How Did the Late 1980s Climate Regime Shift Affect Temperature-Sensitive Fish Population Dynamics: Case Study of Vendace (Coregonus albula) in a Large North-Temperate Lake


Abstract
:1. Introduction
2. Material and Methods
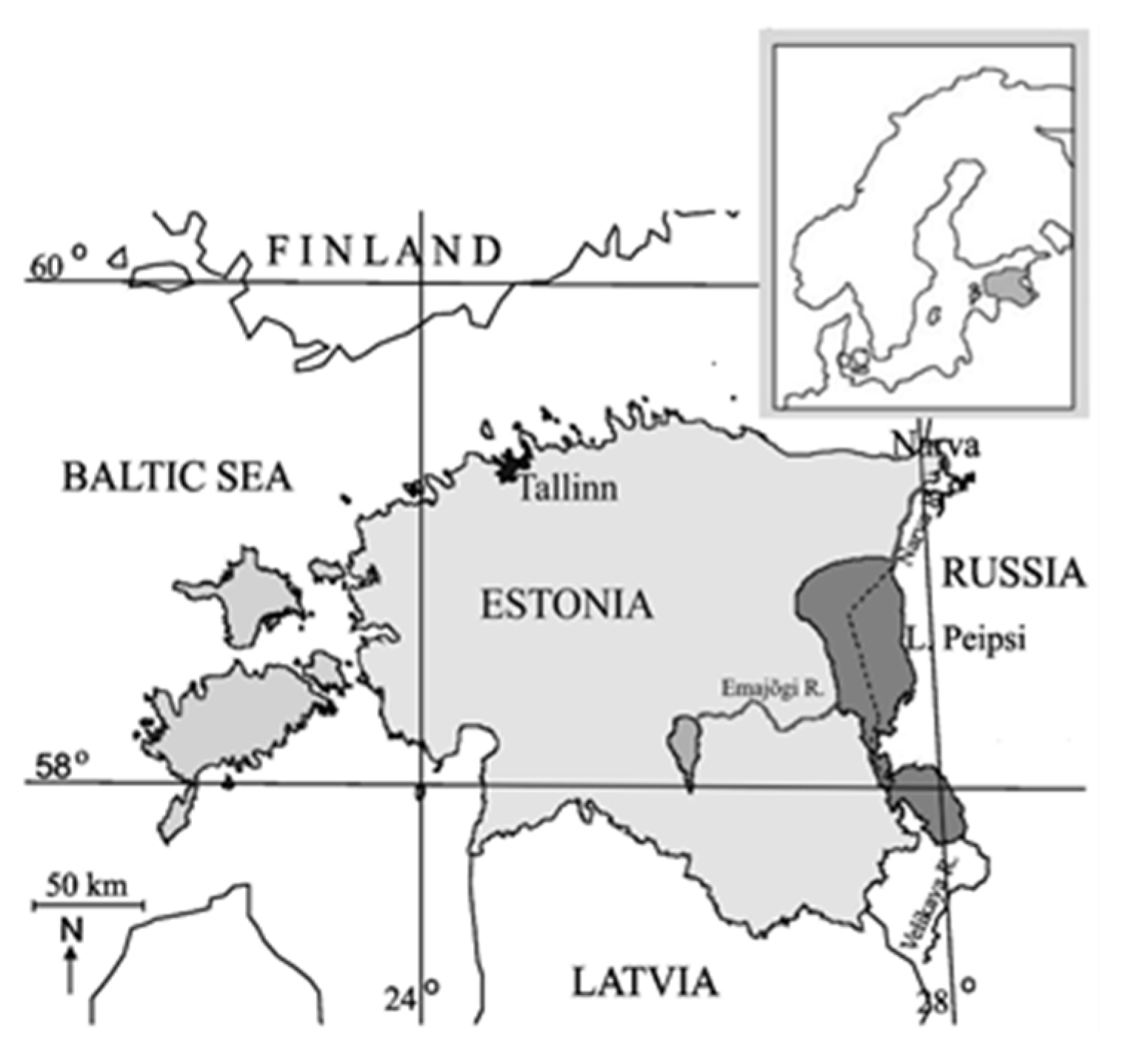
2.1. Study Area
2.2. Sampling and Dataset
2.3. Data Analysis
3. Results
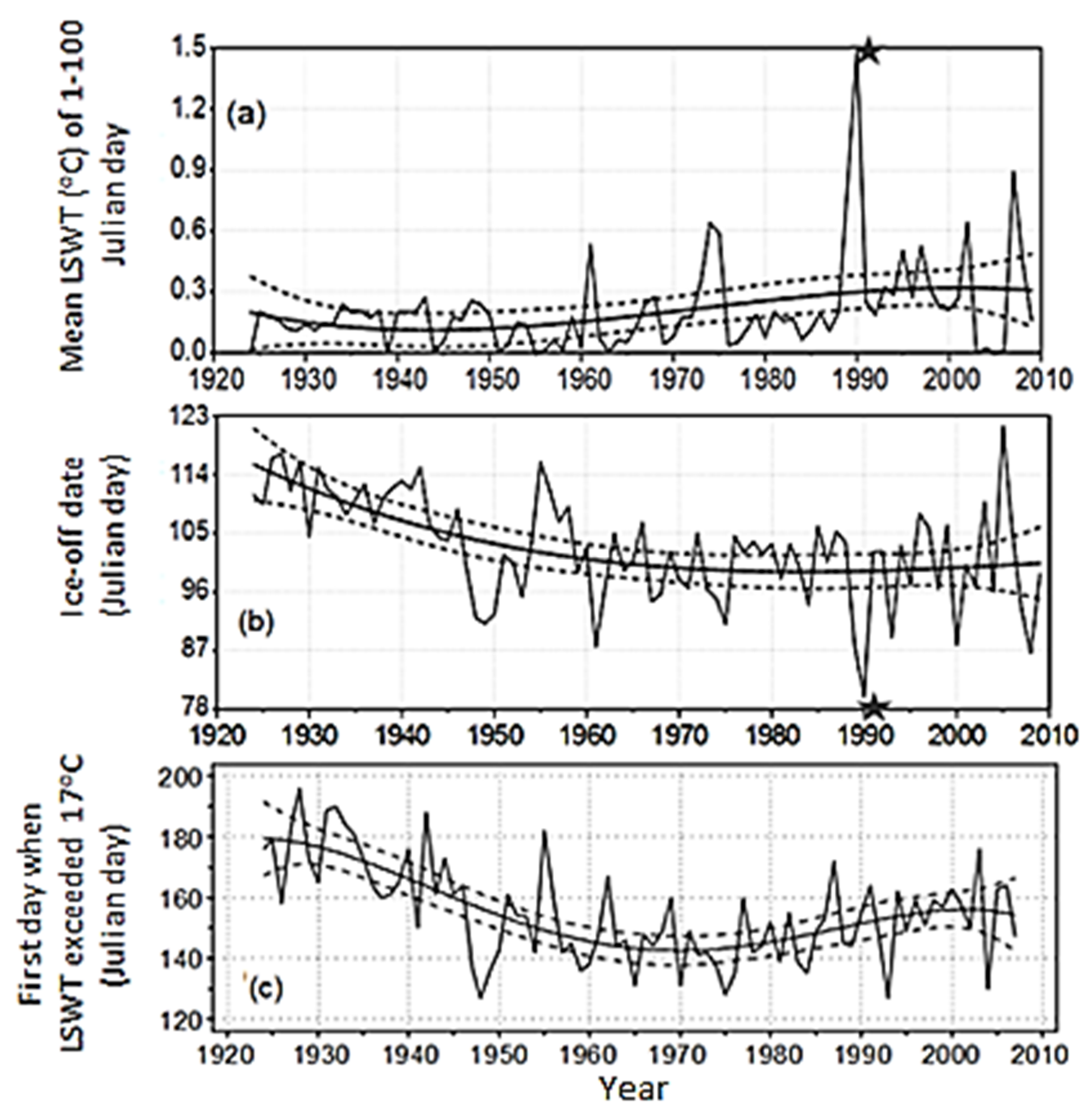
3.1. Variation of Ice Phenology and Lake Surface Water Temperature
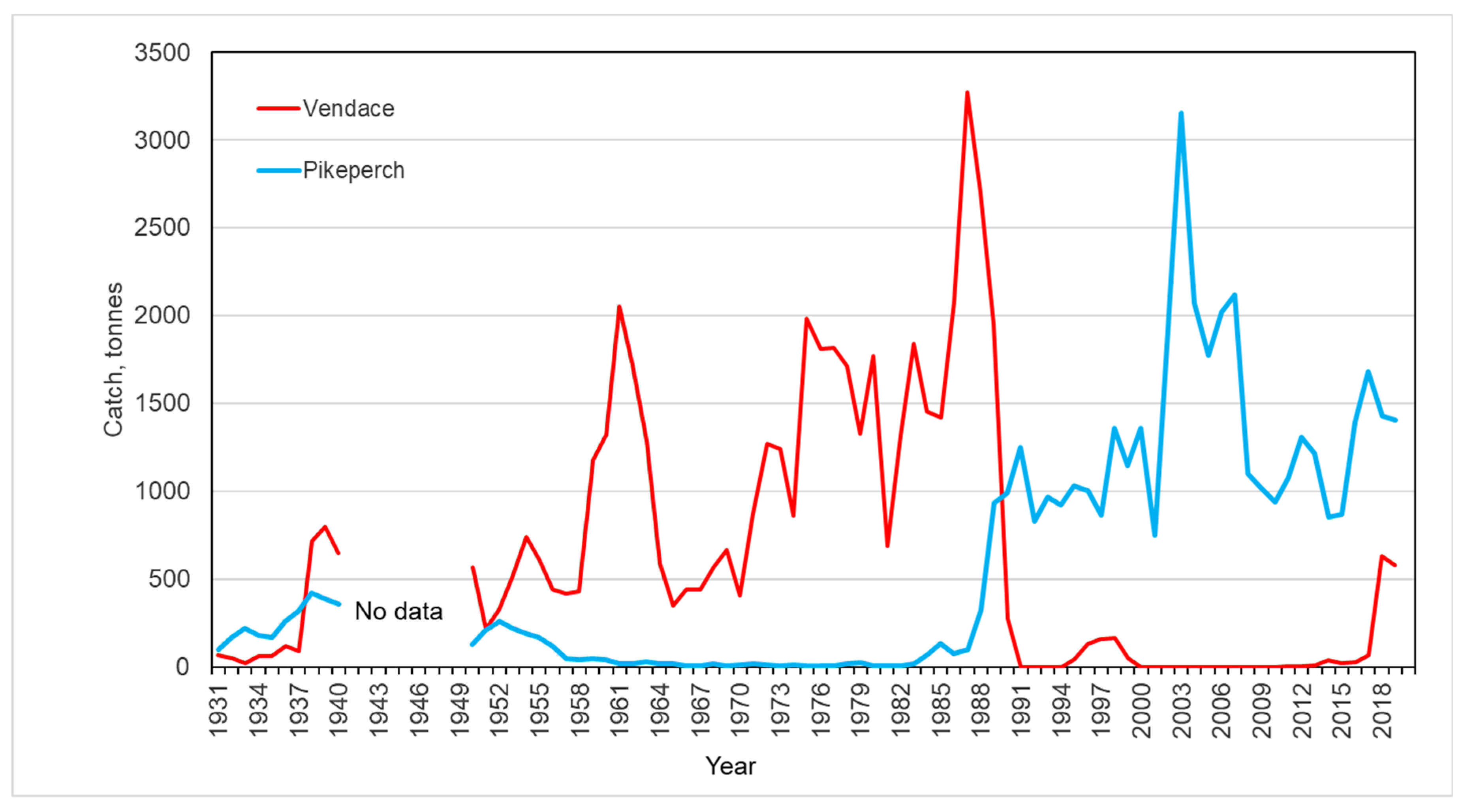
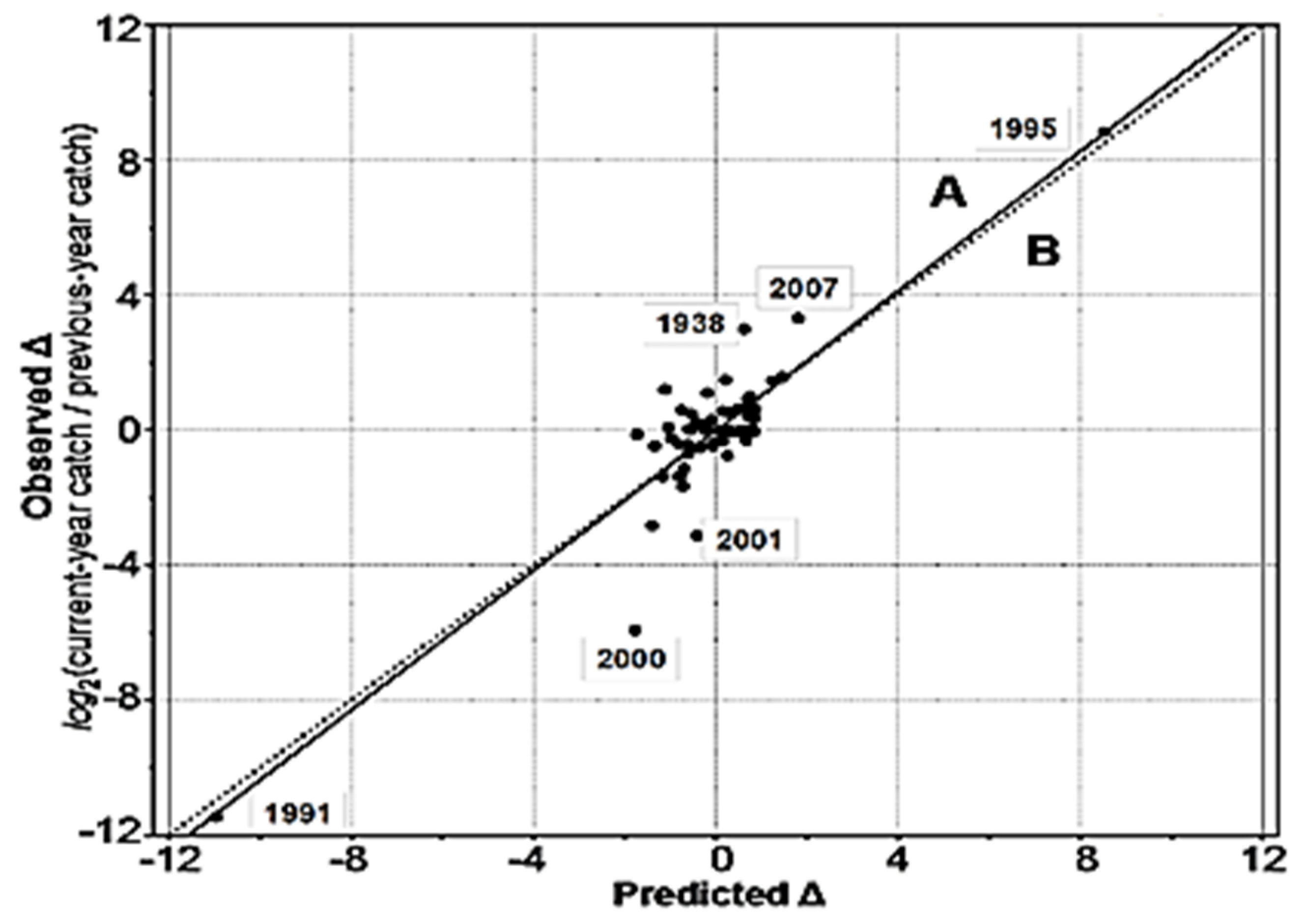
3.2. Trends in Fish Catch and Variability of the Vendace Stock
3.3. Water Temperature Parameters Related to Changes in Vendace Catch
3.4. Correlations between Fish Species in Catches
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allan, J.D.; Abell, R.; Hogan, Z.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K. Overfishing of Inland Waters. Biosci. 2005, 55, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Hunt, T.L.; Douglas, J.W.; Allen, M.S.; Gwinn, D.C.; Tonkin, Z.; Lyon, J.; Pickworth, A. Evaluation of population decline and fishing sustainability of the endangered Australian freshwater fish Macquaria australasica. Fish. Manag. Ecol. 2011, 18, 513–520. [Google Scholar] [CrossRef]
- Pukk, L.; Kuparinen, A.; Järv, L.; Gross, R.; Vasemägi, A. Genetic and life-history changes associated with fisheries-induced population collapse. Evol. Appl. 2013, 6, 749–760. [Google Scholar] [CrossRef]
