. Soil warming and carbon-cycle feedbacks to the climate system. Science 2002, 298, 2173–2176. [Google Scholar] [CrossRef]
- Zimov, S.A.; Schuur, E.A.G.; Chapin III, F.S. Permafrost and the global carbon budget. Science 2006, 312, 1612–1613. [Google Scholar] [CrossRef]
- Still, C.J.; Berry, J.A.; Collatz, G.J.; DeFries, R.S. Global distribution of C3 and C4 vegetation: Carbon cycle implications. Glob. Biogeochem. Cycles 2003, 17, 1006. [Google Scholar] [CrossRef]
- Caruso, G.; Madonia, A.; Bonamano, S.; Miserocchi, S.; Giglio, F.; Maimone, G.; Azzaro, F.; Decembrini, F.; La Ferla, R.; Piermattei, V.; et al. Microbial abundance and enzyme activity patterns: Response to changing environmental characteristics along a transect in Kongsfjorden (Svalbard Islands). J. Mar. Sci. Eng. 2020, 8, 824. [Google Scholar] [CrossRef]
- Fortunato, C.S.; Crump, B.C. Bacterioplankton community variation across river to ocean environmental gradients. Microb. Ecol. 2011, 62, 374–382. [Google Scholar] [CrossRef]
- Fortunato, C.S.; Herfort, L.; Zuber, P.; Baptista, A.M.; Crump, B.C. Spatial variability overwhelms seasonal patterns in bacterioplankton communities across a river to ocean gradient. ISME J. 2012, 6, 554–563. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Kolmakova, O.V.; Tolomeev, A.P.; Anishchenko, O.V.; Makhutova, O.N.; Kolmakova, A.A.; Kravchuk, E.S.; Glushchenko, L.A.; Kolmakov, V.I.; Sushchik, N.N. Differences in organic matter and bacterioplankton between sections of the largest Arctic river: Mosaic or continuum? Limnol. Oceanogr. 2015, 60, 1314–1331. [Google Scholar] [CrossRef]
- Ortega-Retuerta, E.; Jeffrey, W.H.; Babin, M.; Belanger, S.; Benner, R.; Marie, D.; Matsuoka, A.; Raimbault, P.; Joux, F. Carbon fluxes in the Canadian Arctic: Patterns and drivers of bacterial abundance, production and respiration on the Beaufort Sea margin. Biogeosciences 2012, 9, 3679–3692. [Google Scholar] [CrossRef] [Green Version]
- Duda, V.I.; Suzina, N.E.; Polivtseva, V.N.; Boronin, A.M. Ultramicrobacteria: Formation of the concept and contribution of ultramicrobacteria to biology. Microbiology 2012, 81, 379–390. [Google Scholar] [CrossRef]
- Kuhn, E.; Ichimura, A.S.; Peng, V.; Fritsen, C.H.; Trubl, G.; Doran, P.T.; Murray, A.E. Brine assemblages of ultrasmall microbial cells within the ice cover of Lake Vida, Antarctica. Appl. Environ. Microbiol. 2014, 80, 3687–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, G.; Maimone, G.; La Ferla, R.; Azzaro, M.; Catalfamo, M.; Genovese, M.; Santisi, S.; Maldani, M.; Macrì, A.; Cappello, S. Phenotypic variations of Oleispira antarctica RB8T in different growth conditions. Curr. Microbiol. 