Winter Decomposition of Emergent Macrophytes Affects Water Quality under Ice in a Temperate Shallow Lake

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
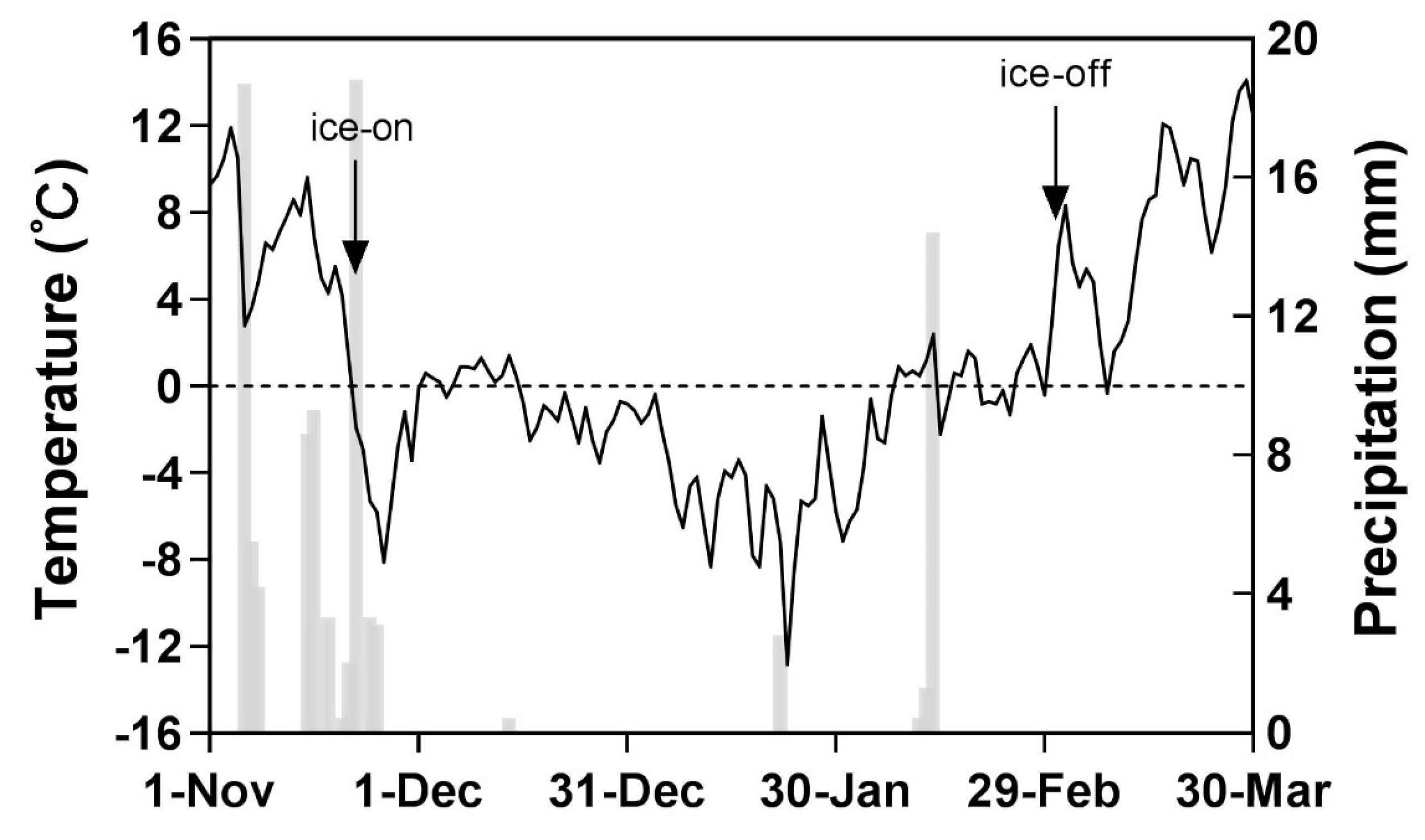
2.1. Study Area
2.2. Experiment Design
2.3. Chemical Analysis
2.4. Calculations and Statistical Analysis
3. Results
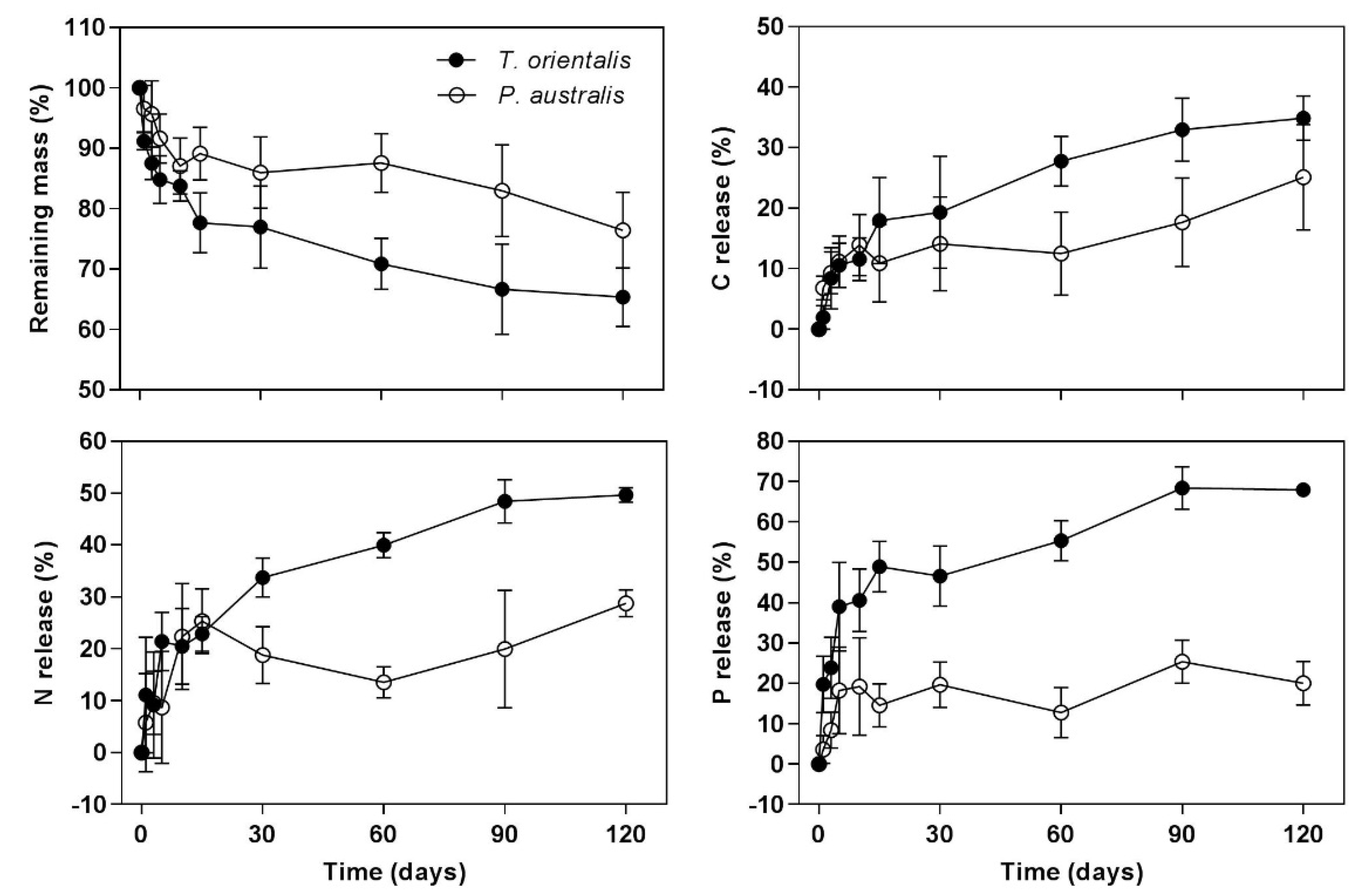
3.1. Mass Loss and C, N, and P Release
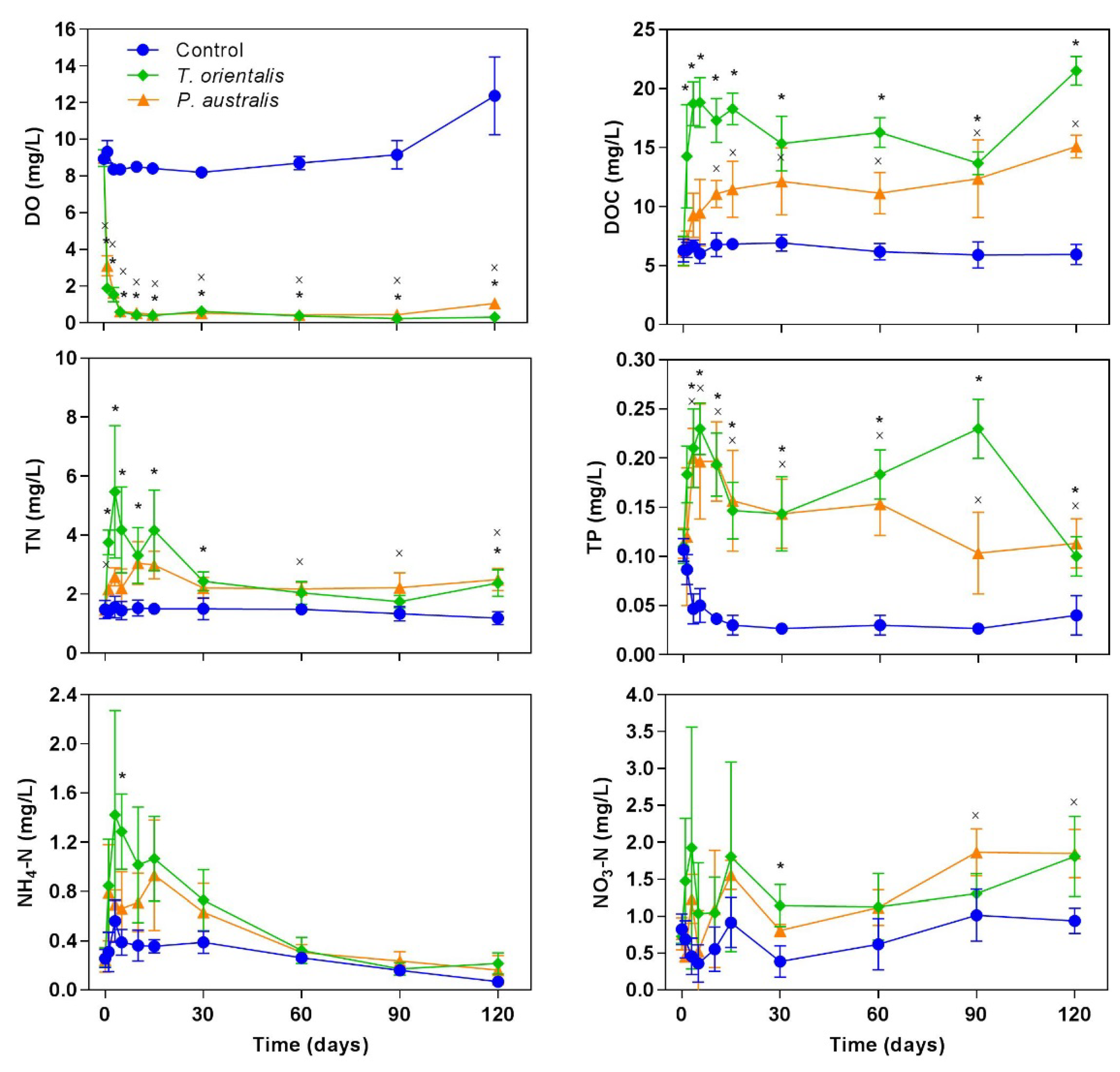
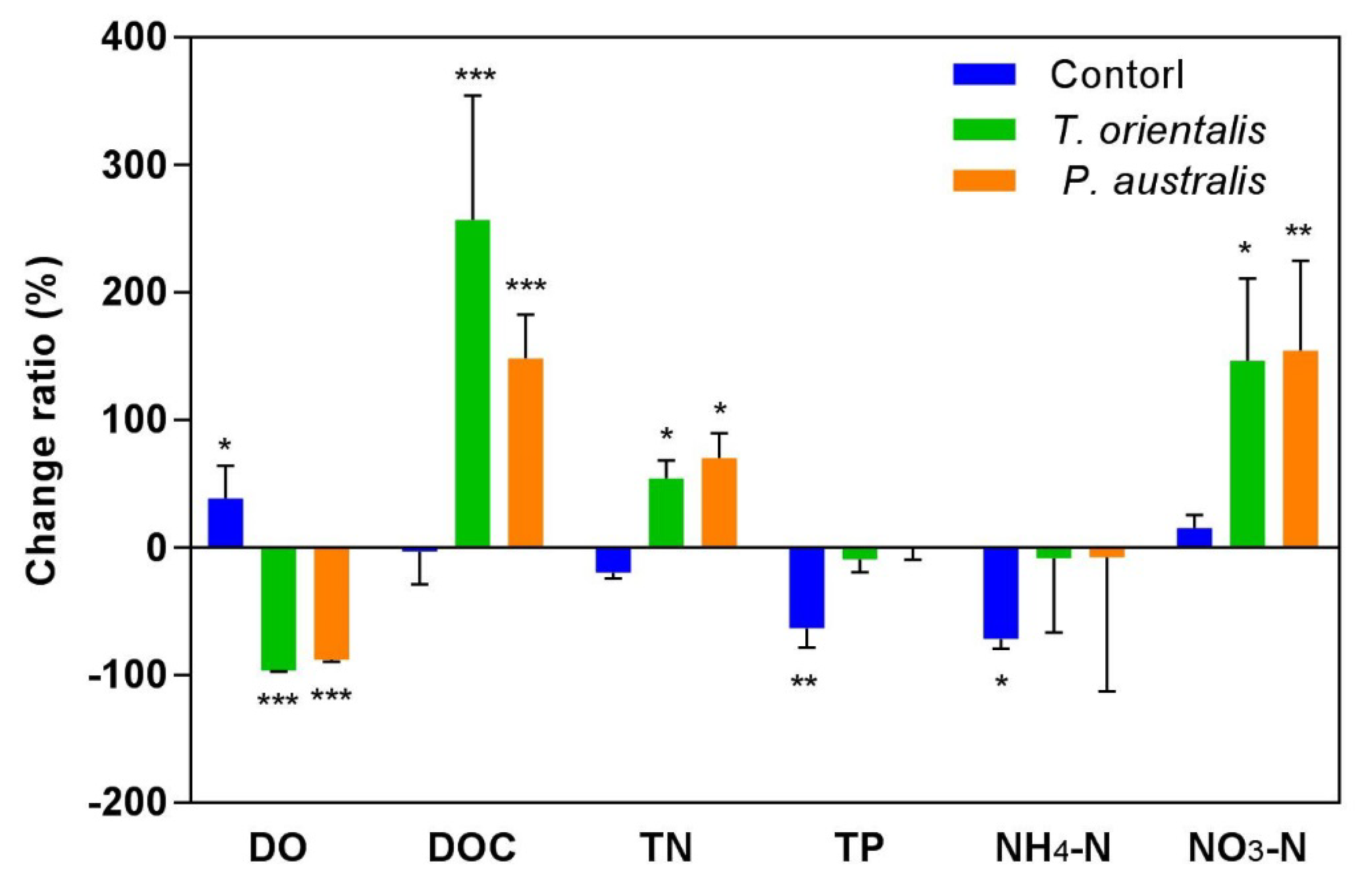
3.2. Dynamics of Water Quality
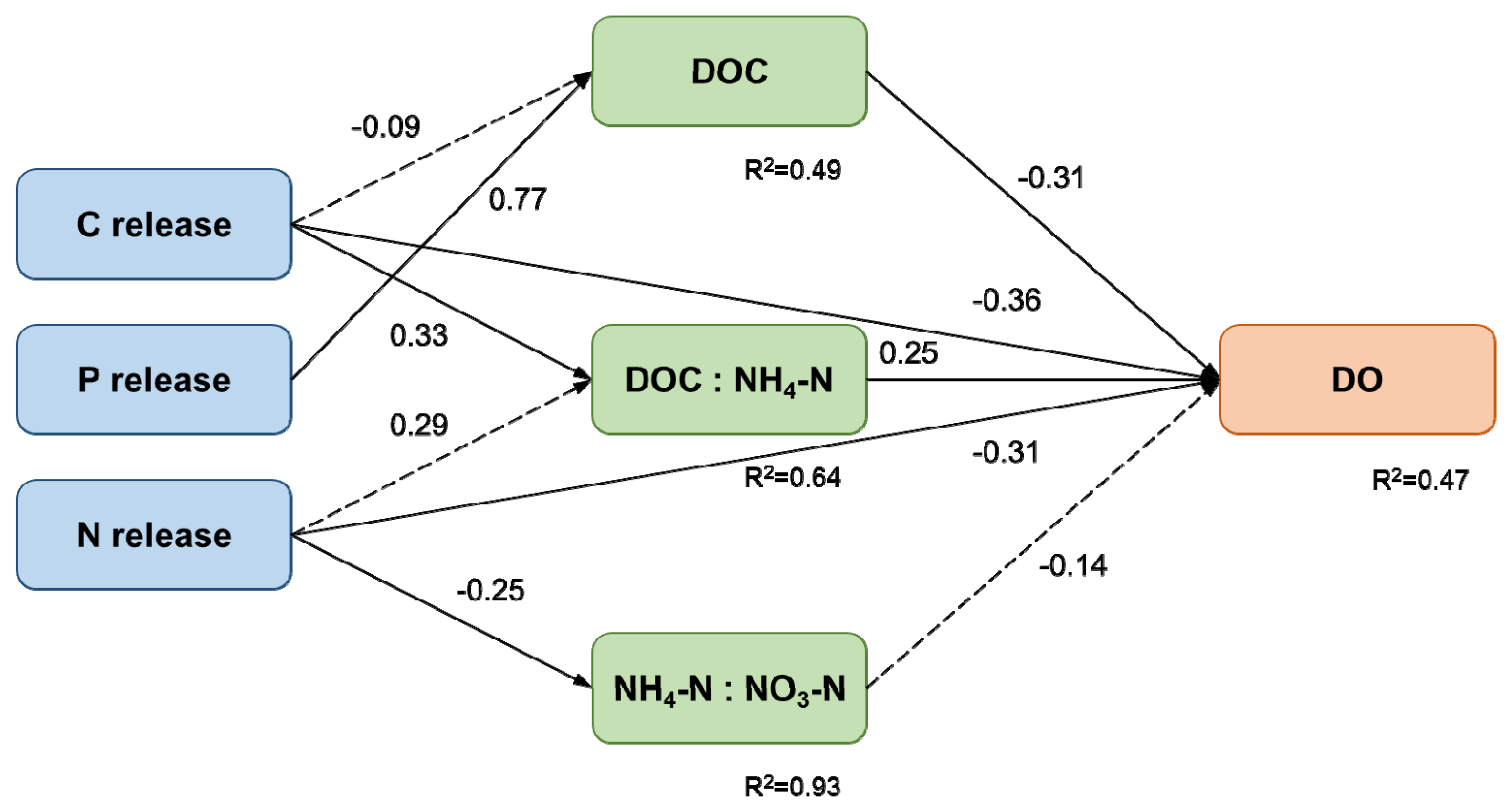
3.3. Relationship between Water Quality and Decomposition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Sand (%) | Silt (%) | Clay (%) | Total Organic Carbon (mg/g) | Available Nitrogen (mg/kg) | Available Phosphorus (mg/kg) | Available Potassium (mg/g) |
|---|---|---|---|---|---|---|---|
| 8.39 | 15.57 | 36.40 | 48.03 | 1.79 | 20.61 | 3.96 | 0.13 |

| C (mg/g) | N (mg/g) | P (mg/g) | |
|---|---|---|---|
| Typha orientalis | 412.56 ± 7.93 a | 13.77 ± 1.68 a | 1.35 ± 0.09 a |
| Phragmites australis | 395.31 ± 9.45 b | 9.95 ± 1.05 b | 0.49 ± 0.05 b |
References
- Asaeda, T.; Trung, V.K.; Manatunge, J. Modeling the effects of macrophyte growth and decomposition on the nutrient budget in Shallow Lakes. Aquat. Bot. 2000, 68, 217–237. [Google Scholar] [CrossRef]