- Cowx, I.G. Characterisation of inland fisheries in Europe. Fish. Manag. Ecol. 2015, 22, 78–87. [Google Scholar] [CrossRef]
- Kidd, K.A.; Blanchfield, P.J.; Mills, K.H.G.; Palace, V.P.; Evans, R.E.; Lazorchak, J.M.; Flick, R.W. Collapse of a fish population after exposure to a synthetic estrogen. Proc. Natl. Acad. Sci. 2007, 104, 8897–8901. [Google Scholar] [CrossRef] [Green Version]
- E Werner, E.; Gilliam, J.F. The Ontogenetic Niche and Species Interactions in Size-Structured Populations. Annu. Rev. Ecol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- Ohlberger, J.; Langangen, Ø.; Edeline, E.; Claessen, D.; Winfield, I.J.; Stenseth, N.C.; Vøllestad, L.A. Stage-specific biomass overcompensation by juveniles in response to increased adult mortality in a wild fish population. Ecology 2011, 92, 2175–2182. [Google Scholar] [CrossRef]
- Shelton, A.O.; Mangel, M. Fluctuations of fish populations and the magnifying effects of fishing. Proc. Natl. Acad. Sci. USA 2011, 108, 7075–7080. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Mehner, T.; Winfield, I.J.; Kangur, K.; Sarvala, J.; Gerdeaux, D.; Rask, M.; Malmquist, H.J.; Holmgren, K.; Volta, P.; et al. Impacts of climate warming on the long-term dynamics of key fish species in 24 European lakes. Hydrobiology 2012, 694, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Herschy, R.W.; Wohlfarth, B.; Kangur, K.; Kangur, A.; Raukas, A.; Pettersson, K.; Davis, J.; Carling, P.; Bengtsson, L. Peipsi Lake in Estonia/Russia. In Encyclopedia of Solid Earth Geophysics; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2012; pp. 596–607. [Google Scholar]
- Durant, J.M.; Hjermann, D.; Ottersen, G.; Stenseth, N.C. Climate and the match or mismatch between predator requirements and resource availability. Clim. Res. 2007, 33, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Søndergaard, M.; Lauridsen, T.L.; Beklioğlu, M.; Brucet, S.; Volta, P.; González-Bergonzoni, I.; et al. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73. [Google Scholar] [CrossRef] [Green Version]
- Woolway, R.I.; Dokulil, M.T.; Marszelewski, W.; Schmid, M.; Bouffard, D.; Merchant, C.J. Warming of Central European lakes and their response to the 1980s climate regime shift. Clim. Chang. 2017, 142, 505–520. [Google Scholar] [CrossRef]
- Olden, J.D.; Hogan, Z.S.; Zanden, M.J.V. Small fish, big fish, red fish, blue fish: Size-biased extinction risk of the world’s freshwater and marine fishes. Glob. Ecol. Biogeogr. 2007, 16, 694–701. [Google Scholar] [CrossRef]
- Helminen, H.; Sarvala, J. Population regulation of vendace (Coregonus albula) in Lake Pyhajarvi, southwest Finland. J. Fish Boil. 1994, 45, 387–400. [Google Scholar] [CrossRef]