2020, 77, 3414–3421. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M.; Michaud, L.; Caruso, G.; Lo Giudice, A.; Paranhos, R.; Cabral, A.S.; Conte, A.; Cosenza, A.; Maimone, G.; et al. Prokaryotic abundance and activity in permafrost of the Northern Victoria Land and Upper Victoria Valley (Antarctica). Microb. Ecol. 2017, 74, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Young, K.D. The selective value of bacterial shape. Microbiol. Mol. Biol. Rev. 2006, 70, 660–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, S.J.; Rappé, M.S. Evolution, diversity and molecular ecology of marine prokaryotes. In Microbial Ecology of the Ocean; Kirchman, D.L., Ed.; Wiley Interscience: New York, NY, USA, 2000; pp. 47–84. [Google Scholar]
- Glöckner, F.O.; Fuchs, B.M.; Amann, R. Bacterioplankton compositions of lakes and oceans: A first comparison based on fluorescence in situ hybridization. Appl. Environ. Microbiol. 1999, 65, 3721–3726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchman, D.L.; Yu, L.; Cottrell, M.T. Diversity and abundance of uncultured cytophaga-like bacteria in the Delaware estuary. Appl. Environ. Microbiol. 2003, 69, 6587–6596. [Google Scholar] [CrossRef] [Green Version]
- Ngugi, D.K.; Stingl, U. High-quality draft single-cell genome sequence of the NS5 Marine Group from the coastal Red Sea. Genome Announc. 2018, 6, e00565-18. [Google Scholar] [CrossRef] [Green Version]
- Díez-Vives, C.; Nielsen, S.; Sánchez, P.; Palenzuela, O.; Ferrera, I.; Sebastián, M.; Pedrós-Alió, C.; Gasol, J.M.; Acinas, S.G. Delineation of ecologically distinct units of marine Bacteroidetes in the Northwestern Mediterranean Sea. Mol. Ecol. 2019, 28, 2846–2859. [Google Scholar] [CrossRef]
- Zeigler, D.R. The Geobacillus paradox: Why is a thermophilic bacterial genus so prevalent on a mesophilic planet? Microbiology 2014, 160, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-C.; Zhu, Z.; Li, X.; Guan, Z.; Zheng, J. Comparative genomics of Exiguobacterium reveals what makes a cosmopolitan bacterium. bioRxiv 2020. [Google Scholar] [CrossRef]
- Hahn, M.W.; Kasalicky, V.; Jezbera, J.; Brandt, U.; Jezberova, J.; Simek, K. Limnohabitans curvus gen. nov., sp nov., a planktonic bacterium isolated from a freshwater lake. Int. J. Syst. Evol. Microbiol. 2010, 60, 1358–1365. [Google Scholar] [CrossRef]
