Relationships Among Animal Communities, Lentic Habitats, and Channel Characteristics for Ecological Sediment Management

Abstract
:1. Introduction
2. Materials and Methods
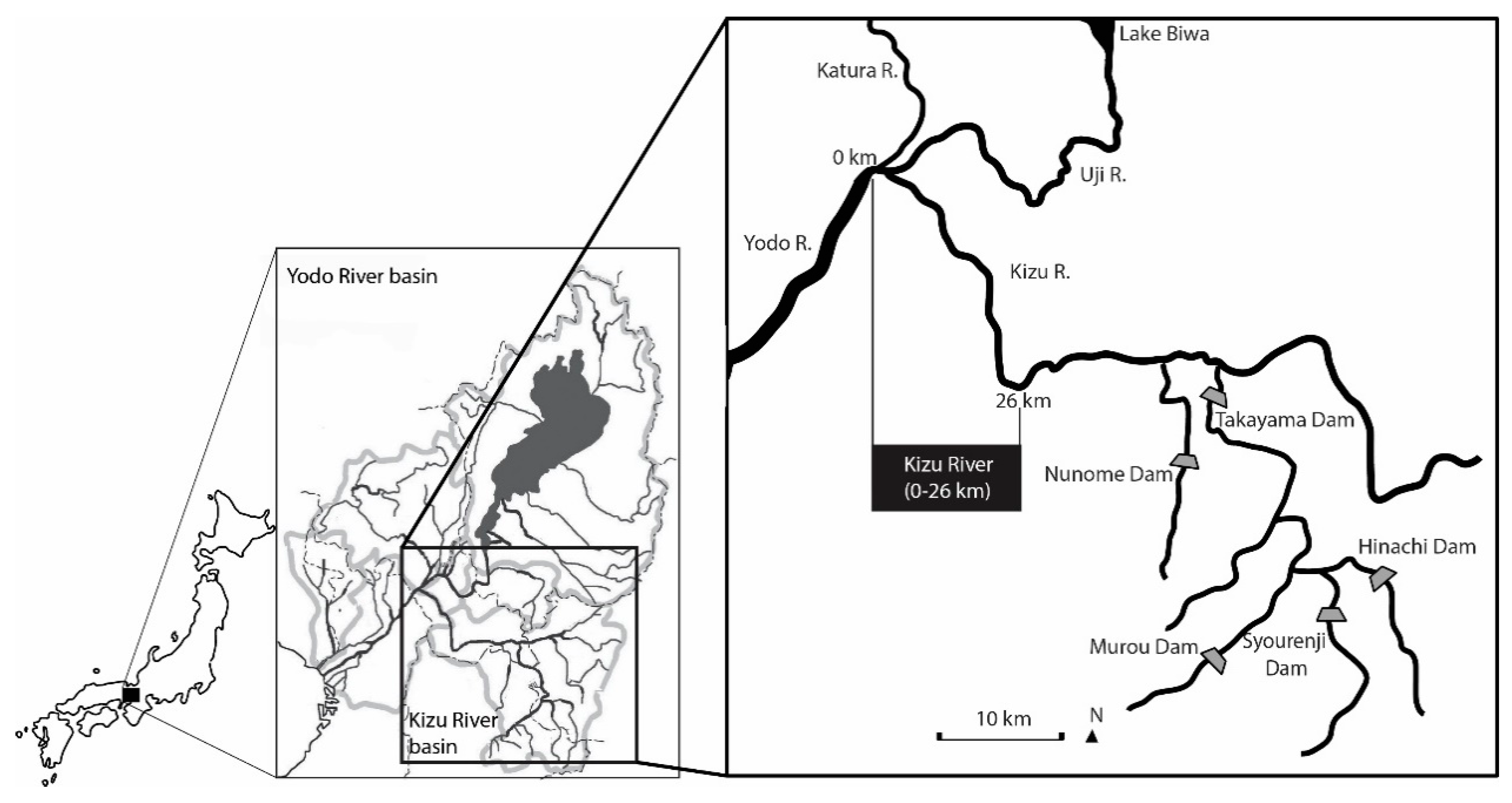
2.1. Study Site
2.2. Data Acquisition of Bitterling and Mussel Communities
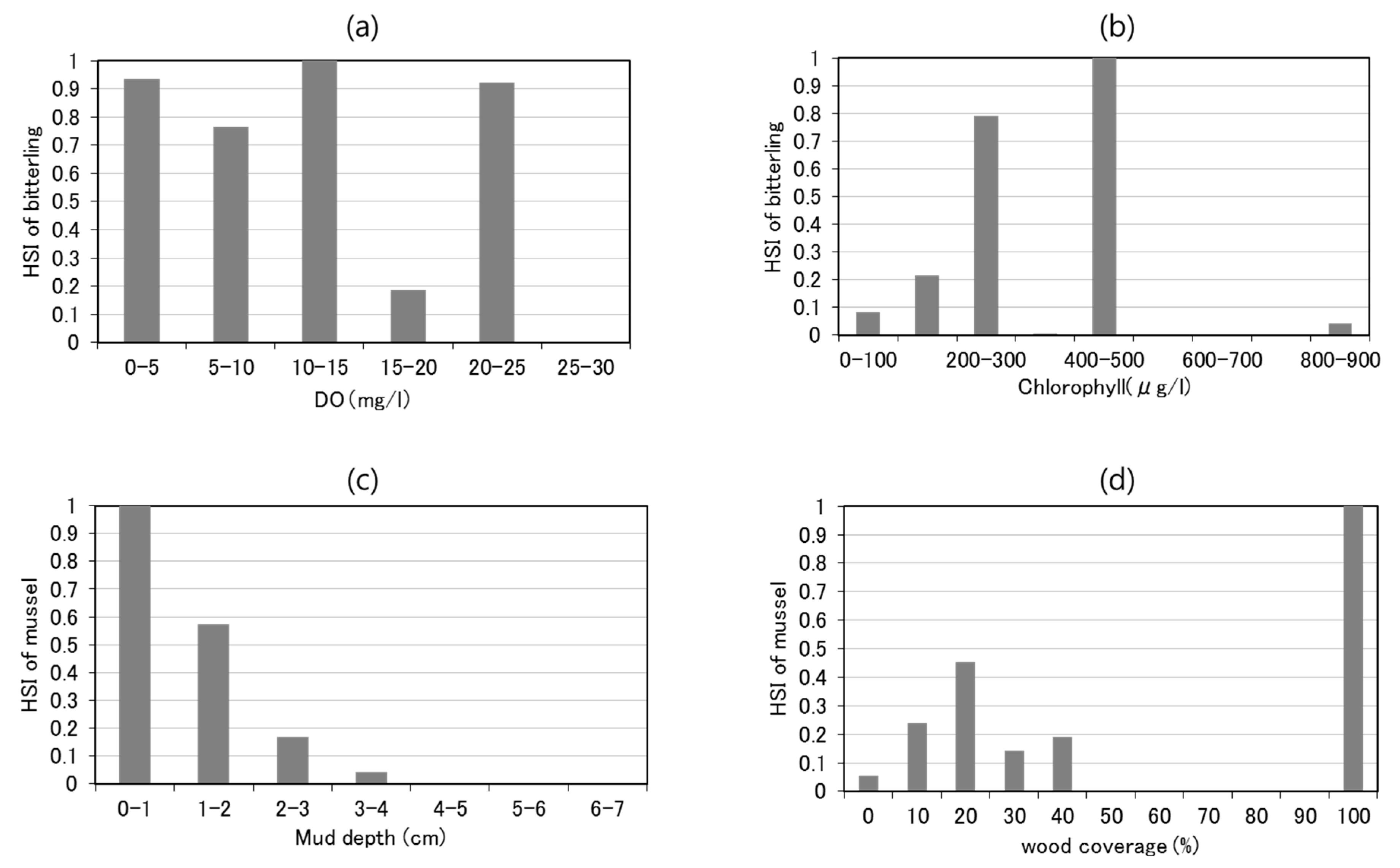
2.3. Data Acquisition of Lentic Habitat Structure and Conditions
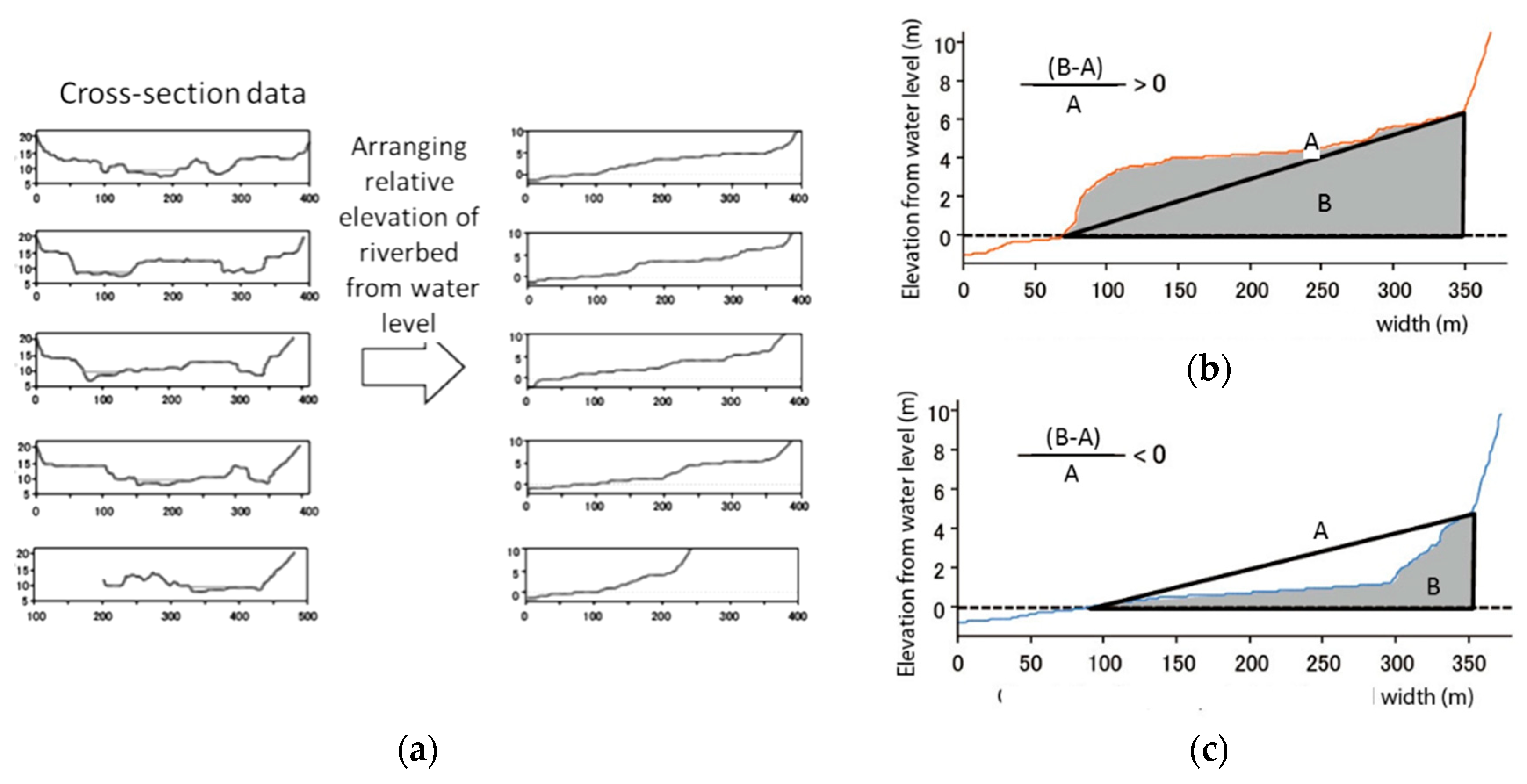