- Marjomäki, T.J.; Auvinen, H.; Helminen, H.; Huusko, A.; Sarvala, J.; Valkeajärvi, P.; Viljanen, M.; Karjalainen, J. Spatial synchrony in the inter-annual population variation of vendace (Coregonus albula (L.)) in Finnish lakes. Ann. Zool. Fenn. 2004, 41, 225–240. [Google Scholar]
- Marjomäki, T.J.; Urpanen, O.; Karjalainen, J. Two-year cyclicity in recruitment of a fish population is driven by an inter-stage effect. Popul. Ecol. 2014, 56, 513–526. [Google Scholar] [CrossRef]
- Nyberg, P.; Bergstrand, E.; Degerman, E.; Enderlein, O. Recruitment of Pelagic Fish in an Unstable Climate: Studies in Sweden’s Four Largest Lakes. Ambio 2001, 30, 559–564. [Google Scholar] [CrossRef]
- Elliott, J.A.; Bell, V.A. Predicting the potential long-term influence of climate change on vendace (Coregonus albula) habitat in Bassenthwaite Lake, U.K. Freshw. Boil. 2010, 56, 395–405. [Google Scholar] [CrossRef]
- Winfield, I.J.; Adams, C.E.; Bean, C.W.; Durie, N.C.; Fletcher, J.M.; Gowans, A.R.; Harrod, C.; James, J.B.; Lyle, A.A.; Maitland, P.S.; et al. Conservation of the vendace (Coregonus albula), the U.K.’s rarest freshwater fish. Adv. Limnol. 2012, 63, 547–559. [Google Scholar]
- Valkeajärvi, P.; Auvinen, H.; Riikonen, R. Vendace stocks 2009–2010. Index of Density for Vendace, Perch, Smelt, Whitefish and Roach Almost for 100 Finnish lakesace Stocks 2009–2010. Available online: https://jukuri.luke.fi/bitstream/handle/10024/531246/rks2010_12.pdf?sequence=1 (accessed on 1 September 2020).