- Jezberová, J.; Jezbera, J.; Brandt, U.; Lindström, E.S.; Langenheder, S.; Hahn, M.W. Ubiquity of Polynucleobacter necessarius ssp. asymbioticus in lentic freshwater habitats of a heterogenous 2000 km2 area. Environ. Microbiol. 2010, 12, 658–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, J.; Lee, H.; Kim, M.; Kim, D.-U.; Ka, J.-O. Massilia aromaticivorans sp. nov., a BTEX degrading bacterium isolated from Arctic soil. Curr. Microbiol. 2021, 78, 2143–2150. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Wang, S.; Zhao, B.; Lou, K.; Xing, X.-H. Massilia violaceinigra sp. nov., a novel purple-pigmented bacterium isolated from glacier permafrost. Int. J. Syst. Evol. Microbiol. 2018, 68, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Dahal, R.H.; Chaudhary, D.K.; Kim, D.-U.; Kim, J. Hymenobacter polaris sp. nov., a psychrotolerant bacterium isolated from an Arctic station. Int. J. Syst. Evol. Microbiol. 2020, 70, 4890–4896. [Google Scholar] [CrossRef]
- Liu, K.; Liu, Y.; Wang, N.; Gu, Z.; Shen, L.; Xu, B.; Zhou, Y.; Liu, H.; Jiao, N. Hymenobacter glacieicola sp. nov., isolated from glacier ice. Int. J. Syst. Evol. Microbiol. 2016, 66, 3793–3798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Niu, F.; Busse, H.-J.; Ma, X.; Liu, W.; Dong, M.; Feng, H.; An, L.; Cheng, G. Hymenobacter psychrotolerans sp. nov., isolated from the Qinghai--Tibet Plateau permafrost region. Int. J. Syst. Evol. Microbiol. 2008, 58, 1215–1220. [Google Scholar] [CrossRef] [Green Version]
- Choidash, B.; Begum, Z.; Shivaji, S. Bacterial diversity of Ny-Ålesund, Arctic Archipelago Svalbard. Mong. J. Biol. Sci. 2012, 10, 67–72. [Google Scholar]
- Yang, G.L.; Hou, S.G.; Le Baoge, R.; Li, Z.G.; Xu, H.; Liu, Y.P.; Du, W.T.; Liu, Y.Q. Differences in bacterial diversity and communities between glacial snow and glacial soil on the Chongce ice cap, West Kunlun Mountains. Sci. Rep. 2016, 6, 36548. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | Riverine Stations | Brackish Stations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | ||
| Temperature | Ice-melt (−) | 5.5 | 3.7 | 3.6 | 2.7 | 5.5 | 3.6 | 3.2 | 4.4 | 4.0 |
| Ice-melt (+) | 19.1 | 20.6 | 17.2 | 15.0 | 14.7 | 13.0 | 14.0 | 10.4 | 15.4 | |
| Salinity | Ice-melt (−) | 2.0 | 3.0 | 6.0 | 4.0 | 4.0 | 15.0 | 5.0 | 15.0 | 16.0 |
| Ice-melt (+) | 0.0 | 0.0 | 0.0 | 0.0 | 1.0 | 15.0 | 6.0 | 30.0 | 17.0 | |
| TDS * (ppm) | Ice-melt (−) | 23.4 | 28.3 | 35.8 | 18.5 | 255.0 | 8.6 | 1.6 (ppt) | 7.5 (ppt) | 13.0 (ppt) |
| Ice-melt (+) | 19.2 | 173.0 | 228.0 | 19.4 | 2.6 | 13.5 | 4.5 | 19.7 | 12.9 | |
| pH | Ice-melt (−) | 5.9 | 6.4 | 6.4 | 7.0 | 6.0 | 7.6 | 6.8 | 7.7 | 7.2 |
| Ice-melt (+) | 6.3 | 7.6 | 7.2 | 7.3 | 5.9 | 7.8 | 7.1 | 8.1 | 8.3 | |
| O2 | Ice-melt (−) | 87.5 | 70.5 | 68 | 74.9 | 88.3 | 93.1 | 88.5 | 101.6 | 102.2 |
| Ice-melt (+) | 83.2 | 107.9 | 93.9 | 94 | 82.2 | 96.7 | 93.8 | 95 | 88.2 | |