2.4. Reach-Scale Channel Characteristics
2.5. Statistical Analyses
- A multiple regression model using the parameters of habitat conditions for the abundance of bitterlings and mussels
- Comparison of the habitat suitability index (HSI) of bitterlings/mussels and habitat conditions
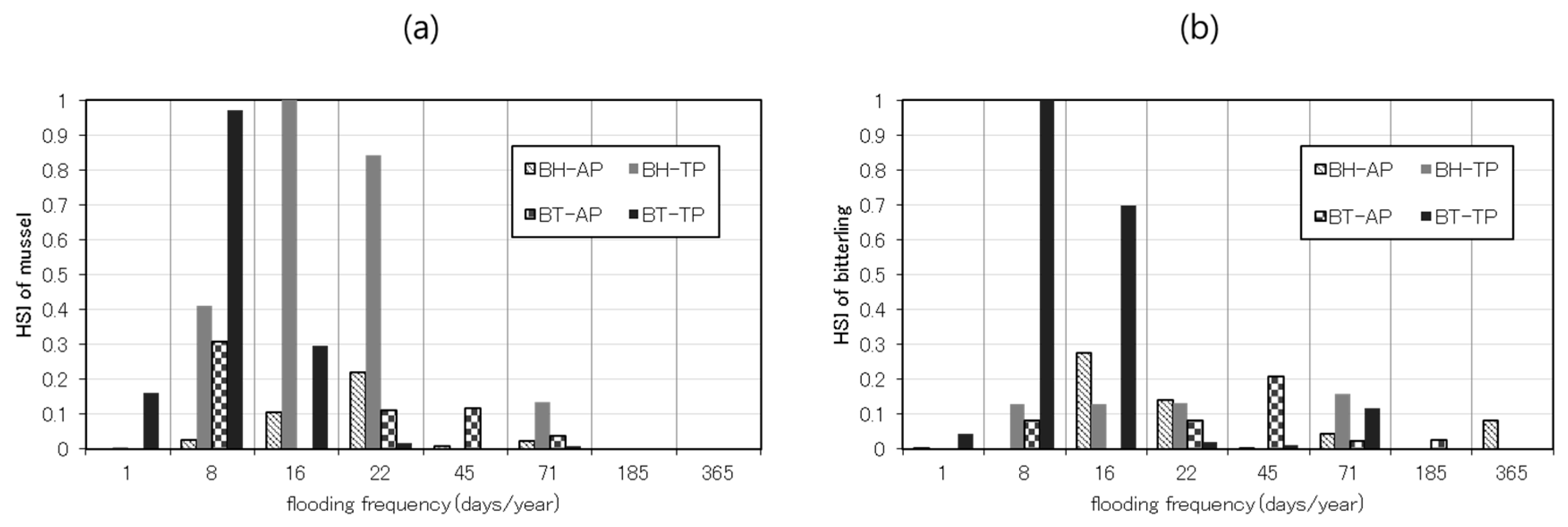
- Comparison of the HSI of bitterlings/mussels and habitat structures
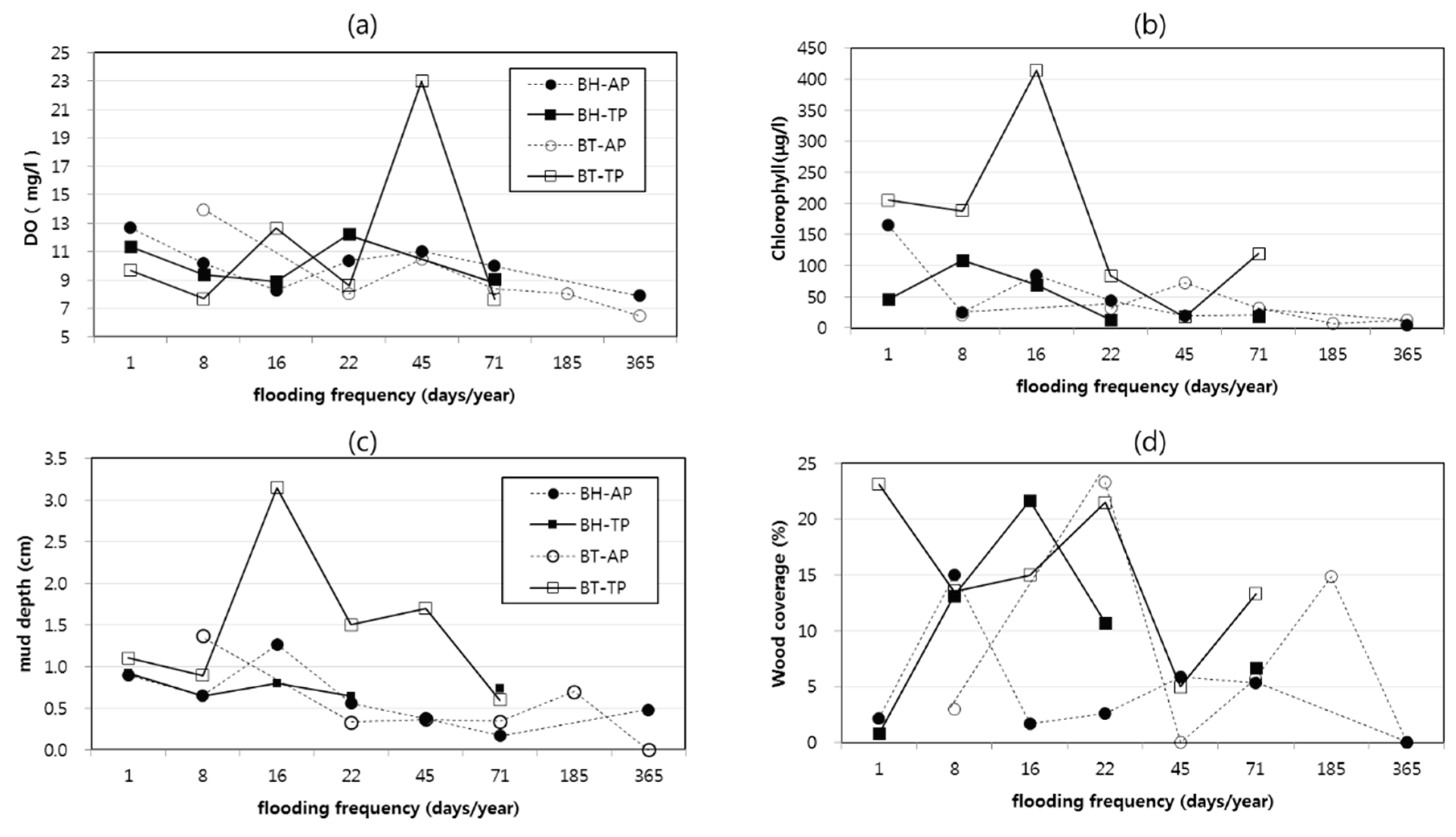
- Comparison of habitat structures and habitat conditions
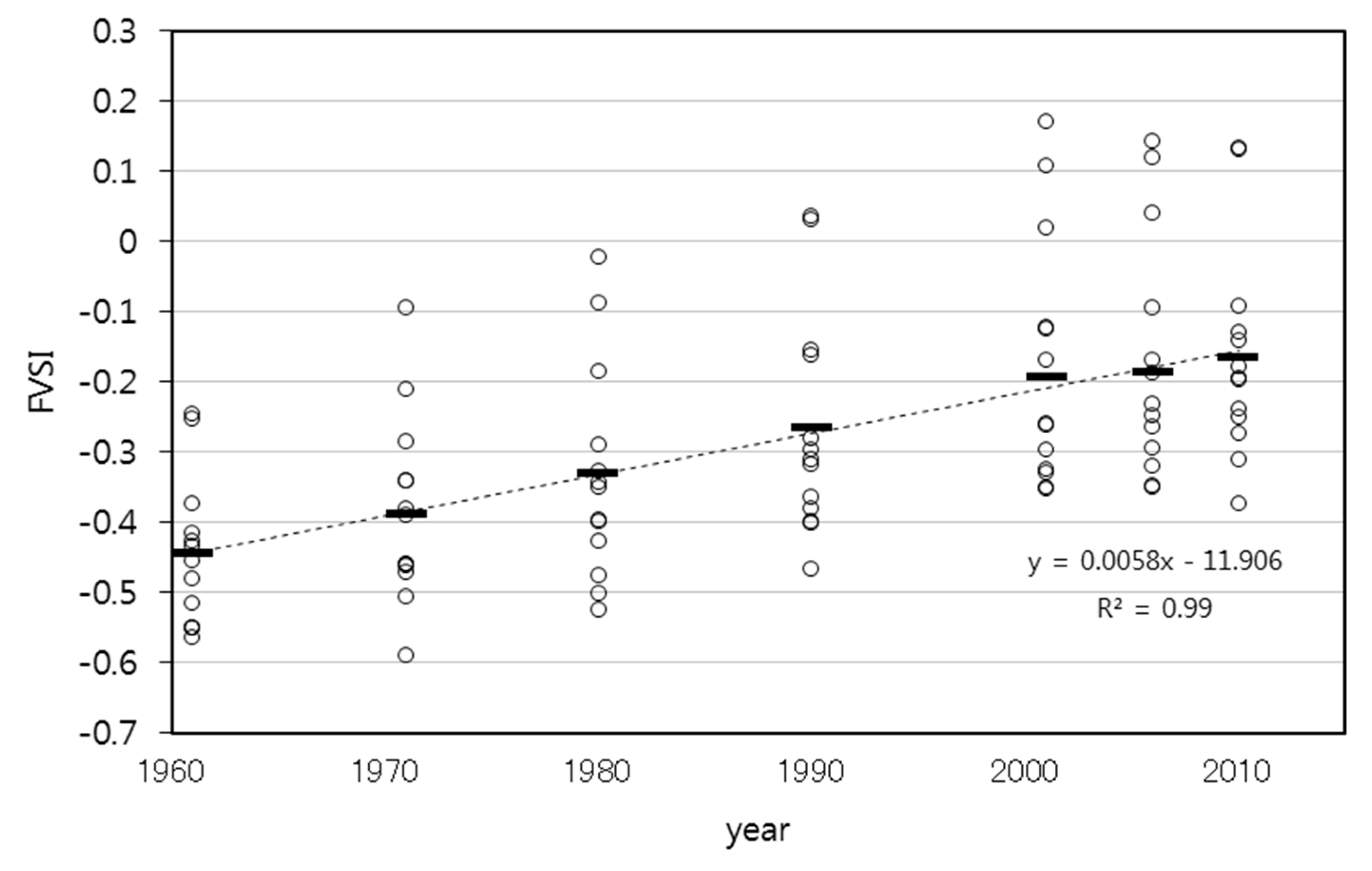
- Comparison of habitat structures and FVSI
3. Results
3.1. Relationships among the Bitterlings/Mussels, Habitat Structures, and Habitat Conditions
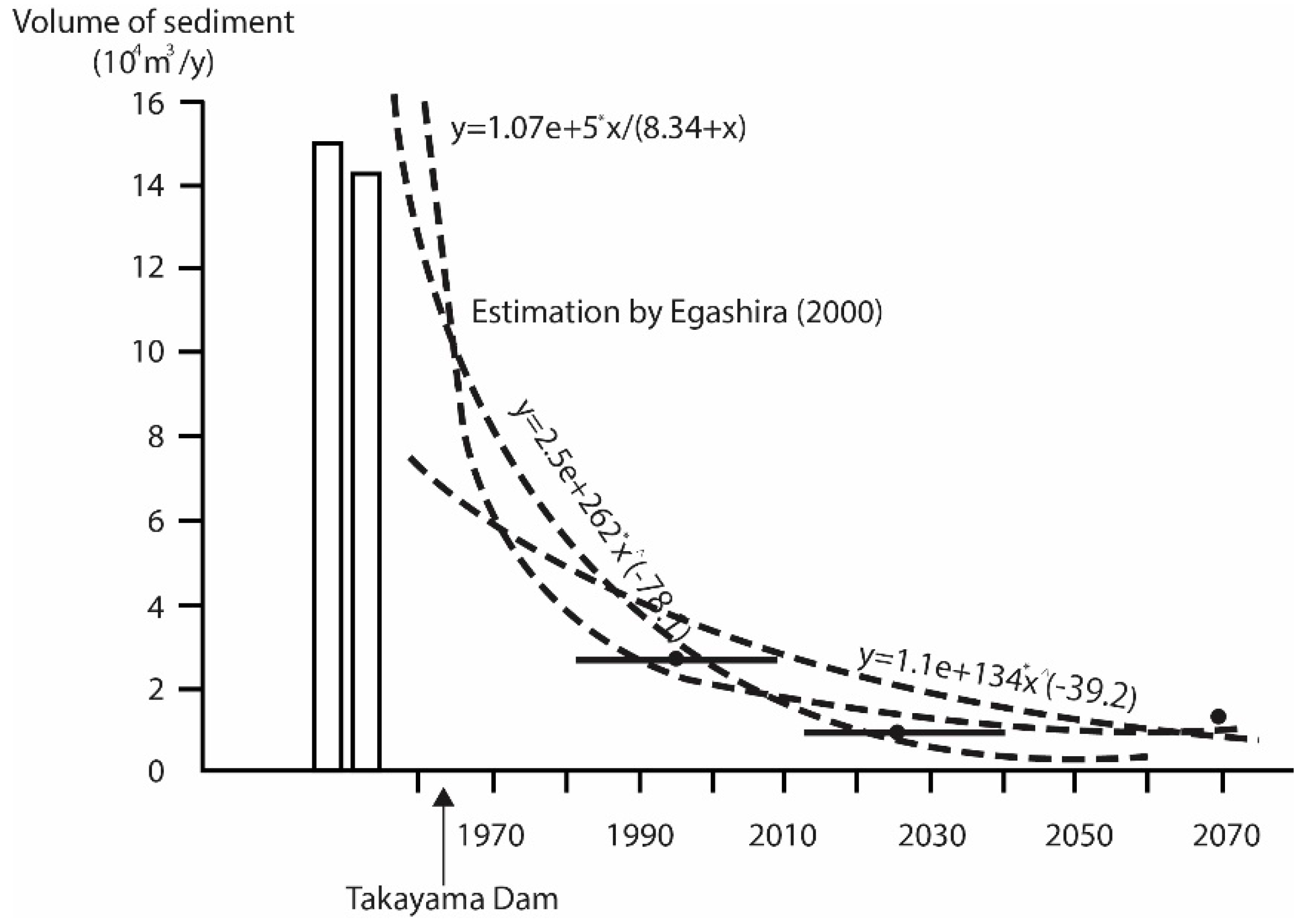
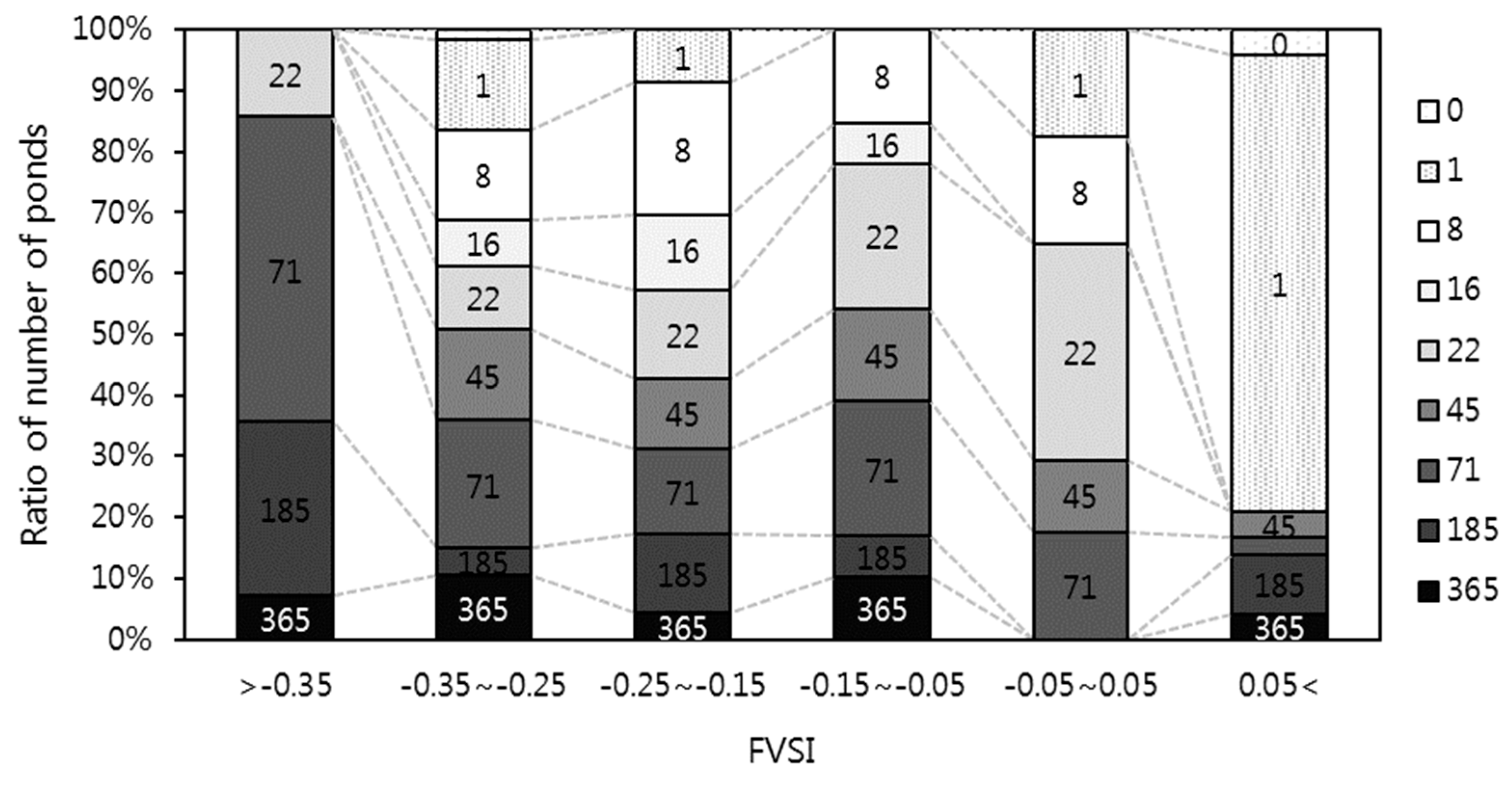
3.2. Historical Changes in FVSI
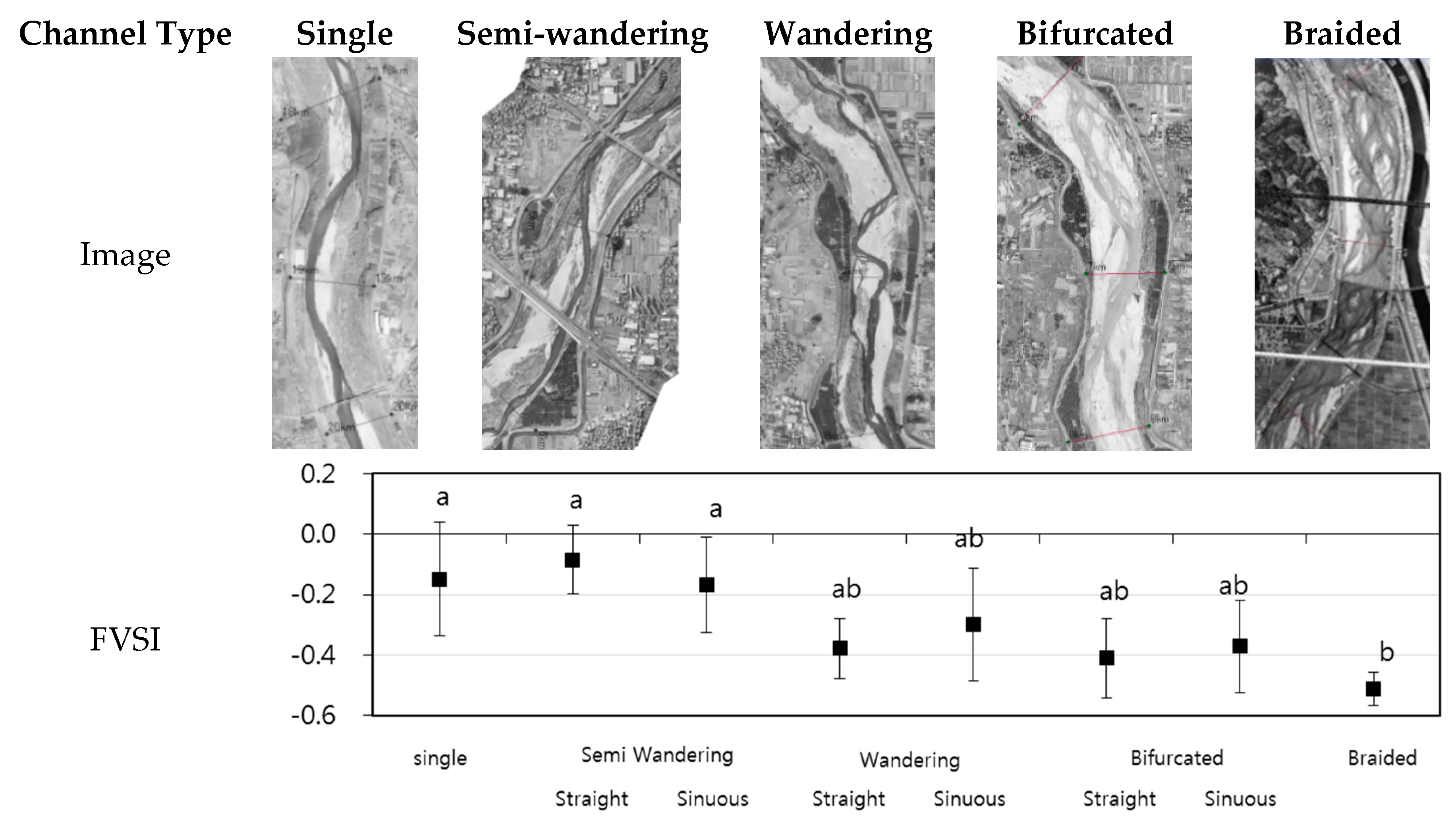
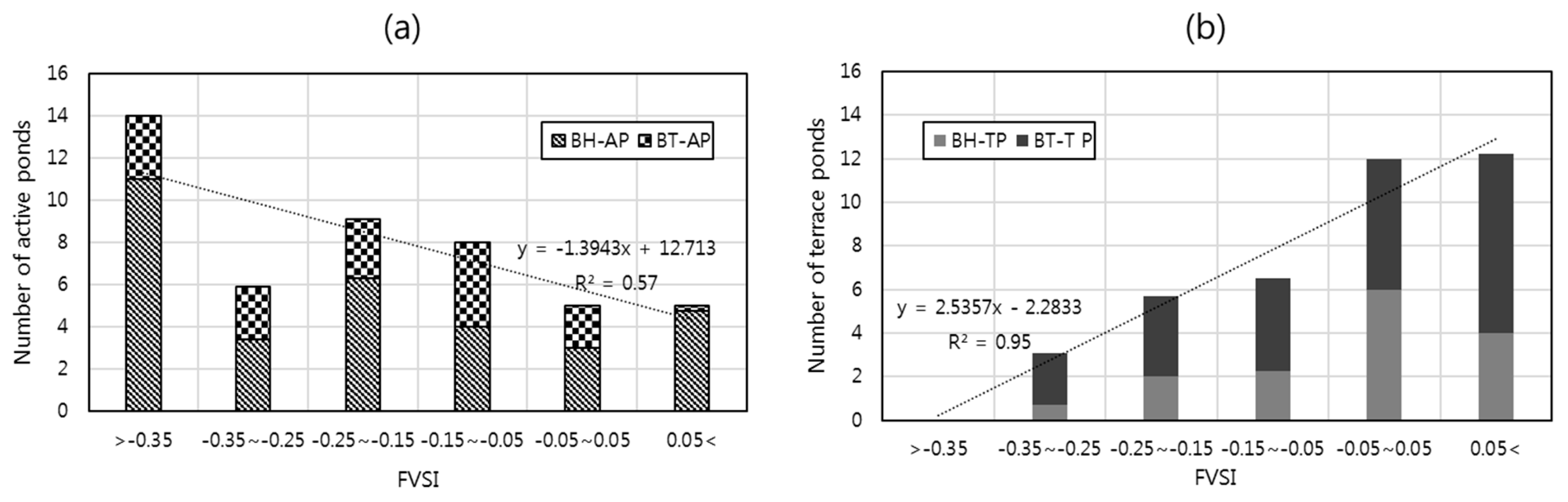
3.3. Relationship between the Habitat Structures and FVSI
4. Discussion
4.1. Relationships among Species Abundance and Habitat
4.2. Relationships between Habitats Structures and FVSI
4.3. Application of FVSI to Sediment Management
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kondolf, G.M.; Gao, Y.; Annandale, G.W.; Morris, G.L.; Jiang, E.; Zhang, J.; Cao, Y.; Carling, P.; Fu, K.; Guo, Q.; et al. Sustainable sediment management in reservoirs and regulated rivers: Experiences from five continents. Earth’s Future 2014, 2, 256–280. [Google Scholar] [CrossRef]
- Kondolf, G.M. Hungry Water: Effects of Dams and Gravel Mining on River Channels. Environ. Manag. 1997, 21, 533–551. [Google Scholar] [CrossRef]
- Auel, C.; Kobayashi, T.; Takemon, Y. Effects of sediment bypass tunnels on sediment grain size distribution and benthic habitats. Int. J. River Basin Manag. 2017, 15, 433–444. [Google Scholar] [CrossRef]
- Kantoush, S.A.; Sumi, T.; Kubota, A. Geomorphic response of rivers below dams by sediment replenishment technique. In River Flow 2010; Dittrich, A., Koll, K., Aberle, J., Geisenhainer, P., Eds.; Bundesanstalt für Wasserbau: Karlsruhe, Germany, 2010; pp. 1155–1163. [Google Scholar]
- Ock, G.; Sumi, T.; Takemon, Y. Sediment replenishment to downstream reaches below dams: Implementation perspectives. Hydrol. Res. Lett. 2013, 7, 54–59. [Google Scholar] [CrossRef]
- Wood, P.J.; Armitage, P.D. Biological effects of fine sediment in the lotic environment. Environ. Manag. 1997, 21, 203–217. [Google Scholar] [CrossRef]
- Leopold, L.B.; Wolman, M.G. River Channel Patterns: Braided, Meandering and Straight; US Geological Survey Professional Paper; U.S. Government Printing Office: Washington, DC, USA, 1957; Volume 282B, pp. 39–85.
- Schumm, S.A. Patterns of alluvial rivers. Annu. Rev. Earth Planet. Sci. 1985, 13, 5–27. [Google Scholar] [CrossRef]
- Rosgen, D.L. A classification of natural rivers. Catena 1994, 22, 169–199. [Google Scholar] [CrossRef] [Green Version]
- Brierley, G.; Fryirs, K. Geomorphology and River Management: Applications of the River Styles Framework; Blackwell: Oxford, UK, 2005. [Google Scholar]
- Wheaton, J.M.; Brasington, J.; Darby, S.E.; Merz, J.; Pasternack, G.B.; Sear, D.; Vericat, D. Linking geomorphic changes to salmonid habitat at a scale relevant to fish. River Res. Appl. 2010, 26, 469–486. [Google Scholar] [CrossRef]
- Wyrick, J.R.; Pasternack, G.B. Geospatial organization of fluvial landforms in a gravel-cobble river: Beyond the riffle-pool couplet. Geomorphology 2014, 213, 48–65. [Google Scholar] [CrossRef]
- Choi, M.; Takemon, Y.; Yu, W.; Jung, K. Ecological evaluation of reach scale channel configuration based on habitat structures for river management. J. Hydroinform. 2018, 20, 622–632. [Google Scholar] [CrossRef]
- Frothingham, K.M.; Rhoads, B.L.; Herricks, E.E. A multiscale conceptual framework for integrated eco-geomorphological research to support stream naturalization in the agricultural Midwest. Environ. Manag. 2002, 29, 16–23. [Google Scholar] [CrossRef]
- Sukhodolov, A.; Bertoldi, W.; Wolter, C.; Surian, N.; Tubino, M. Implication of channel processes for juvenile fish habitats in Alpine rivers. Aquat. Sci. 2009, 71, 338–349. [Google Scholar] [CrossRef]
- Payne, B.A.; Lapointe, M.F. Channel morphology and lateral stability: Effects on distribution of spawning and rearing habitat for Atlantic salmon in a wandering cobble-bed river. Can. J. Fish. Aquat. Sci. 1997, 54, 2627–2636. [Google Scholar] [CrossRef]
- Beechie, T.J.; Liermann, M.; Pollock, M.M.; Baker, S.; Davies, J. Channel pattern and river-floodplain dynamics in forested mountain river systems. Geomorphology 2006, 78, 124–141. [Google Scholar] [CrossRef]
- Tockner, K.; Ward, J.V.; Arscott, D.B.; Edwards, P.J.; Kollmann, J.; Gurnell, A.M.; Petts, G.E.; Maiolini, B. The Tagliamento River: A model ecosystem of European importance. Aquat. Sci. 2003, 65, 239–253. [Google Scholar] [CrossRef]
- Arscott, D.B.; Tockner, K.; van der Nat, D.; Ward, J.V. Aquatic Habitat Dynamics along a Braided Alpine River Ecosystem (Tagliamento River, Northeast Italy). Ecosystems 2002, 5, 802–814. [Google Scholar] [CrossRef]
- Kizu River Research Group. Integrated Research of the Kizu River II; Kizu River Research Group: Kyoto, Japan, 2003. (In Japanese) [Google Scholar]
- Negishi, J.N.; Sagawa, S.; Kayaba, Y.; Sanada, S.; Kume, M.; Miyashita, T. Mussel responses to flood pulse frequency: The important of local habitat. Freshw. Biol. 2012, 57, 1500–1511. [Google Scholar] [CrossRef]
- Takemon, Y.; Kobayashi, S.; Choi, M.; Terada, M.; Takebayashi, H.; Sumi, T. River Habitat Evaluation based on Cross-sectional Bed Profile and Frequency Distribution of Relative Elevation. Adv. River Eng. 2013, 19, 519–524. (In Japanese) [Google Scholar]
- Egashira, S.; Jin, H.; Takebayashi, H.; Nida, B.; Nagata, T. Bed variation and sediment budget in the downstream reach of Kizu River. Annu. J. Hydraul. Eng. 2002, 44, 777–782. (In Japanese) [Google Scholar] [CrossRef]
- Jowett, I.G.; Davey, A.J.H. A Comparison of Composite Habitat Suitability Indices and Generalized Additive Models of Invertebrate Abundance and Fish Presence-Habitat Availability. Trans. Am. