Characteristics of Ammonia Removal and Nitrifying Microbial Communities in a Hybrid Biofloc-RAS for Intensive Litopenaeus vannamei Culture: A Pilot-Scale Study

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
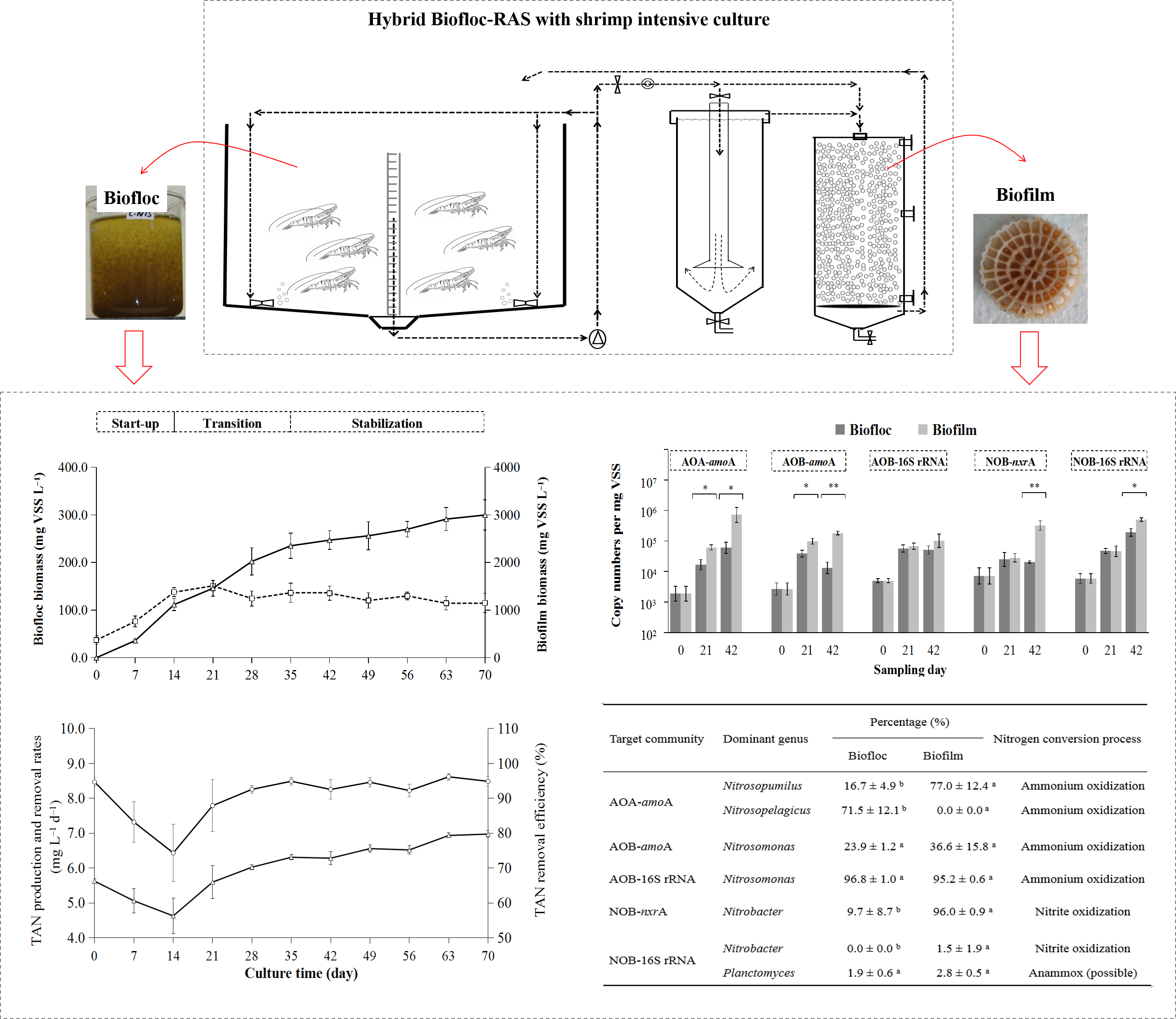
2.1. Design and Setup of a Hybrid RAS
2.2. Shrimp Production Trial and System Operation Condition
2.3. Determination of Nitrogen Dynamics, TAN Removal, and Shrimp Performance
2.4. A Sampling of Biofloc and Biofilm and Determination of Their Biomass
2.5. Microbial DNA Extraction and qPCR Analysis
2.6. Miseq Sequencing and Bioinformatics Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Operating Mode, Nitrogen Dynamics, and Nitrification Establishment
3.2. Comparison of Nitrifying Microbial Communities Between Biofloc and Biofilm
3.3. Correlation between TAN Removal and Nitrifying Functional Communities of Biofloc and Biofilm
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO (Food and Agriculture Organization of the United Nations). Global Aquaculture Production 1950–2018 (Online Query); FAO Fisheries and Aquaculture Department: Rome, Italy, 2020; Available online: http://www.fao.org/fishery/topic/16140/en (accessed on 18 September 2020).