| Redox (mV) | Ice-melt (−) | 442.0 | 175.0 | 178.0 | 134.0 | 252.0 | 125.0 | 224.0 | 140.0 | 82.0 |
| Ice-melt (+) | 252.0 | 246.0 | 198.0 | 207.0 | 110.0 | 37.0 | 93.0 | 86.0 | 81.0 | |
| Parameter * | Riverine Stations | Brackish Stations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | |||
| Particulate matter | TSM (mg L−1) | Ice-melt (−) | 1.75 | 3.2 | 4.15 | 0.35 | 1.175 | 5.07 | 5.35 | 15.25 | 2.10 |
| Ice-melt (+) | 1.49 | 5.57 | 6.725 | 33.53 | 14.42 | 0.98 | 1.49 | 1.26 | 33.30 | ||
| POC (µg L−1) | Ice-melt (−) | 243.61 | 504.78 | 594.95 | 153.38 | 221.31 | 225.76 | 583.96 | 560.09 | 211.04 | |
| Ice-melt (+) | 242.25 | 819.49 | 722.39 | 725.72 | 740.90 | 120.70 | 159.67 | 228.33 | 1443.75 | ||
| δ13 POC (‰) | Ice-melt (−) | −26.66 | −29.69 | −30.14 | −31.66 | −31.58 | −27.48 | −28.04 | −27.26 | −26.43 | |
| Ice-melt (+) | −27.78 | −25.62 | −24.53 | −28.39 | −25.11 | −25.22 | −25.60 | −24.27 | −25.77 | ||
| PN tot (µg L−1) | Ice-melt (−) | 43.19 | 90.93 | 128.86 | 27.72 | 36.95 | 36.91 | 102.47 | 95.37 | 33.38 | |
| Ice-melt (+) | 47.65 | 161.54 | 119.15 | 90.90 | 102.49 | 25.55 | 31.93 | 41.40 | 197.52 | ||
| Ctot (µg L−1) | Ice-melt (−) | 314.84 | 571.96 | 717.68 | 180.18 | 228.76 | 226.77 | 715.02 | 680.85 | 213.87 | |
| Ice-melt (+) | 271.98 | 956.08 | 845.23 | 780.22 | 748.33 | 134.33 | 194.15 | 235.33 | 1525.50 | ||
| PIC (µg) | Ice-melt (−) | 71.23 | 67.18 | 122.72 | 26.80 | 7.45 | 1.01 | 131.07 | 120.76 | 2.84 | |
| Ice-melt (+) | 29.73 | 136.59 | 122.83 | 54.50 | 7.43 | 13.63 | 34.47 | 7.01 | 81.75 | ||
| POC (%) | Ice-melt (−) | 13.92 | 15.77 | 14.34 | 43.82 | 18.84 | 4.45 | 10.92 | 3.67 | 10.05 | |
| Ice-melt (+) | 16.25 | 14.70 | 10.74 | 2.16 | 5.14 | 12.27 | 10.72 | 18.12 | 4.34 | ||
| PN tot (%) | Ice-melt (−) | 2.47 | 2.84 | 3.11 | 7.92 | 3.14 | 0.73 | 1.92 | 0.63 | 1.59 | |
| Ice-melt (+) | 3.20 | 2.90 | 1.77 | 0.27 | 0.71 | 2.60 | 2.14 | 3.29 | 0.59 | ||
| PIC (%) | Ice-melt (−) | 4.07 | 2.10 | 2.96 | 7.66 | 0.63 | 0.02 | 2.45 | 0.79 | 0.14 | |
| Ice-melt (+) | 1.99 | 2.45 | 1.83 | 0.16 | 0.05 | 1.39 | 2.31 | 0.56 | 0.25 | ||
| C/N molar/ratio | Ice-melt (−) | 6.58 | 6.48 | 5.39 | 6.45 | 6.99 | 7.14 | 6.65 | 6.85 | 7.38 | |
| Ice-melt (+) | 5.93 | 5.92 | 7.07 | 9.31 | 8.43 | 5.51 | 5.83 | 6.43 | 8.53 | ||
| Chlorophyll-a content | Chl-a (µg L−1) | Ice-melt (−) | 1.609 | 0.953 | 1.961 | 0.828 | 1.181 | 0.951 | 2.347 | 1.432 | 0.638 |
| Ice-melt (+) | 1.495 | 1.298 | 1.027 | 0.826 | 0.854 | 1.179 | 0.754 | 0.455 | 1.860 | ||
| Pheo (µg L−1) | Ice-melt (−) | 0.519 | 0.464 | 0.796 | 0.150 | 0.417 | 0.426 | 0.673 | 0.196 | 0.824 | |
| Ice-melt (+) | 0.549 | 1.970 | 0.345 | 0.457 | 0.643 | 0.900 | 0.313 | 0.287 | 0.728 | ||
