Lake Phytoplankton Assemblage Altered by Irregularly Shaped PLA Body Wash Microplastics but Not by PS Calibration Beads


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
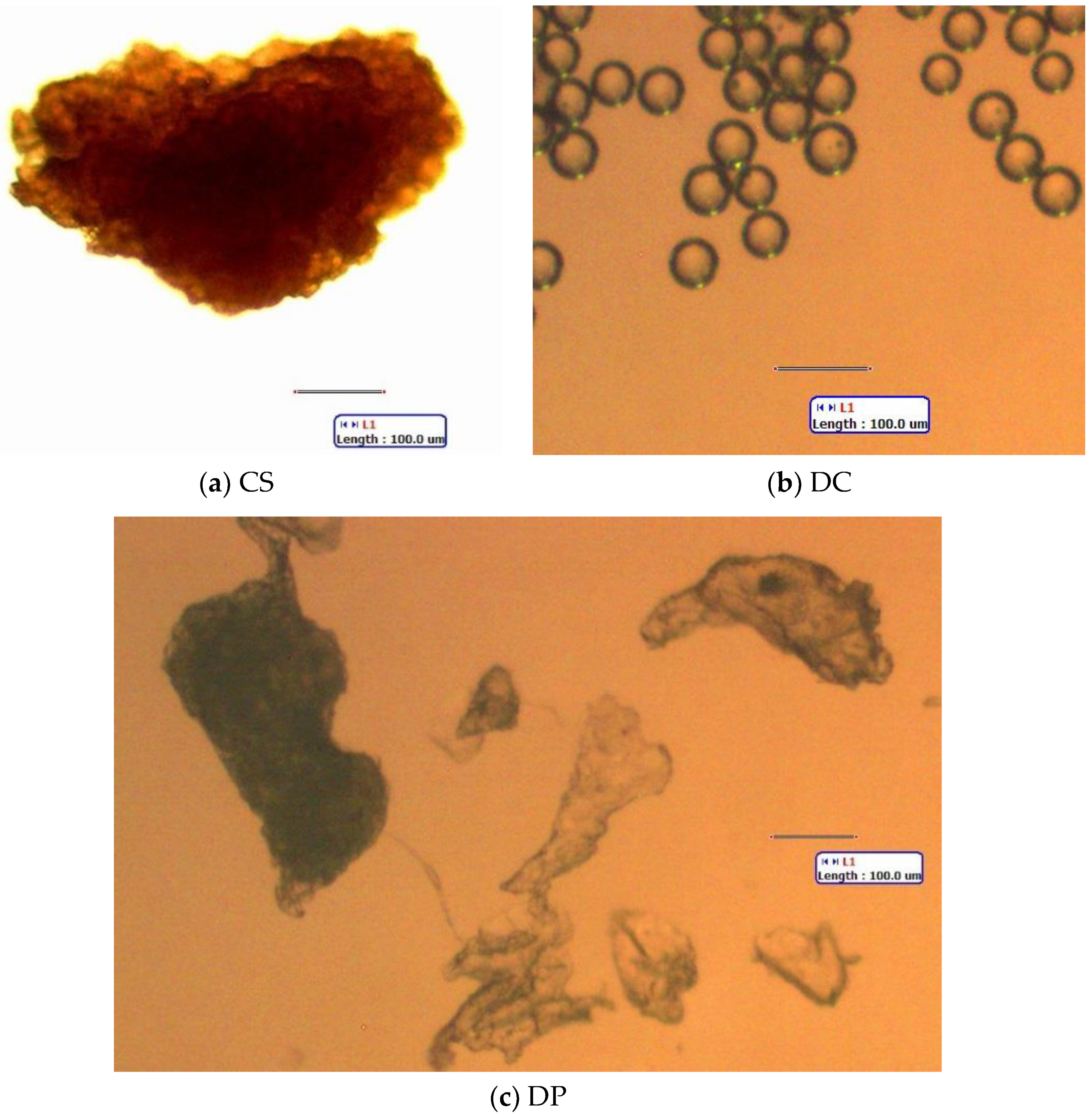
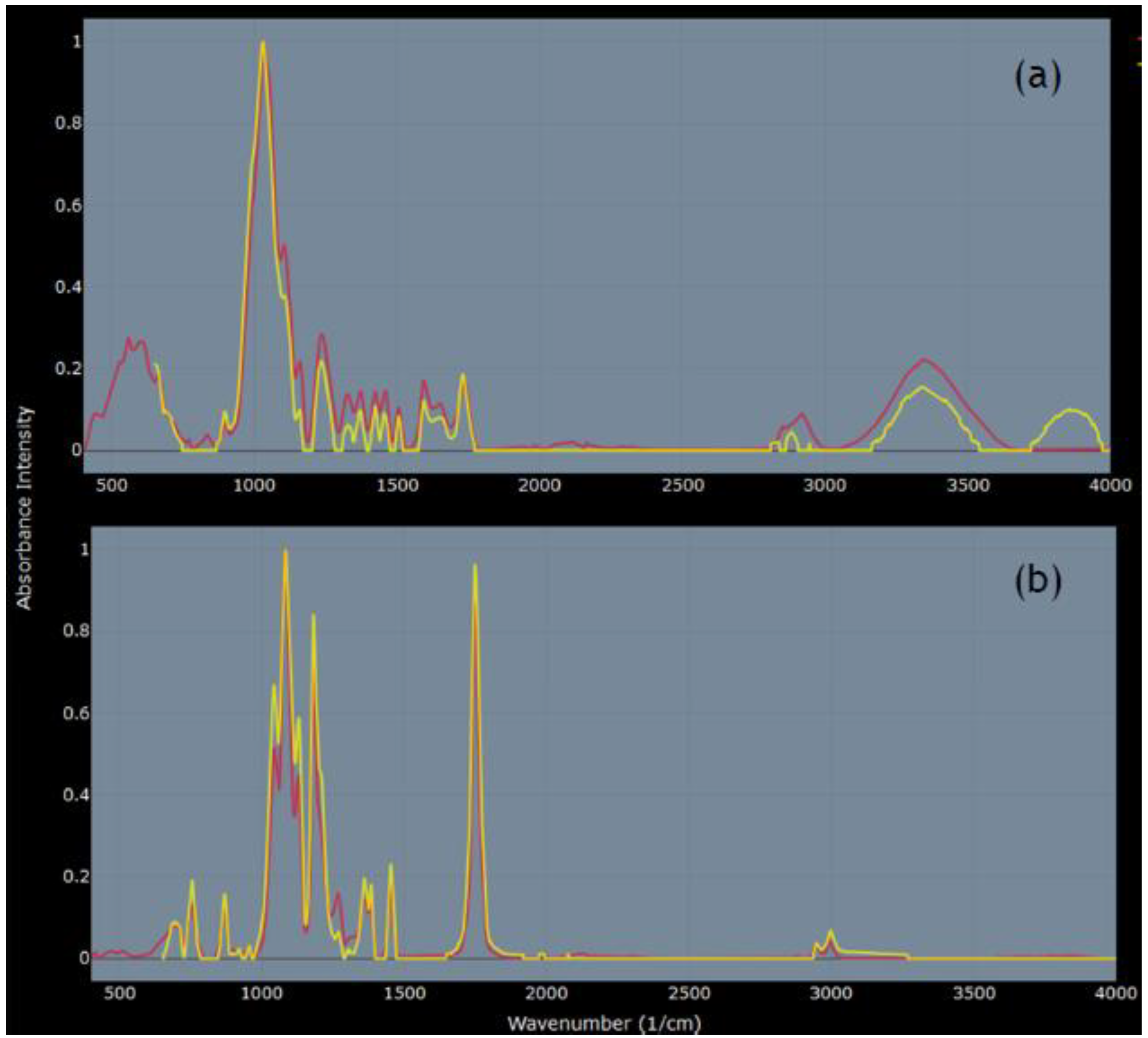
3.1. FTIR Analysis of Body Wash Particles
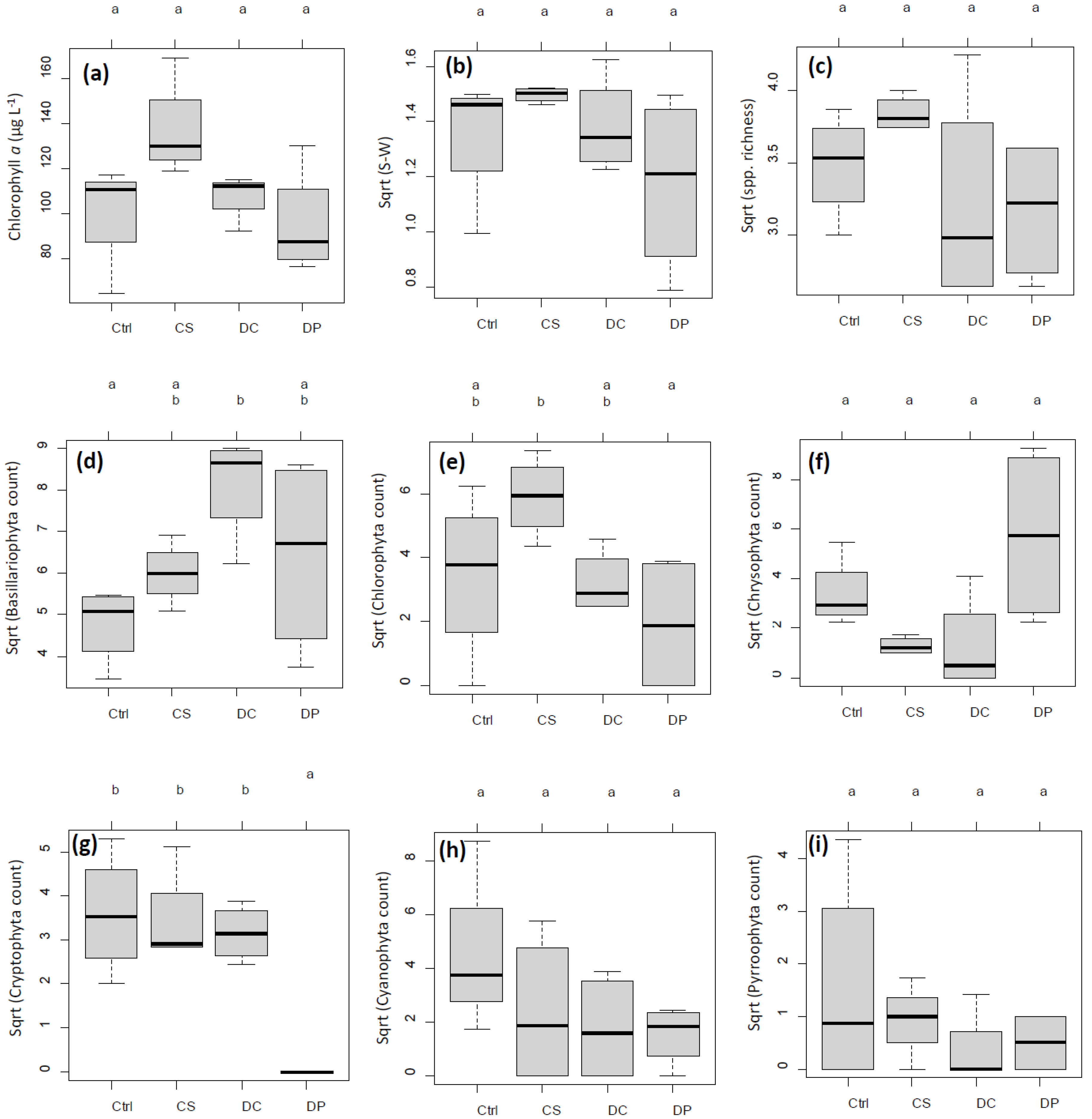
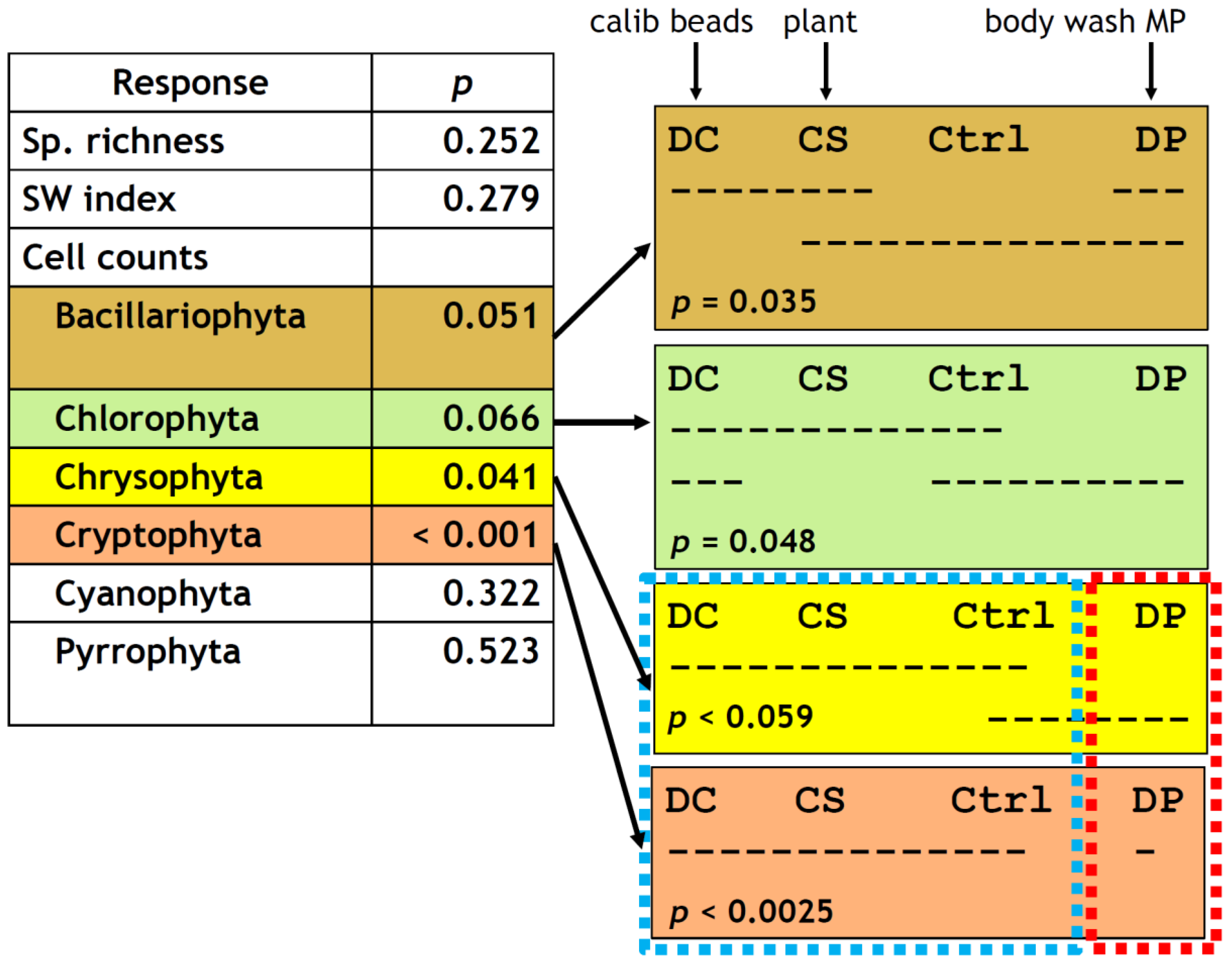
3.2. Chl. a and Diversity Indices
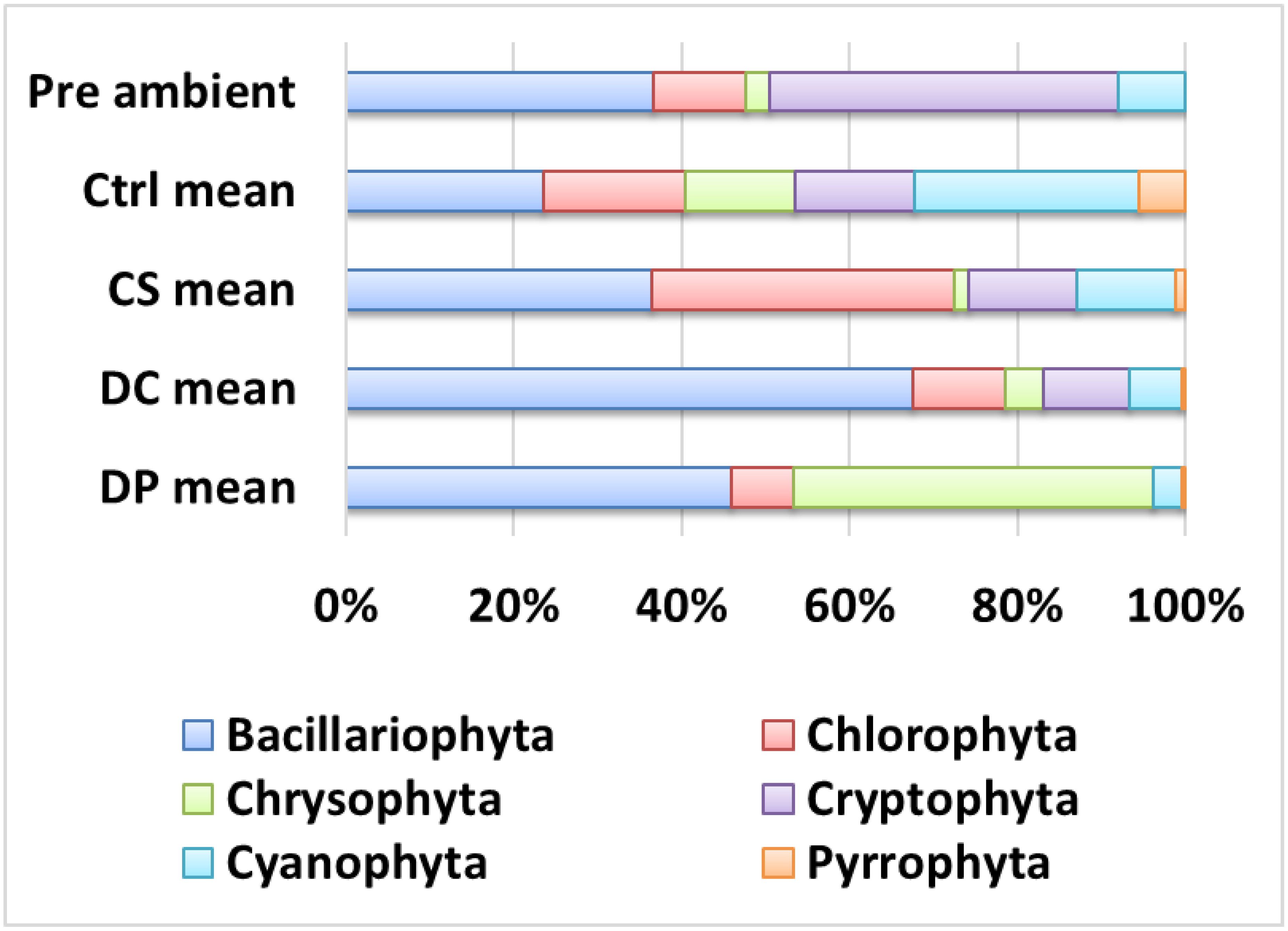
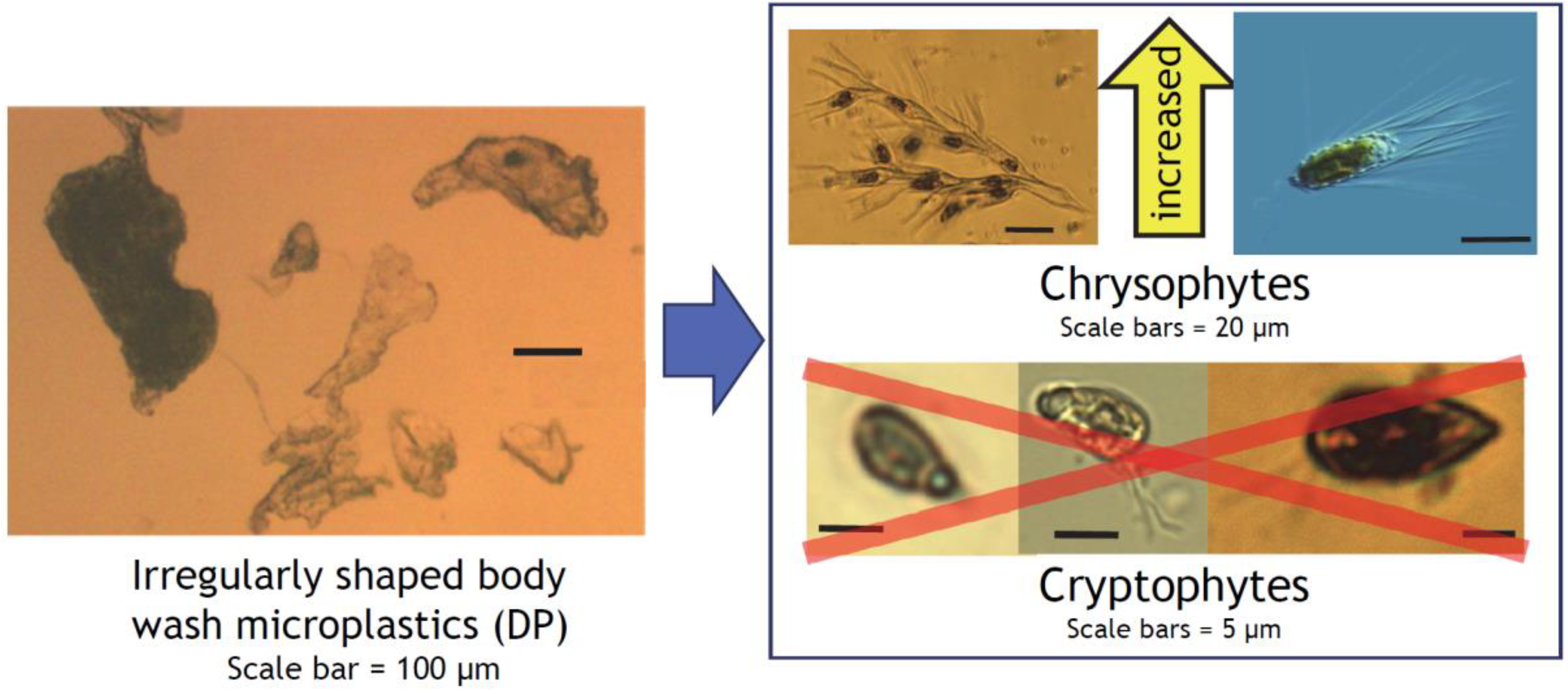
3.3. Phytoplankton Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carpenter, E.J.; Smith, K.L.; Burke, J.A.; Schubert, W.K. Plastics on the Sargasso Sea Surface. Science 1972, 175, 1240–1241. [Google Scholar] [CrossRef]
- Andrady, A.L. Microplastics in the marine environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Sul, J.A.I.D.; Costa, M.F. The present and future of microplastic pollution in the marine environment. Environ. Pollut. 2014, 185, 352–364. [Google Scholar] [CrossRef]
- Ivleva, N.P.; Wiesheu, A.C.; Niessner, R. Microplastic in Aquatic Ecosystems. Angew. Chem. Int. Ed. 2016, 56, 1720–1739. [Google Scholar] [CrossRef]
- Rochman, C.M. Microplastics research—from sink to source. Science 2018, 360, 28–29. [Google Scholar] [CrossRef]
- Duis, K.; Coors, A. Microplastics in the aquatic and terrestrial environment: Sources (with a specific focus on personal care products), fate and effects. Environ. Sci. Eur. 2016, 28, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Van Sebille, E.; Wilcox, C.; Lebreton, L.; Maximenko, N.; Hardesty, B.D.; Van Franeker, J.A.; Eriksen, M.; Siegel, D.; Galgani, F.; Law, K.L. A global inventory of small floating plastic debris. Environ. Res. Lett. 2015, 10, 124006. [Google Scholar] [CrossRef]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R.C. Accumulation of Microplastic on Shorelines Woldwide: Sources and Sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef]
- Hoellein, T.; McCormick, A.R.; Hittie, J.; London, M.G.; Scott, J.W.; Kelly, J.J. Longitudinal patterns of microplastic concentration and bacterial assemblages in surface and benthic habitats of an urban river. Freshw. Sci. 2017, 36, 491–507. [Google Scholar] [CrossRef]
- McCormick, A.; Hoellein, T.J.; Mason, S.A.; Schluep, J.; Kelly, J.J. Microplastic is an Abundant and Distinct Microbial Habitat in an Urban River. Environ. Sci. Technol. 2014, 48, 11863–11871. [Google Scholar] [CrossRef]
- McCormick, A.R.; Hoellein, T.; London, M.G.; Hittie, J.; Scott, J.W.; Kelly, J.J. Microplastic in surface waters of urban rivers: Concentration, sources, and associated bacterial assemblages. Ecosphere 2016, 7. [Google Scholar] [CrossRef]
- Smith, E.; Dziewatkoski, M.; Henrie, T.; Seidel, C.; Rosen, J. Microplastics: What Drinking Water Utilities Need to Know. J. Am. Water Work. Assoc. 2019, 111, 26–37. [Google Scholar] [CrossRef]
- Mao, Y.; Ai, H.; Chen, Y.; Zhang, Z.; Zeng, P.; Kang, L.; Li, W.; Gu, W.; He, Q.; Li, H. Phytoplankton response to polystyrene microplastics: Perspective from an entire growth period. Chemosphere 2018, 208, 59–68. [Google Scholar] [CrossRef]
- Arias-Andres, M.; Klümper, U.; Rojas-Jimenez, K.; Grossart, H.-P. Microplastic pollution increases gene exchange in aquatic ecosystems. Environ. Pollut. 2018, 237, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Triebskorn, R.; Braunbeck, T.; Grummt, T.; Hanslik, L.; Huppertsberg, S.; Jekel, M.; Knepper, T.P.; Krais, S.; Müller, Y.K.; Pittroff, M.; et al. Relevance of nano- and microplastics for freshwater ecosystems: A critical review. TrAC Trends Anal. Chem. 2019, 110, 375–392. [Google Scholar] [CrossRef]
- Yokota, K.; Waterfield, H.; Hastings, C.; Davidson, E.; Kwietniewski, E.; Wells, B. Finding the missing piece of the aquatic plastic pollution puzzle: Interaction between primary producers and microplastics. Limnol. Oceanogr. Lett. 2017, 2, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Oberbeckmann, S.; Löder, M.G.J.; Labrenz, M. Marine microplastic-associated biofilms—A review. Environ. Chem. 2015, 12, 551–562. [Google Scholar] [CrossRef]
- Miao, L.; Hou, J.; You, G.; Liu, Z.; Liu, S.; Li, T.; Mo, Y.; Guo, S.; Qu, H. Acute effects of nanoplastics and microplastics on periphytic biofilms depending on particle size, concentration and surface modification. Environ. Pollut. 2019, 255, 113300. [Google Scholar] [CrossRef]
- Fendall, L.S.; Sewell, M.A. Contributing to marine pollution by washing your face: Microplastics in facial cleansers. Mar. Pollut. Bull. 2009, 58, 1225–1228. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.; Hastings, C.; Waterfield, H.; Yokota, K. Development of Methods to Characterize & Extract Plastic Microparticles from Personal Cleansing Products; 47th Annual Report; Biological Field Station: Cooperstown, NY, USA, 2014; pp. 142–148. [Google Scholar]
- Kalčíková, G.; Alič, B.; Skalar, T.; Bundschuh, M.; Gotvajn, A. Žgajnar Wastewater treatment plant effluents as source of cosmetic polyethylene microbeads to freshwater. Chemosphere 2017, 188, 25–31. [Google Scholar] [CrossRef]
- Free, C.M.; Jensen, O.P.; Mason, S.A.; Eriksen, M.; Williamson, N.; Boldgiv, B. High-levels of microplastic pollution in a large, remote, mountain lake. Mar. Pollut. Bull. 2014, 85, 156–163. [Google Scholar] [CrossRef]
- Barrows, A.P.; Cathey, S.; Petersen, C. Marine environment microfiber contamination: Global patterns and the diversity of microparticle origins. Environ. Pollut. 2018, 237, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Rochman, C.M.; Kross, S.M.; Armstrong, J.B.; Bogan, M.T.; Darling, E.S.; Green, S.J.; Smyth, A.R.; Veríssimo, D. Scientific Evidence Supports a Ban on Microbeads. Environ. Sci. Technol. 2015, 49, 10759–10761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerranti, C.; Martellini, T.; Perra, G.; Scopetani, C.; Cincinelli, A. Microplastics in cosmetics: Environmental issues and needs for global bans. Environ. Toxicol. Pharmacol. 2019, 68, 75–79. [Google Scholar] [CrossRef]
- Lewis, A.S.L.; Kim, B.S.; Edwards, H.L.; Wander, H.L.; Garfield, C.M.; Murphy, H.E.; Poulin, N.D.; Princiotta, S.D.; Rose, K.C.; Taylor, A.E.; et al. Prevalence of phytoplankton limitation by both nitrogen and phosphorus related to nutrient stoichiometry, land use, and primary producer biomass across the northeastern United States. Inland Waters 2020, 10, 42–50. [Google Scholar] [CrossRef]
- De Witte, B.; Devriese, L.; Bekaert, K.; Hoffman, S.; Vandermeersch, G.; Cooreman, K.; Robbens, J. Quality assessment of the blue mussel (Mytilus edulis): Comparison between commercial and wild types. Mar. Pollut. Bull. 2014, 85, 146–155. [Google Scholar] [CrossRef]
- Cowger, W.; Gray, A.; Hapich, H.; Rochman, C.; Lynch, J.; Primpke, S.; Munno, K.; De Frond, H.; Herodotou, O. Open Specy. Available online: www.openspecy.org (accessed on 20 September 2020).
- Arar, E.J.; Collins, G.B. Method 445.0 in Vitro Determination of Chlorophyll a and Pheophytin a in Marine and Freshwater Algae by Fluorescence; USEPA: Cincinnati, OH, USA, 1997; p. 22.
- Mehlrose, M.; Yokota, K. Evaluation of Chlorophyll a Extraction Techniques; State Universtiy of New York College at Oneonta Biological Field Station Annual Report; State Universtiy of New York College at Oneonta Biological Field Station: Cooperstown, NY, USA, 2016; pp. 66–75. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- R Studio Team. R Studio: Integrated Development for R; PBC: Boston, MA, USA, 2020. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Graves, S.; Piepho, H.-P.; Selzer, L. multcompView: Visualizations of Paired Comparisons. Available online: https://cran.r-project.org/web/packages/multcompView/index.html (accessed on 20 September 2020).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Primpke, S.; Wirth, M.; Lorenz, C.; Gerdts, G. Reference database design for the automated analysis of microplastic samples based on Fourier transform infrared (FTIR) spectroscopy. Anal. Bioanal. Chem. 2018, 410, 5131–5141. [Google Scholar] [CrossRef] [Green Version]
- Long, M.; Moriceau, B.; Gallinari, M.; Lambert, C.; Huvet, A.; Raffray, J.; Soudant, P. Interactions between microplastics and phytoplankton aggregates: Impact on their respective fates. Mar. Chem. 2015, 175, 39–46. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Lin, S.; Turner, J.P.; Ke, P.C. Physical Adsorption of Charged Plastic Nanoparticles Affects Algal Photosynthesis. J. Phys. Chem. C 2010, 114, 16556–16561. [Google Scholar] [CrossRef]
- Sjollema, S.B.; Redondo-Hasselerharm, P.; Leslie, H.; Kraak, M.H.S.; Vethaak, A.D. Do plastic particles affect microalgal photosynthesis and growth? Aquat. Toxicol. 2016, 170, 259–261. [Google Scholar] [CrossRef]
- Feng, L.-J.; Sun, X.-D.; Zhu, F.-P.; Feng, Y.; Duan, J.-L.; Xiao, F.; Li, X.-Y.; Shi, Y.; Wang, Q.; Sun, J.-W.; et al. Nanoplastics Promote Microcystin Synthesis and Release from Cyanobacterial Microcystis aeruginosa. Environ. Sci. Technol. 2020, 54, 3386–3394. [Google Scholar] [CrossRef]
- Smith, S.A. Beyond Toxins: A Source-to-Treatment Strategy for Harmful Algal Blooms. J. Am. Water Work. Assoc. 2019, 111, 14–22. [Google Scholar] [CrossRef]
- Casalini, T.; Rossi, F.; Castrovinci, A.; Perale, G. A Perspective on Polylactic Acid-Based Polymers Use for Nanoparticles Synthesis and Applications. Front. Bioeng. Biotechnol. 2019, 7, 259. [Google Scholar] [CrossRef]
- Madival, S.; Auras, R.; Singh, S.P.; Narayan, R. Assessment of the environmental profile of PLA, PET and PS clamshell containers using LCA methodology. J. Clean. Prod. 2009, 17, 1183–1194. [Google Scholar] [CrossRef]
- Fieschi, M.; Pretato, U. Role of compostable tableware in food service and waste management. A life cycle assessment study. Waste Manag. 2018, 73, 14–25. [Google Scholar] [CrossRef]
- Gantt, E. Micromorphology of the Periplast of Chroomonas sp. (Cryptophyceae)2. J. Phycol. 1971, 7, 177–184. [Google Scholar] [CrossRef]
- Sheath, R.G.; Wehr, J.D. Introduction to freshwater algae. In Freshwater Algae of North. America: Ecology and Classification; Academic Press: San Diego, CA, USA, 2003; pp. 1–9. ISBN 0-12-741550-5. [Google Scholar]
- Kugrens, P.; Lee, R.E. An Ultrastructural survey of Cryptomonad Periplasts Using Quick-Freezing Freeze Fracture Techniques. J. Phycol. 1987, 23, 365–376. [Google Scholar] [CrossRef]
- Siegelman, H.; Kycia, J. Algal biliproteins. In Handbook of Phycological Methods: Psychological Biochemistry Methods; Cambridge University Press: Cambridge, UK, 1978; pp. 71–79. [Google Scholar]
- Obayashi, Y.; Suzuki, S. Proteolytic enzymes in coastal surface seawater: Significant activity of endopeptidases and exopeptidases. Limnol. Oceanogr. 2005, 50, 722–726. [Google Scholar] [CrossRef]
- Kiersztyn, B.; Siuda, W.; Chróst, R.J. Persistence of bacterial proteolytic enzymes in lake ecosystems. FEMS Microbiol. Ecol. 2012, 80, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Proyecto Agua Mallomonas, el cometa entre estrellas. S.O.S. Lago de Sanabria. Available online: https://www.flickr.com/photos/25898159@N07/49646121252. (accessed on 20 September 2020).
- Kristiansen, J.; Preisig, H.R. Phylum Chrysophyta (golden algae). In The Freshwater Algal Flora of the British Isles: An. Identification Guide to Freshwater and Terrestrial Algae; Cambridge University Press: Cambridge, UK, 2011; pp. 281–317. [Google Scholar]
- Lee, R.E. Phycology; Cambridge University Press (CUP): Cambridge, UK, 2008. [Google Scholar]
- Biggs, B.J.F.; Stevenson, R.J.; Lowe, R.L. A habitat matrix conceptual model for stream periphyton. Fundam. Appl. Limnol./Archiv für Hydrobiologie 1998, 143, 21–56. [Google Scholar] [CrossRef]
- González-Pleiter, M.; Belda, M.T.; Pulido-Reyes, G.; Amariei, G.; Leganes, F.; Rosal, R.; Fernández-Piñas, F. Secondary nanoplastics released from a biodegradable microplastic severely impact freshwater environments. Environ. Sci. Nano 2019, 6, 1382–1392. [Google Scholar] [CrossRef]
- Baldwin, A.K.; Spanjer, A.R.; Rosen, M.R.; Thom, T. Microplastics in Lake Mead National Recreation Area, USA: Occurrence and biological uptake. PLoS ONE 2020, 15, e0228896. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yokota, K.; Mehlrose, M. Lake Phytoplankton Assemblage Altered by Irregularly Shaped PLA Body Wash Microplastics but Not by PS Calibration Beads. Water 2020, 12, 2650. https://doi.org/10.3390/w12092650
Yokota K, Mehlrose M. Lake Phytoplankton Assemblage Altered by Irregularly Shaped PLA Body Wash Microplastics but Not by PS Calibration Beads. Water. 2020; 12(9):2650. https://doi.org/10.3390/w12092650
Chicago/Turabian StyleYokota, Kiyoko, and Marissa Mehlrose. 2020. "Lake Phytoplankton Assemblage Altered by Irregularly Shaped PLA Body Wash Microplastics but Not by PS Calibration Beads" Water 12, no. 9: 2650. https://doi.org/10.3390/w12092650