- Belton, B.; Reardon, T.; Zilberman, D. Sustainable commoditization of seafood. Nat. Sustain. 2020, 3, 677–684. [Google Scholar] [CrossRef]
- Nguyen, T.A.T.; Nguyen, K.A.T.; Jolly, C. Is Super-Intensification the Solution to Shrimp Production and Export Sustainability? Sustainability 2019, 11, 5277. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, J.S.; Hamilton II, R.D.; Sandier, P.A.; Browdy, C.L.; Stokes, A.D. Effect of water exchange rate on production, water quality, effluent characteristics and nitrogen budgets of intensive shrimp ponds. J. World Aquac. Soc. 1993, 24, 304–320. [Google Scholar] [CrossRef]
- Han, S.; Wang, B.; Wang, M.; Liu, Q.; Zhao, W.; Wang, L. Effects of ammonia and nitrite accumulation on the survival and growth performance of white shrimp Litopenaeus vannamei. Invert. Surviv. J. 2017, 14, 221–232. [Google Scholar] [CrossRef]
- Hopkins, J.S.; DeVoe, M.R.; Holland, A.F. Environmental impacts of shrimp farming with special reference to the situation in the continental United States. Estuaries 1995, 18, 25–42. [Google Scholar] [CrossRef]
- Chen, Z.; Ge, H.X.; Chang, Z.Q.; Song, X.F.; Zhao, F.Z.; Li, J. Nitrogen Budget in Recirculating Aquaculture and Water Exchange Systems for Culturing Litopenaeus vannamei. J. Ocean Univ. China 2018, 17, 905–912. [Google Scholar] [CrossRef]
- Hamilton, S. Assessing the Role of Commercial Aquaculture in Displacing Mangrove Forest. Bull. Mar. Sci. 2013, 89, 585–601. [Google Scholar] [CrossRef]
- Hatje, V.; de Souza, M.M.; Ribeiro, L.F.; Eca, G.F.; Barros, F. Detection of environmental impacts of shrimp farming through multiple lines of evidence. Environ. Pollut. 2016, 219, 672–684. [Google Scholar] [CrossRef]
- Páez-Osuna, F. The environmental impact of shrimp aquaculture: A global perspective. Environ. Pollut. 2001, 112, 229–231. [Google Scholar] [CrossRef]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Joffre, O.M.; Klerkx, L.; Khoa, T.N.D. Aquaculture innovation system analysis of transition to sustainable intensification in shrimp farming. Agron. Sustain. Dev. 2018, 38, 34. [Google Scholar] [CrossRef] [Green Version]
- Klinger, D.; Naylor, R. Searching for Solutions in Aquaculture: Charting a Sustainable Course. Annu. Rev. Environ. Resour. 2012, 37, 247–276. [Google Scholar] [CrossRef] [Green Version]
- Robles-Porchas, G.R.; Gollas-Galván, T.; Martínez-Porchas, M.; Martínez-Cordova, L.R.; Miranda-Baeza, A.; Vargas-Albores, F. The nitrification process for nitrogen removal in biofloc system aquaculture. Rev. Aquacult. 2020, 1–22. [Google Scholar] [CrossRef]
- Rurangwa, E.; Verdegem, M.C.J. Microorganisms in recirculating aquaculture systems and their management. Rev. Aquacult. 2015, 7, 117–130. [Google Scholar] [CrossRef]
- Avnimelech, Y. Bio-filters: The need for a new comprehensive approach. Aquac. Eng. 2006, 34, 172–178. [Google Scholar] [CrossRef]
- Schreier, H.J.; Mirzoyan, N.; Saito, K. Microbial diversity of biological filters in recirculating aquaculture systems. Curr. Opin. Biotech. 2010, 21, 318–325. [Google Scholar] [CrossRef]
- Brown, M.N.; Briones, A.; Diana, J.; Raskin, L. Ammonia-oxidizing archaea and nitrite-oxidizing nitrospiras in the biofilter of a shrimp recirculating aquaculture system. FEMS Microbiol. Ecol. 2013, 83, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Holl, C.M.; Glazer, C.T.; Moss, S.M. Nitrogen stable isotopes in recirculating aquaculture for super-intensive shrimp production: Tracing the effects of water filtration on microbial nitrogen cycling. Aquaculture 2011, 311, 146–154. [Google Scholar] [CrossRef]
- Kumar, V.J.R.; Joseph, V.; Vijai, R.; Philip, R.; Singh, I.S.B. Nitrification in a packed bed bioreactor integrated into a marine recirculating maturation system under different substrate concentrations and flow rates. J. Chem. Technol. Biot. 2011, 86, 790–797. [Google Scholar] [CrossRef]
- Suantika, G.; Situmorang, M.L.; Kurniawan, J.B.; Pratiwi, S.A.; Aditiawati, P.; Astuti, D.I.; Azizah, F.F.N.; Djohan, Y.A.; Zuhri, U.; Simatupang, T.M. Development of a zero water discharge (ZWD)-Recirculating aquaculture system (RAS) hybrid system for super intensive white shrimp (Litopenaeus vannamei) culture under low salinity conditions and its industrial trial in commercial shrimp urban farming in Gresik, East Java, Indonesia. Aquac. Eng. 2018, 82, 12–24. [Google Scholar] [CrossRef]
- Kaya, D.; Genc, E.; Genc, M.A.; Aktas, M.; Eroldogan, O.T.; Guroy, D. Biofloc technology in recirculating aquaculture system as a culture model for green tiger shrimp, Penaeus semisulcatus: Effects of different feeding rates and stocking densities. Aquaculture 2020, 528, 735526. [Google Scholar] [CrossRef]
- Gutierrez-Wing, M.T.; Malone, R.F. Biological filters in aquaculture: Trends and research directions for freshwater and marine applications. Aquac. Eng. 2006, 34, 163–171. [Google Scholar] [CrossRef]
- Ruiz, P.; Vidal, J.M.; Sepulveda, D.; Torres, C.; Villouta, G.; Carrasco, C.; Aguilera, F.; Ruiz-Tagle, N.; Urrutia, H. Overview and future perspectives of nitrifying bacteria on biofilters for recirculating aquaculture systems. Rev. Aquacult. 2019, 12, 1478–1494. [Google Scholar] [CrossRef]
- Avnimelech, Y. Biofloc Technology—A Practical Guide Book, 3rd ed.; The World Aquaculture Society: Baton Rouge, LA, USA, 2015. [Google Scholar]
- Samocha, T.M.; Prangnell, D.I.; Hanson, T.R.; Treece, G.D.; Morris, T.C.; Castro, L.F.; Staresinic, N. Design and Operation of Super Intensive, Biofloc-Dominated Systems for Indoor Production of the Pacific White Shrimp, Litopenaeus vannamei—The Texas A&M AgriLife Research Experience; The World Aquaculture Society: Baton Rouge, LA, USA, 2017. [Google Scholar]
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia-nitrogen in aquaculture systems. Aquaculture 2006, 257, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, J.A. Biofloc Production Systems for Aquaculture; No. 4503; Southern Regional Aquaculture Center: Stoneville, MS, USA, 2013; pp. 1–12. [Google Scholar]
- Ray, A.J.; Lewis, B.L.; Browdy, C.L.; Leffler, J.W. Suspended solids removal to improve shrimp (Litopenaeus vannamei) production and an evaluation of a plant-based feed in minimal-exchange, superintensive culture systems. Aquaculture 2010, 299, 89–98. [Google Scholar] [CrossRef]
- Schveitzer, R.; Arantes, R.; Costodio, P.F.S.; Santo, C.M.D.; Arana, L.V.; Seiffert, W.Q.; Andreatta, E.R. Effect of different biofloc levels on microbial activity, water quality and performance of Litopenaeus vannamei in a tank system operated with no water exchange. Aquac. Eng. 2013, 56, 59–70. [Google Scholar] [CrossRef]
- Ray, A.J.; Drury, T.H.; Cecil, A. Comparing clear-water RAS and biofloc systems: Shrimp (Litopenaeus vannamei) production, water quality, and biofloc nutritional contributions estimated using stable isotopes. Aquac. Eng. 2017, 77, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Tierney, T.W.; Ray, A.J. Comparing biofloc, clear-water, and hybrid nursery systems (Part I): Shrimp (Litopenaeus vannamei) production, water quality, and stable isotope dynamics. Aquac. Eng. 2018, 82, 73–79. [Google Scholar] [CrossRef]
- Xu, W.; Xu, Y.; Su, H.; Hu, X.; Xu, Y.; Li, Z.; Wen, G.; Cao, Y. Production performance, inorganic nitrogen control and bacterial community characteristics in a controlled biofloc-based system for indoor and outdoor super-intensive culture of Litopenaeus Vannamei. Aquaculture 2021, 531, 735749. [Google Scholar] [CrossRef]
- APHA (American Public Health Association). Standard Methods for the Examination of the Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [Green Version]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [Green Version]
- Kowalchuk, G.A.; Stephen, J.R.; De Boer, W.; Prosser, J.I.; Embley, T.M.; Woldendorp, J.W. Analysis of ammonia-oxidizing bacteria of the beta subdivision of the class proteobacteria in coastal sand dunes by denaturing gradient gel electrophoresis and sequencing of PCR-amplified 16s ribosomal DNA fragments. Appl. Environ. Microbiol. 1997, 63, 1489–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poly, F.; Wertz, S.; Brothier, E.; Degrange, V. First exploration of Nitrobacter diversity in soils using functional gene nxrA encoding nitrite oxido-reductase. FEMS Microbiol. Ecol. 2008, 63, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Degrange, V.; Bardin, R. Detection and counting of Nitrobacter populations in soil by PCR. Appl. Environ. Microbiol. 1995, 61, 2093–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Xu, W.J.; Morris, T.C.; Samocha, T.M. Effects of C/N ratio on biofloc development, water quality, and performance of Litopenaeus vannamei juveniles in a biofloc-based, high-density, zero-exchange, outdoor tank system. Aquaculture 2016, 453, 169–175. [Google Scholar] [CrossRef]
- Bassin, J.P.; Kleerebezem, R.; Rosado, A.S.; van Loosdrecht, M.C.M.; Dezotti, M. Effect of Different Operational Conditions on Biofilm Development, Nitrification, and Nitrifying Microbial Population in Moving-Bed Biofilm Reactors. Environ. Sci. Technol. 2012, 46, 1546–1555. [Google Scholar] [CrossRef]
- Kumar, V.J.R.; Achuthan, C.; Manju, N.J.; Philip, R.; Singh, I.S. Mass production of nitrifying bacterial consortia for the rapid establishment of nitrification in saline recirculating aquaculture systems. World J. Microb. Biot. 2009, 25, 407–414. [Google Scholar] [CrossRef]
- Shao, Y.X.; Shi, Y.J.; Mohammed, A.; Liu, Y. Wastewater ammonia removal using an integrated fixed-film activated sludge-sequencing batch biofilm reactor (IFAS-SBR): Comparison of suspended flocs and attached biofilm. Int. Biodeter. Biodegr. 2017, 116, 38–47. [Google Scholar] [CrossRef]
- Blancheton, J.P.; Attramadal, K.J.K.; Michaud, L.; d’Orbcastel, E.R.; Vadstein, O. Insight into bacterial population in aquaculture systems and its implication. Aquac. Eng. 2013, 53, 30–39. [Google Scholar] [CrossRef]
- Wittebolle, L.; Verstraete, W.; Boon, N. The inoculum effect on the ammonia-oxidizing bacterial communities in parallel sequential batch reactors. Water Res. 2009, 43, 4149–4158. [Google Scholar] [CrossRef]
- Sudarno, U.; Bathe, S.; Winter, J.; Gallert, C. Nitrification in fixed-bed reactors treating saline wastewater. Appl. Microbiol. Biot. 2010, 85, 2017–2030. [Google Scholar] [CrossRef] [Green Version]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Carini, P.; Dupont, C.L.; Santoro, A.E. Patterns of thaumarchaeal gene expression in culture and diverse marine environments. Environ. Microbiol. 2018, 20, 2112–2124. [Google Scholar] [CrossRef]
- Santoro, A.E.; Dupont, C.L.; Richter, R.A.; Craig, M.T.; Carini, P.; McIlvin, M.R.; Yang, Y.; Orsi, W.D.; Moran, D.M.; Saito, M.A. Genomic and proteomic characterization of “Candidatus Nitrosopelagicus brevis”: An ammonia-oxidizing archaeon from the open ocean. Proc. Natl. Acad. Sci. USA 2015, 112, 1173–1178. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.B.; de la Torre, J.R.; Klotz, M.G.; Urakawa, H.; Pinel, N.; Arp, D.J.; Brochier-Armanet, C.; Chain, P.S.G.; Chan, P.P.; Gollabgir, A.; et al. Nitrosopumilus maritimus genome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine crenarchaea. Proc. Natl. Acad. Sci. USA 2010, 107, 8818–8823. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.T.; Wan, R.; Song, X.F.; Liu, Y.; Hallerman, E.; Dong, D.P.; Zhai, J.M.; Zhang, H.S.; Sun, L.Y. Metagenomic analysis shows diverse, distinct bacterial communities in biofilters among different marine recirculating aquaculture systems. Aquacult. Int. 2016, 24, 1393–1408. [Google Scholar] [CrossRef]
- Sakami, T.; Andoh, T.; Morita, T.; Yamamoto, Y. Phylogenetic diversity of ammonia-oxidizing archaea and bacteria in biofifilters of recirculating aquaculture systems. Mar. Genom. 2012, 7, 27–31. [Google Scholar] [CrossRef]
- Foesel, B.U.; Gieseke, A.; Schwermer, C.; Stief, P.; Koch, L.; Cytryn, E.; de la Torré, J.R.; van Rijn, J.; Minz, D.; Drake, H.L.; et al. Nitrosomonas Nm143-like ammonia oxidizers and Nitrospira marina-like nitrite oxidizers dominate the nitrifier community in a marine aquaculture biofilm. FEMS Microbiol. Ecol. 2008, 63, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Chen, J.Y.; Gou, J.W.; Hou, J.; Li, D.P.; He, X.G. The effect of different carbon sources on water quality, microbial community and structure of biofloc systems. Aquaculture 2018, 482, 103–110. [Google Scholar] [CrossRef]
- Keuter, S.; Kruse, M.; Lipski, A.; Spieck, E. Relevance of Nitrospira for nitrite oxidation in a marine recirculation aquaculture system and physiological features of a Nitrospira marina-like isolate. Environ. Microbiol. 2011, 13, 2536–2547. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Lee, J.; Kim, Y.M.; Myeong, J.I.; Kim, K.H. Uncultured bacterial diversity in a seawater recirculating aquaculture system revealed by 16S rRNA gene amplicon sequencing. J. Microbiol. 2016, 54, 296–304. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name and Sequence | Target Gene | qPCR Amplification Procedure | Reference |
|---|---|---|---|
| Arch-amoAF 5′-STAATGGTCTGGCTTAGACG-3′ Arch-amoAR 5′-GCGGCCATCCATCTGTATGT-3′ | AOA-amoA | 95 °C for 30 s; 40 cycles (95 °C for 10 s, 50 °C for 30 s), and 72 °C for 45 s | [35] |
| amoA-1F 5′-GGGGTTTCTACTGGTGGT-3′ amoA-2R 5′-CCCCTCKGSAAAGCCTTCTTC-3′ | AOB-amoA | 95 °C for 30 s; 40 cycles (95 °C for 10 s, 55 °C for 30 s), and 72 °C for 30 s | [36] |
| CTO-189F 5′-GGAGRAAAGYAGGGGATCG-3′ CTO-654R 5′-CTAGCYTTGTAGTTTCAAACGC-3′ | AOB-16S rRNA | 95 °C for 30 s, 40 cycles (95 °C for 10 s, 55 °C for 30 s), and 72 °C for 30 s | [37] |
| nxrAF 5′-CAGACCGACGTGTGCGAAAG-3′ nxrAR 5′-TCYACAAGGAACGGAAGGTC-3′ | NOB-nxrA | 95 °C for 30 s, 40 cycles (95 °C for 10 s, 55 °C for 30 s), and 72 °C for 30 s | [38] |
| FGPS-872 5′-TTTTTTGAGATTTGCTAG-3′ FGPS-1269 5′-CTAAAACTCAAAGGAATTGA-3′ | NOB-16S rRNA | 95 °C for 30 s, 40 cycles (95 °C for 10 s, 50 °C for 30 s), and 72 °C for 30 s | [39] |
| Parameter | Mean ± S.D. | Min‒Max |
|---|---|---|
| Light intensity (lx) | 628 ± 11 | 475–942 |
| Salinity (g L−1) | 27.2 ± 0.1 | 25.9–28.5 |
| Water temperature (°C) | 28.9 ± 0.1 | 27.7–30.2 |
| Dissolved oxygen (mg L−1) | 5.2 ± 0 | 4.6–6.2 |
| pH | 7.3 ± 0 | 7.0–7.8 |
| Alkalinity (mg·L−1 as CaCO3) | 232 ± 9 | 155–330.0 |
| TAN (mg L−1) | 0.6 ± 0.1 | 0.21–1.81 |
| NO2−-N (mg L−1) | 0.6 ± 0.1 | 0.16–2.41 |
| NO3−-N (mg L−1) | 76.5 ± 2.7 | 2.7–146.3 |
| TN (mg L−1) | 89.9 ± 0.8 | 5.7–154.3 |
| VSS (mg L−1) | 116.1 ± 3.6 | 30.2–170.9 |
| Initial Weight (g) | Final Weight (g) | Growth Rate (g wk‒1) | Survival Rate (%) | Yield (kg m‒3) | Feed Conversion Rate |
|---|---|---|---|---|---|
| 1.92 ± 0.43 | 17.85 ± 0.56 | 2.15 ± 0.18 | 93.11 ± 1.66 | 4.98 ± 0.10 | 1.60 ± 0.03 |
| Target Community | Dominant Genus | Percentage (%) | Nitrogen Conversion Process | |
|---|---|---|---|---|
| Biofloc | Biofilm | |||
| AOA-amoA | Nitrosopumilus | 16.7 ± 4.9 b | 77.0 ± 12.4 a | Ammonium oxidization |
| Nitrosopelagicus | 71.5 ± 12.1 b | 0 ± 0 a | Ammonium oxidization | |
| AOB-amoA | Nitrosomonas | 23.9 ± 1.2 a | 36.6 ± 15.8 a | Ammonium oxidization |
| AOB-16S rRNA | Nitrosomonas | 96.8 ± 1.0 a | 95.2 ± 0.6 a | Ammonium oxidization |
| NOB-nxrA | Nitrobacter | 9.7 ± 8.7 b | 96.0 ± 0.9 a | Nitrite oxidization |
| NOB-16S rRNA | Nitrobacter | 0 ± 0 b | 1.5 ± 1.9 a | Nitrite oxidization |
| Planctomyces | 1.9 ± 0.6 a | 2.8 ± 0.5 a | Anammox (possible) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Xu, Y.; Su, H.; Hu, X.; Yang, K.; Wen, G.; Cao, Y. Characteristics of Ammonia Removal and Nitrifying Microbial Communities in a Hybrid Biofloc-RAS for Intensive Litopenaeus vannamei Culture: A Pilot-Scale Study. Water 2020, 12, 3000. https://doi.org/10.3390/w12113000
Xu W, Xu Y, Su H, Hu X, Yang K, Wen G, Cao Y. Characteristics of Ammonia Removal and Nitrifying Microbial Communities in a Hybrid Biofloc-RAS for Intensive Litopenaeus vannamei Culture: A Pilot-Scale Study. Water. 2020; 12(11):3000. https://doi.org/10.3390/w12113000
Chicago/Turabian StyleXu, Wujie, Yu Xu, Haochang Su, Xiaojuan Hu, Keng Yang, Guoliang Wen, and Yucheng Cao. 2020. "Characteristics of Ammonia Removal and Nitrifying Microbial Communities in a Hybrid Biofloc-RAS for Intensive Litopenaeus vannamei Culture: A Pilot-Scale Study" Water 12, no. 11: 3000. https://doi.org/10.3390/w12113000