- Karjalainen, J.; Auvinen, H.; Helminen, H.; Arjomäki, T.J.; Niva, T.; Sarvala, J.; Vilhanen, M. Unpredictability of fish recruitment: Interannual variation in young-of-the-year abundance. J. Fish Boil. 2000, 56, 837–857. [Google Scholar] [CrossRef]
- Winfield, I.J.; Fletcher, J.M.; James, J.B. Conservation ecology of the vendace (Coregonus albula) in Bassenthwaite Lake and Derwent Water, U.K. Ann. Zool. Fenn. 2004, 41, 155–164. [Google Scholar]
- Auvinen, H. Factors affecting the year-class strength of vendace (Coregonus albula (L.)) in Lake Pyhäjärvi (Karelia, SE-Finland). Finn. Fish. Res. 1988, 9, 235–243. [Google Scholar]
- Tapaninen, M.; Keskinen, T.; Marjomäki, T.J. The seasonal final temperature preferenda of immature vendace, Coregonus albula (L.). Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 1998, 50, 131–141. [Google Scholar]
- Sarvala, J.; Helminen, H.; Auvinen, H. Portrait of a flourishing freshwater fishery: Pyhäjärvi, a lake in SW-Finland. Boreal Env. Res. 1999, 3, 329–345. [Google Scholar]
- Dembiński, W. Vertical distribution of vendace Coregonus albula L. and other pelagic fish species in some Polish lakes. J. Fish Boil. 1971, 3, 341–357. [Google Scholar] [CrossRef]
- George, D.G.; Bell, V.A.; Parker, J.; Moore, R.J. Using a 1-D mixing model to assess the potential impact of year-to-year changes in weather on the habitat of vendace (Coregonus albula) in Bassenthwaite Lake, Cumbria. Freshw. Boil. 2006, 51, 1407–1416. [Google Scholar] [CrossRef]
- Ohlberger, J.; Mehner, T.; Staaks, G.; Hölker, F. Temperature-related physiological adaptations promote ecological divergence in a sympatric species pair of temperate freshwater fish, Coregonusspp. Funct. Ecol. 2008, 22, 501–508. [Google Scholar] [CrossRef]
- Ohlberger, J.; Mehner, T.; Staaks, G.; Hölker, F. Is ecological segregation in a pair of sympatric coregonines supported by divergent feeding efficiencies? Can. J. Fish. Aquat. Sci. 2008, 65, 2105–2113. [Google Scholar] [CrossRef]
- Mehner, T.; Busch, S.; Helland, I.P.; Emmrich, M.; Freyhof, J. Temperature-related nocturnal vertical segregation of coexisting coregonids. Ecol. Freshw. Fish 2010, 19, 408–419. [Google Scholar] [CrossRef]
- Potapova, O.I. Large Vendace Coregonus Albula L.; Nauka: Leningrad, Russia, 1978. [Google Scholar]
- Tuvikene, L.; Saat, T. Vendace, Coregonus albula (L.). In Fishes of Estonia; Ojaveer, E., Pihu, E., Saat, T., Eds.; Estonian Academy Publishers: Tallinn, Estonia, 2001; pp. 114–120. [Google Scholar]
- Yefimova, A.I. Vendace in Lake Peipsi. In Hydrobiology and fisheries of Lake Peipsi-Pskov; Pihu, E., Mäting, A., Eds.; Valgus: Tallinn, Estonia, 1966; pp. 140–174. [Google Scholar]
- Viljanen, M. Biology, propagation, exploitation and management of vendace (Coregonus albula L.) in Finland. Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 1986, 22, 73–97. [Google Scholar]
- Auvinen, H. Intra- and interspecific factors in the dynamics of vendace (Coregonus albula (L.)) populations. Finn. Fish. Res. 1994, 15, 49–58. [Google Scholar]
- Sandström, A.; Ragnarsson-Stabo, H.; Axenrot, T.; Bergstrand, E. Has climate variability driven the trends and dynamics in recruitment of pelagic fish species in Swedish Lakes Vänern and Vättern in recent decades? Aquat. Ecosyst. Heal. Manag. 2014, 17, 349–356. [Google Scholar] [CrossRef]
- Ventelä, A.-M.; Kirkkala, T.; Lendasse, A.; Tarvainen, M.; Helminen, H.; Sarvala, J. Climate-related challenges in long-term management of Säkylän Pyhäjärvi (SW Finland). Hydrobiology 2010, 660, 49–58. [Google Scholar] [CrossRef]
- Helminen, H.; Sarvala, J.; Karjalainen, J. Patterns in vendace recruitment in Lake Pyhajarvi, south-west Finland. J. Fish Boil. 1997, 51, 303–316. [Google Scholar] [CrossRef]
- Heikinheimo, O. Effect of predation on the low-density dynamics of vendace: Significance of the functional response. Can. J. Fish. Aquat. Sci. 2001, 58, 1909–1923. [Google Scholar] [CrossRef]
- Valkeajärvi, P.; Marjomäki, T.J. Perch (Perca fluviatilis) as a factor in recruitment variations of vendace (Coregonus albula) in lake Konnevesi, Finland. Ann. Zool. Fenn. 2004, 41, 329–338. [Google Scholar]
- Karjalainen, J.; Ruokonen, T.J.; Marjomäki, T.J.; Martikainen, A.; Pursiainen, M.; Sarvala, J.; Tarvainen, M.; Ventelä, A.-M. Predation by signal crayfish Pacifastacus leniusculus on fish eggs and its consequences for coregonid recruitment. J. Fish Boil. 2015, 86, 651–667. [Google Scholar] [CrossRef]
- Clarke, S.J. Adapting to Climate Change: Implications for Freshwater Biodiversity and Management in the UK. Freshw. Rev. 2009, 2, 51–64. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. B: Boil. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [Green Version]
- Karjalainen, J.; Keskinen, T.; Pulkkanen, M.; Marjomäki, T.J. Climate change alters the egg development dynamics in cold-water adapted coregonids. Environ. Boil. Fishes 2014, 98, 979–991. [Google Scholar] [CrossRef]
- Stockwell, J.D.; Doubek, J.P.; Adrian, R.; Anneville, O.; Carey, C.C.; Carvalho, L.; Domis, L.N.D.S.; Dur, G.; Frassl, M.A.; Grossart, H.; et al. Storm impacts on phytoplankton community dynamics in lakes. Glob. Chang. Boil. 2020, 26, 2756–2784. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.; Calder, W.J.; Cumming, G.S.; Hughes, T.P.; Jentsch, A.; LaDeau, S.L.; Lenton, T.M.; Shuman, B.N.; Turetsky, M.R.; Ratajczak, Z.; et al. Climate change, ecosystems and abrupt change: Science priorities. Philos. Trans. R. Soc. B Boil. Sci. 2020, 375, 20190105. [Google Scholar] [CrossRef] [Green Version]
- Jaagus, J.; Sepp, M.; Tamm, T.; Järvet, A.; Mõisja, K. Trends and regime shifts in climatic conditions and river runoff in Estonia during 1951–2015. Earth Syst. Dyn. 2017, 8, 963–976. [Google Scholar] [CrossRef] [Green Version]
- Kotta, J.; Herkül, K.; Jaagus, J.; Kaasik, A.; Raudsepp, U.; Alari, V.; Arula, T.; Haberman, J.; Järvet, A.; Kangur, K.; et al. Linking atmospheric, terrestrial and aquatic environments: Regime shifts in the Estonian climate over the past 50 years. PLoS ONE 2018, 13, e0209568. [Google Scholar] [CrossRef]
- Kangur, K.; Möls, T. Changes in spatial distribution of phosphorus and nitrogen in the large north-temperate lowland Lake Peipsi (Estonia/Russia). Hydrobiology 2008, 599, 31–39. [Google Scholar] [CrossRef]
- Fink, G.; Burke, S.; Simis, S.G.H.; Kangur, K.; Kutser, T.; Mulligan, M. Management Options to Improve Water Quality in Lake Peipsi: Insights from Large Scale Models and Remote Sensing. Water Resour. Manag. 2018, 34, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jaani, A. The location, size and general characterisation of Lake Peipsi and its catchment area. In Lake Peipsi: Meteorology, Hydrology, Hydrochemistry; Nõges, T., Ed.; Sulemees Publishers: Tartu, Estonia, 2001; pp. 10–17. [Google Scholar]
- Estonian Environment Agency 2019/2020 aasta talv on esimene talv alates vaatlusajaloo algusest 1921. aastal kui Peipsil püsivat jääkatet ei tekkinud 2020. Available online: www.keskkonnaagentuur.ee (accessed on 28 April 2020).
- Vollenweider, R.A.; Kerekes, J. Eutrophication of Waters. Monitoring, Assessment and Control; OECD: Paris, France, 1982. [Google Scholar]
- Kangur, A.; Kangur, P.; Pihu, E.; Vaino, V.; Tambets, M.; Krause, T.; Kangur, K. Kalad ja kalandus. In Peipsi; Haberman, J., Timm, T., Raukas, A., Eds.; Eesti Loodusfoto Publishers: Tallinn, Estonia, 2008; pp. 317–340. [Google Scholar]
- Tammiksaar, E.; Kangur, K. Fish and fishing in Lake Peipsi (Estonia/Russia) since 1851: Similarities and differences between historical and modern times. J. Great Lakes Res. 2020, 46, 862–869. [Google Scholar] [CrossRef]
- Kangur, K.; Kangur, P.; Ginter, K.; Orru, K.; Haldna, M.; Möls, T.; Kangur, A. Long-term effects of extreme weather events and eutrophication on the fish community of shallow Lake Peipsi (Estonia/Russia). J. Limnol. 2013, 72, e30. [Google Scholar] [CrossRef] [Green Version]
- Kangur, K.; Park, Y.-S.; Kangur, A.; Kangur, P.; Lek, S. Patterning long-term changes of fish community in large shallow Lake Peipsi. Ecol. Model. 2007, 203, 34–44. [Google Scholar] [CrossRef]
- OnlineDoc 9.2.; SAS Institute Inc: Cary, NC, USA, 2008.
- Möls, T. Linear Statistical Methods for Estonian Freshwater Waterbodies; Estonian Naturalists’ Society: Tartu, Estonia, 2005. [Google Scholar]
- Hou, X.-G.; Siveter, D.J.; Aldridge, R.J.; Cong, P.-Y.; Gabbott, S.E.; Ma, X.; Purnell, M.A.; Williams, M. Algae; Wiley: Hoboken, NJ, USA, 2017; pp. 39–47. [Google Scholar]
- Dahlke, F.T.; Wohlrab, S.; Butzin, M.; Pörtner, H.-O. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science 2020, 369, 65–70. [Google Scholar] [CrossRef]
- Laugaste, R.; Haberman, J. Seasonality of zoo- and phytoplankton in Lake Peipsi (Estonia) as a function of water temperature. Proc. Estonian Acad. Sci. Biol. Ecol. 2005, 54, 18–39. [Google Scholar]
- Helminen, H.; Sarvala, J.; Hirvonen, A. Growth and food consumption of vendace (Coregonus albula (L.)) in Lake Pyhäjärvi, SW Finland: A bioenergetics modeling analysis. Hydrobiology 1990, 200, 511–522. [Google Scholar] [CrossRef]
- Pihu, E. The Importance of Pike, Perch, Pikeperch and Burbot as Biological Control in Lake Peipsi-Pskov; Valgus: Tallinn, Estonia, 1966; Volume 4. [Google Scholar]
- Kangur, A.; Kangur, P. The condition, length and age distribution of pikeperch, Stizostedion lucioperca (L.) in Lake Peipsi. Hydrobiology 1996, 338, 179–183. [Google Scholar] [CrossRef]
- Kangur, A.; Kangur, P. Diet composition and size-related changes in the feeding of pikeperch, Stizosteion lucioperca (Percidae) and pike, Esox lucius (Esocidae) in the Lake Peipsi (Estonia). Ital. J. Zool. 1998, 65, 255–259. [Google Scholar] [CrossRef]
- Lehtonen, H. Potential effects of global warming on northern European freshwater fish and fisheries. Fish. Manag. Ecol. 1996, 3, 59–71. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication; FishBase Mirror: Stockholm, Sweden, 2019. [Google Scholar]
- McCullough, D.R.; Reynolds, J.D.; Mace, G.M.; Redford, K.H.; Robinson, J.G. Conservation of Exploited Species. J. Wildl. Manag. 2003, 67, 656. [Google Scholar] [CrossRef]
- Reynolds, J.D. Life histories and extinction risk. In Macroecology; Blackburn, T.M., Gaston, K.J., Eds.; Blackwell: Oxford, UK, 2003; pp. 195–217. [Google Scholar]
- Kangur, K.; Kangur, A.; Kangur, P.; Laugaste, R. Fish kill in Lake Peipsi in summer 2002 as a synergistic effect of cyanobacterial bloom, high temperature and low water level. Proc. Estonian Acad. Sci. Biol. Ecol. 2005, 54, 67–80. [Google Scholar]
- Liso, S.; Gjelland, K.Ø.; Reshetnikov, Y.S.; Amundsen, P.-A. A planktivorous specialist turns rapacious: Piscivory in invading vendace Coregonus albula. J. Fish Boil. 2010, 78, 332–337. [Google Scholar] [CrossRef]
- Liso, S.; Gjelland, K.Ø.; Amundsen, P. Resource partitioning between pelagic coregonids in a subarctic watercourse following a biological invasion. J. Ichthyol. 2013, 53, 101–110. [Google Scholar] [CrossRef]
- Kangur, M.; Kangur, K.; Laugaste, R.; Punning, J.-M.; Möls, T. Combining limnological and palaeolimnological approaches in assessing degradation of Lake Pskov. Hydrobiology 2007, 584, 121–132. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. ISBN 978-92-9169-143-2. [Google Scholar]
- Woolway, R.I.; Kraemer, B.M.; Lenters, J.D.; Merchant, C.J.; O’Reilly, C.M.; Sharma, S. Global lake responses to climate change. Nat. Rev. Earth Environ. 2020. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | 57°51′–59°01′ N 26°57′–28°10′ E |
|---|---|
| Drainage area (km2) | 47,800 |
| Lake area (km2) | 3555 |
| Volume (km3) | 25.07 |
| Mean depth (m) | 7.1 |
| Maximum depth (m) | 15.3 |
| Hydrological turnover time (year) | 2 |
| Total phosphorus (mg P m−3) | 54 (13–220) |
| Total nitrogen (mg N m−3) | 748 (250–2100) |
| Chlorophyll a (mg m−3) | 29 (4–117) |
| Secchi disc water transparency (m) | 1.4 (0.4–3.8) |
| Trophic status [54] | eutrophic |
| Fish Species | Commercial Catch (103 kg) | Trawl Samples Catch (kg) | ||
|---|---|---|---|---|
| 1986 | 1998–2012 | 1986 | 1998–2012 | |
| Pikeperch Sander lucioperca | 75 (0.8%) | 1542 (25.1%) | 5.4 (1.4%) | 61.1 (28.5%) |
| Smelt Osmerus eperlanus | 1 556 (15.9%) | 860 (14%) | 15.9 (4.1%) | 1.9 (0.9%) |
| Common bream Abramis brama | 610 (6.2%) | 1037 (16.9%) | 169.7 (43.7%) | 66.0 (30.9%) |
| Eurasian perch Perca fluviatilis | 1 267 (12.9%) | 1101 (17.9%) | 92.1 (23.7%) | 40.3 (18.8%) |
| Roach Rutilus rutilus | 732 (7.5%) | 757 (12.3%) | 40.5 (10.4%) | 17.9 (8.4%) |
| Ruffe Gymnocephalus cernuus | 2 842 (29%) | 473 (7.7%) | 28.2 (7.3%) | 13.2 (6.2%) |
| Northern pike Esox lucius | 426 (4.3%) | 232 (3.8%) | 6.3 (1.6%) | 8.8 (4%) |
| Burbot Lota lota | 113 (1.2%) | 48 (0.8%) | 1.0 (0.3%) | 1.8 (0.8%) |
| Vendace Coregonus albula | 2 084 (21.2%) | 16 (0.3%) | 26.9 (6.9%) | 0.018 (0.007%) |
| Peipsi whitefish Coregonus lavaretus | 106 (1.1%) | 15 (0.2%) | 1.1 (0.3%) | 0.04 (0.02%) |
| Other fish species | 4 (0.0%) | 55 (1%) | 0.9 (0.2%) | 2.8 (1.3%) |
| Total | 9 815 (100%) | 6136 (100%) | 387.9 (100%) | 213.7 (100%) |
| Temperature Parameter | 1930–1960 | 1961–1985 | 1986–2010 | 1988 | 1989 | 1990 | 1991 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | S | Mean | Min | Max | S | Mean | Min | Max | S | Value | Value | Value | Value | |
| Ice-off date * (Julian day) | 107 | 91 | 117 | \ | 100 | 88 | 107 | – | 101 | 87 | 121 | – | 104 | 88 | 80 | 102 |
| Maxtemp * (°C) | 22.7 | 17.8 | 28.4 | / | 25.3 | 22.3 | 28.0 | – | 24.6 | 20.6 | 28.8 | – | 28.8 | 26.7 | 25.3 | 26.4 |
| Firsthightempday (17 °C) *(Julian day) | 163 | 127 | 196 | \ | 145 | 128 | 167 | – | 156 | 127 | 194 | – | 145 | 144 | 155 | 164 |
| Highdays (≥17 °C) *(days) | 64 | 8 | 107 | / | 84 | 46 | 102 | – | 79 | 48 | 110 | – | 90 | 91 | 81 | 77 |
| Ice formation date (Julian day) | 331 | 311 | 361 | – | 331 | 311 | 362 | – | 333 | 312 | 364 | / | 311 | 326 | 325 | 339 |
| Mean LSWT of 100–130 Julian days * | 3.76 | 0.62 | 8.42 | / | 5.96 | 2.98 | 9.46 | – | 5.97 | 0.0 | 9.19 | – | 3.85 | 9.71 | 12.8 | 5.90 |
| Time Lag | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| N | 68 | 66 | 64 | 62 | 60 | 58 | 56 | 54 |
| Pikeperch | −0.67 | −0.70 | −0.75 | −0.75 | −0.74 | −0.74 | −0.75 | −0.70 |
| Bream | −0.46 | −0.48 | −0.49 | −0.52 | −0.53 | −0.48 | −0.37 * | −0.32 * |
| Burbot | 0.64 | 0.62 | 0.60 | 0.61 | 0.64 | 0.67 | 0.62 | 0.59 |
| Perch | 0.41 ‡ | 0.40 ‡ | 0.38 † | 0.33 * | 0.28 * | 0.21 * | 0.06 * | −0.02 * |
| Pike | 0.35 † | 0.38 † | 0.34 * | 0.25 * | 0.21 * | 0.15 * | 0.14 * | 0.11 * |
| Vendace | 1.00 | 0.89 | 0.74 | 0.59 | 0.46 ‡ | 0.36 * | 0.30 * | 0.25 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kangur, K.; Ginter, K.; Kangur, A.; Kangur, P.; Möls, T. How Did the Late 1980s Climate Regime Shift Affect Temperature-Sensitive Fish Population Dynamics: Case Study of Vendace (Coregonus albula) in a Large North-Temperate Lake. Water 2020, 12, 2694. https://doi.org/10.3390/w12102694
Kangur K, Ginter K, Kangur A, Kangur P, Möls T. How Did the Late 1980s Climate Regime Shift Affect Temperature-Sensitive Fish Population Dynamics: Case Study of Vendace (Coregonus albula) in a Large North-Temperate Lake. Water. 2020; 12(10):2694. https://doi.org/10.3390/w12102694
Chicago/Turabian StyleKangur, Külli, Kai Ginter, Andu Kangur, Peeter Kangur, and Tõnu Möls. 2020. "How Did the Late 1980s Climate Regime Shift Affect Temperature-Sensitive Fish Population Dynamics: Case Study of Vendace (Coregonus albula) in a Large North-Temperate Lake" Water 12, no. 10: 2694. https://doi.org/10.3390/w12102694