| Dissolved nutrients | PO43− (µM) | Ice-melt (−) | 0.061 | 0.704 | 0.861 | 0.282 | 0.078 | 0.164 | 0.080 | 0.086 | 0.091 |
| Ice-melt (+) | 0.013 | 0.055 | 0.032 | 0.029 | 0.013 | 0.056 | 0.027 | 0.026 | 0.017 | ||
| NH4+ (µM) | Ice-melt (−) | 2.565 | 1.482 | 1.7 | 0.517 | 0.464 | 1.866 | 0.651 | 1.407 | 0.783 | |
| Ice-melt (+) | 1.760 | 0.828 | 0.817 | 0.449 | 1.403 | 6.820 | 0.909 | 0.840 | 3.554 | ||
| NO3− (µM) | Ice-melt (−) | 0.157 | 0.023 | 0.487 | 2.171 | 2.494 | 0.474 | 0.451 | 0.8 | 0.335 | |
| Ice-melt (+) | 0.435 | 0.448 | 0.656 | 0.288 | 0.208 | 1.179 | 0.253 | 0.392 | 0.462 | ||
| NO2− (µM) | Ice-melt (−) | 0.001 | 0.011 | 0.023 | 0.055 | 0.001 | 0.026 | 0.001 | 0.015 | 0.024 | |
| Ice-melt (+) | 0.021 | 0.039 | 0.032 | 0.051 | 0.012 | 0.108 | 0.002 | 0.002 | 0.013 | ||
| Microbial Abundance | Riverine Stations | Brackish Stations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| St.9 | St.5 | St.1 | St.2 | St.8 | St.3 | St.7 | St.6 | St.4 | ||
| Heterotrophs (Prokaryotes) (105 cells mL−1) | Ice-melt (−) | 2.09 | 8.56 | 10.3 | 1.42 | 3.77 | 4.63 | 5.13 | 6.79 | 3.57 |
| Ice-melt (+) | 5.87 | 15.3 | 5.54 | 4.15 | 6.53 | 5.55 | 3.94 | 3.64 | 7.15 | |
| Heterotrophs (nano-plankton) (104 cells mL−1) | Ice-melt (−) | 1.24 | 2.46 | 2.93 | 1.30 | 0.97 | 1.51 | 1.94 | 1.02 | 0.79 |
| Ice-melt (+) | 1.24 | 1.37 | 1.41 | 0.99 | 1.49 | 1.68 | 0.86 | 1.22 | 1.33 | |
| Autotrophs (PE-rich Cyanobacteria) (103 cells mL−1) | Ice-melt (−) | 0.39 | 3.27 | 43.6 | 3.87 | 0.45 | 15.5 | 1.74 | 1.66 | 3.20 |
| Ice-melt (+) | 0.58 | 0.37 | 0.18 | 0.48 | 0.28 | 0.03 | 0.09 | 0.01 | 0.07 | |
| Autotrophs (PC-rich Cyanobacteria) (102 cells mL−1) | Ice-melt (−) | 0.70 | 0.56 | 14.2 | 1.47 | 0.68 | 2.40 | 3.85 | 0.89 | 0.56 |
| Ice-melt (+) | 0.40 | 0.74 | 0.59 | 0.40 | 0.26 | 0.11 | 0.11 | 0.14 | 0.26 | |
| Autotrophs (pico-eukaryotes) (102 cells mL−1) | Ice-melt (−) | 2.79 | 3.90 | 66.9 | 3.01 | 2.40 | 15.4 | 26.7 | 5.06 | 2.81 |
| Ice-melt (+) | 1.82 | 4.54 | 3.22 | 2.42 | 1.26 | 1.05 | 0.55 | 0.51 | 2.27 | |
| Autotrophs (nano-plankton) (102 cells mL−1) | Ice-melt (−) | 0.74 | 1.85 | 17.7 | 1.17 | 0.52 | 14.9 | 4.22 | 3.68 | 3.00 |
| Ice-melt (+) | 0.59 | 2.53 | 3.00 | 1.34 | 0.37 | 0.63 | 0.25 | 0.59 | 6.07 | |
| Morphometric Traits | Riverine Stations | Brackish Stations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | ||
| Mean lenght (µm) | Ice-melt (−) | 0.47 | 0.55 | 0.55 | 0.56 | 0.52 | 0.62 | 0.48 | 0.61 | 0.63 |
| Ice-melt (+) | 0.45 | 0.49 | 0.94 | 0.64 | 0.44 | 0.59 | 0.65 | 0.78 | 0.68 | |
| Mean width (µm) | Ice-melt (−) | 0.31 | 0.34 | 0.29 | 0.32 | 0.31 | 0.32 | 0.32 | 0.30 | 0.34 |
| Ice-melt (+) | 0.28 | 0.28 | 0.26 | 0.30 | 0.27 | 0.26 | 0.30 | 0.29 | 0.29 | |
| Mean volume (µm3) | Ice-melt (−) | 0.03 | 0.04 | 0.03 | 0.04 | 0.04 | 0.04 | 0.03 | 0.04 | 0.04 |
| Ice-melt (+) | 0.02 | 0.02 | 0.04 | 0.04 | 0.02 | 0.03 | 0.04 | 0.05 | 0.04 | |
| CCC (fg C cell−1) | Ice-melt (−) | 10.7 | 13.7 | 10.6 | 12.1 | 12.4 | 13.1 | 11.1 | 13.7 | 14.7 |
| Ice-melt (+) | 7.9 | 8.9 | 14.5 | 13.1 | 7.5 | 9.1 | 14.2 | 15.3 | 13.2 | |
| Cocci (%) | Ice-melt (−) | 71.2 | 71.2 | 52.9 | 56.3 | 71.7 | 47.4 | 66.7 | 56.4 | 50.5 |
| Ice-melt (+) | 68.5 | 68.3 | 32.1 | 65.0 | 68.5 | 57.1 | 53.0 | 39.9 | 51.2 | |
| Rods (%) | Ice-melt (−) | 16.9 | 13.6 | 25.9 | 26.4 | 11.3 | 27.8 | 19.4 | 24.8 | 25.2 |
| Ice-melt (+) | 9.3 | 11.7 | 14.9 | 9.4 | 17.4 | 4.8 | 28.0 | 14.0 | 15.2 | |
| Vibrios (%) | Ice-melt (−) | 5.1 | 3.0 | 4.7 | 6.9 | 5.7 | 12.4 | 2.2 | 2.6 | 6.8 |
| Ice-melt (+) | 3.7 | 8.3 | 17.9 | 6.0 | 2.2 | 1.9 | 6.8 | 16.9 | 14.4 | |
| Spirillae (%) | Ice-melt (−) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.7 | 1.9 |
| Ice-melt (+) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.8 | 0.0 | |
| Coccobacilli (%) | Ice-melt (−) | 3.4 | 3.0 | 10.6 | 8.0 | 7.5 | 7.2 | 6.5 | 6.8 | 10.7 |
| Ice-melt (+) | 5.6 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 3.8 | 4.5 | 2.4 | |
| Curved rods(%) | Ice-melt (−) | 3.4 | 9.1 | 5.9 | 2.3 | 3.8 | 5.2 | 5.4 | 7.7 | 4.9 |
| Ice-melt (+) | 13.0 | 11.7 | 35.1 | 19.7 | 7.6 | 36.2 | 8.3 | 21.9 | 16.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papale, M.; Rizzo, C.; Caruso, G.; Amalfitano, S.; Maimone, G.; Miserocchi, S.; La Ferla, R.; Aspholm, P.E.; Decembrini, F.; Azzaro, F.; et al. Ice Melt-Induced Variations of Structural and Functional Traits of the Aquatic Microbial Community along an Arctic River (Pasvik River, Norway). Water 2021, 13, 2297. https://doi.org/10.3390/w13162297
Papale M, Rizzo C, Caruso G, Amalfitano S, Maimone G, Miserocchi S, La Ferla R, Aspholm PE, Decembrini F, Azzaro F, et al. Ice Melt-Induced Variations of Structural and Functional Traits of the Aquatic Microbial Community along an Arctic River (Pasvik River, Norway). Water. 2021; 13(16):2297. https://doi.org/10.3390/w13162297
Chicago/Turabian StylePapale, Maria, Carmen Rizzo, Gabriella Caruso, Stefano Amalfitano, Giovanna Maimone, Stefano Miserocchi, Rosabruna La Ferla, Paul Eric Aspholm, Franco Decembrini, Filippo Azzaro, and et al. 2021. "Ice Melt-Induced Variations of Structural and Functional Traits of the Aquatic Microbial Community along an Arctic River (Pasvik River, Norway)" Water 13, no. 16: 2297. https://doi.org/10.3390/w13162297