- Lawniczak, A.E.; Zbierska, J.; Choiński, A.; Szczepaniak, W. Response of emergent macrophytes to hydrological changes in a shallow lake, with special reference to nutrient cycling. Hydrobiologia 2010, 656, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Christensen, J.R.; Crumpton, W.G.; van der Valk, A.G. Estimating the breakdown and accumulation of emergent macrophyte litter: A mass-balance approach. Wetlands 2009, 29, 204–214. [Google Scholar] [CrossRef]
- Banks, L.K.; Frost, P.C. Biomass loss and nutrient release from decomposing aquatic macrophytes: Effects of detrital mixing. Aquat. Sci. 2017, 79, 881–890. [Google Scholar] [CrossRef]
- Pieczyńska, E. Detritus and nutrient dynamics in the shore zone of lakes: A review. Hydrobiologia 1993, 251, 49–58. [Google Scholar] [CrossRef]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here. Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef]
- Sarneel, J.M.; Geurts, J.J.M.; Beltman, B.; Lamers, L.P.M.; Nijzink, M.M.; Soons, M.B. The effect of nutrient enrichment of either the bank or the surface water on shoreline vegetation and decomposition. Ecosystems 2010, 13, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Meerhoff, M.; Jacobsen, B.A.; Hansen, R.S.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.L.; Mazzeo, N.; Branco, C.W.C. Restoration of shallow lakes by nutrient control and biomanipulation-the successful strategy varies with lake size and climate. Hydrobiologia 2007, 581, 269–285. [Google Scholar] [CrossRef]
- Erik, J.; Martin, S.; Liu, Z. Lake restoration and management in a climate, change perspective: An introduction. Water 2017, 9, 122. [Google Scholar]
- Mann, C.J.; Wetzel, R.G. Effects of the emergent macrophyte Juncus Effusus L. on the chemical composition of interstitial water and bacterial productivity. Biogeochemistry 2000, 48, 307–322. [Google Scholar] [CrossRef]
- Pan, X.; Ping, Y.; Cui, L.; Li, W.; Zhang, X.; Zhou, J.; Yu, F.; Prinzing, A. Plant litter submergence affect the water quality of a constructed wetland. PLoS ONE 2017, 12, e0171019. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; He, S.; Huang, J.; Gu, J.; Zhou, W.; Gao, L. Decomposition of emergent aquatic plant (cattail) litter under different conditions and the influence on water quality. Water Air Soil Poll. 2017, 228, 70. [Google Scholar] [CrossRef]
- Morris, J.T.; Lajtha, K. Decomposition and nutrient dynamics of litter from four species of freshwater emergent macrophytes. Hydrobiologia 1986, 131, 215–223. [Google Scholar] [CrossRef]
- Belova, M. Microbial decomposition of freshwater macrophytes in the littoral zone of lakes. Hydrobiologia 1993, 251, 59–64. [Google Scholar] [CrossRef]
- Kröger, R.; Holland, M.M.; Moore, M.T.; Cooper, C.M. Plant senescence: A mechanism for nutrient release in temperate agricultural wetlands. Environ. Poll. 2007, 146, 114–119. [Google Scholar] [CrossRef]
- Menon, R.; Holland, M.M. Phosphorus release due to decomposition of wetland plants. Wetlands 2014, 34, 1191–1196. [Google Scholar] [CrossRef]
- Longhi, D.; Bartoli, M.; Viaroli, P. Decomposition of four macrophytes in wetland sediments: Organic matter and nutrient decay and associated benthic processes. Aquat. Bot. 2008, 89, 303–310. [Google Scholar] [CrossRef]
- Eriksson, P.G.; Andersson, J.L. Potential nitrification and cation exchange on litter of emergent, freshwater macrophytes. Freshw. Biol. 1999, 42, 479–486. [Google Scholar]
- Li, X.; Cui, B.; Yang, Q.; Lan, Y.; Wang, T.; Han, Z. Effects of plant species on macrophyte decomposition under three nutrient conditions in a eutrophic shallow lake, North China. Ecol. Model. 2013, 252, 121–128. [Google Scholar] [CrossRef]
- Gingerich, R.T.; Merovich, G.; Anderson, J.T. Influence of environmental parameters on litter decomposition in wetlands in West Virginia, USA. J. Freshwater Ecol. 2014, 29, 535–549. [Google Scholar] [CrossRef]
- Kirillin, G.; Lepparanta, M.; Terzhevik, A.; Nikolai, G.; Bernhardt, J.; Engelhardt, C.; Efremova, T.; Golosov, S.; Palshin, N.; Sherstyankin, P.; et al. Physics of seasonally ice-covered lakes: A review. Aquat. Sci. 2012, 74, 659–682. [Google Scholar] [CrossRef]
- Lindenschmidt, K.; Baulch, H.M.; Cavaliere, E. River and lake ice processes—Impacts of freshwater ice on aquatic ecosystems in a changing globe. Water 2018, 10, 1586. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Yu, Y.; Ma, M.; Guo, J.; Zhao, N.; Li, X. Effects of water replenishment from Yellow River on water quality of Hengshui Lake. J. Marine Biol. Aquacul. 2018, 4, 11–13. [Google Scholar]
- Kim, Y.; Ullah, S.; Moore, T.R.; Roulet, N.T. Dissolved organic carbon and total dissolved nitrogen production by boreal soils and litter: The role of flooding, oxygen concentration, and temperature. Biogeochemistry 2014, 118, 35–48. [Google Scholar] [CrossRef]
- Margesin, R.; Miteva, V. Diversity and ecology of psychrophilic microorganisms. Res. Microbiol. 2011, 162, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M.; Fujiu, S.; Xiao, N.; Hanada, Y.; Kudoh, S.; Kondo, H.; Tsuda, S.; Hoshino, T. Cold adaptation of fungi obtained from soil and lake sediment in the Skarvsnes ice-free area, Antarctica. FEMS Microbiol. Lett. 2013, 346, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Beall, B.F.N.; Twiss, M.R.; Smith, D.E.; Oyserman, B.O.; Mckay, R.M.L. Ice cover extent drives phytoplankton- and bacterial community structure in a large north-temperate lake: Implications for a warming climate. Environ. Microbiol. 2016, 18, 1704–1719. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; He, Y.; Jiang, H. Inferior adaptation of bay sediments in a eutrophic shallow lake to winter season for organic matter decomposition. Environ. Poll. 2016, 219, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yang, W.; Zhang, J.; Deng, R. Fine root decomposition in two subalpine forests during the freeze-thaw season. Can. J. For. Res. 2010, 40, 135–140. [Google Scholar] [CrossRef]
- Bohman, I.M.; Herrmann, J. The timing of winter-growing shredder species and leaf litter turnover rate in an oligotrophic lake, SE Sweden. Hydrobiologia 2006, 556, 99–108. [Google Scholar] [CrossRef]
- Wang, M.; Hao, T.; Deng, X.; Wang, Z.; Li, Z. Effects of sediment-borne nutrient and litter quality on macrophyte decomposition and nutrient release. Hydrobiologia 2017, 787, 205–215. [Google Scholar] [CrossRef]
- Xie, Y.; Xie, Y.; Xiao, H.; Chen, X.; Li, F. Controls on litter decomposition of emergent macrophyte in Dongting lake wetlands. Ecosystems 2017, 20, 1383–1389. [Google Scholar] [CrossRef]
- Villa, J.A.; Mitsch, W.J.; Song, K.; Miao, S. Contribution of different wetland plant species to the DOC exported from a mesocosm experiment in the Florida Everglades. Ecol. Eng. 2014, 71, 118–125. [Google Scholar] [CrossRef]
- Bonanomia, G.; Senatorea, M.; Migliozzia, A.; de Marcob, A.; Pintimallic, A.; Lanzottia, V.; Mazzoleniaa, S. Decomposition of submerged plant litter in a Mediterranean reservoir: A microcosm study. Aquat. Bot. 2015, 120, 169–177. [Google Scholar] [CrossRef]
- Assunção, A.W.d.A.; Souza, B.P.; da Cunha-Santino, M.B.; Bianchini, I., Jr. Formation and mineralization kinetics of dissolved humic substances from aquatic macrophytes decomposition. J. Soil Sediment. 2016, 18, 1252–1264. [Google Scholar]
- Whitworth, K.L.; Baldwin, D.S.; Kerr, J.L. The effect of temperature on leaching and subsequent decomposition of dissolved carbon from inundated floodplain litter: Implications for the generation of hypoxic blackwater in lowland floodplain rivers. Chem. Ecol. 2014, 30, 491–500. [Google Scholar] [CrossRef]
- Hume, N.P.; Fleming, M.S.; Horne, A.J. Denitrification potential and carbon quality of four aquatic plants in wetland microcosms. Soil Sci. Soc. Am. J. 2002, 66, 1706–1712. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Xu, J. Improving winter performance of constructed wetlands for wastewater treatment in Northern China: A review. Wetlands 2014, 34, 243–253. [Google Scholar] [CrossRef]
- Mueller, S.; Mitrovic, S.M.; & Baldwin, D.S. Oxygen and dissolved organic carbon control release of N, P and Fe from the sediments of a shallow, polymictic lake. J. Soil Sediment. 2016, 16, 1109–1120. [Google Scholar] [CrossRef]
- Meding, M.E.; Jackson, L.J. Biological implications of empirical models of winter oxygen depletion. Can. J. Fish. Aquat. Sci. 2001, 58, 1727–1736. [Google Scholar] [CrossRef]
- Powers, S.M.; Baulch, H.M.; Hampton, S.E.; Labou, S.G.; Lottig, N.R.; Stanley, E.H. Nitrification contributes to winter oxygen depletion in seasonally frozen forested lakes. Biogeochemistry 2017, 136, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Sanchez, A.; Gonzalez-Martinez, A.; Martinez-Toledo, M.V.; Garcia-Ruiz, M.J.; Osorio, F.; Gonzalez-Lopez, J. The effect of influent characteristics and operational conditions over the performance and microbial community structure of partial nitritation reactors. Water 2014, 6, 1905–1924. [Google Scholar] [CrossRef] [Green Version]
- Strauss, E.A.; Lamberti, G.A. Effect of dissolved organic carbon quality on microbial decomposition and nitrification rates in stream sediments. Freshw. Biol. 2002, 47, 65–74. [Google Scholar] [CrossRef]






| Source | DO | DOC | TN | TP | NH4-N | NO3-N |
|---|---|---|---|---|---|---|
| Treatment (T) | F2,6 = 2702.39 ** | F2,6 = 471.37 ** | F2,6 = 24.26 ** | F2,6 = 244.92 ** | F2,6 = 15.47 ** | F2,6 = 16.07 ** |
| Sampling time (S) | F9,54 = 125.36 ** | F9,54 = 11.72 ** | F9,54 = 6.63 ** | F9,54 = 4.71 ** | F9,54 = 10.25 ** | F9,54 = 3.02 ** |
| T × S | F18,54 = 37.29 ** | F18,54 = 5.22 ** | F18,54 = 3.72** | F18,54 = 4.11 ** | F18,54 = 1.74 ns | F18,54 = 0.88 ns |
| DO | DOC | TN | TP | NH4-N | NO3-N | |
|---|---|---|---|---|---|---|
| T. orientalis | ||||||
| C release | −0.605 ns | 0.453 ns | −0.409 ns | −0.045 ns | −0.528 ns | 0.277 ns |
| N release | −0.656 * | 0.447 ns | −0.441 ns | 0.003 ns | −0.523 ns | 0.183 ns |
| P release | −0.788 * | 0.599 ns | −0.238 ns | −0.112 ns | −0.320 ns | 0.306 ns |
| P. australis | ||||||
| C release | −0.717 * | 0.954 ** | 0.458 ns | −0.119 ns | −0.313 ns | 0.683 * |
| N release | −0.696 * | 0.923 ** | 0.718 * | −0.022 ns | −0.015 ns | 0.750 * |
| P release | −0.785 ** | 0.842 * | 0.458 ns | 0.071 ns | −0.139 ns | 0.547 ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Zhang, M.; Cui, L.; Pan, X.; Liu, W.; Li, W.; Lei, Y. Winter Decomposition of Emergent Macrophytes Affects Water Quality under Ice in a Temperate Shallow Lake. Water 2020, 12, 2640. https://doi.org/10.3390/w12092640
Wei Y, Zhang M, Cui L, Pan X, Liu W, Li W, Lei Y. Winter Decomposition of Emergent Macrophytes Affects Water Quality under Ice in a Temperate Shallow Lake. Water. 2020; 12(9):2640. https://doi.org/10.3390/w12092640
Chicago/Turabian StyleWei, Yuanyun, Manyin Zhang, Lijuan Cui, Xu Pan, Weiwei Liu, Wei Li, and Yinru Lei. 2020. "Winter Decomposition of Emergent Macrophytes Affects Water Quality under Ice in a Temperate Shallow Lake" Water 12, no. 9: 2640. https://doi.org/10.3390/w12092640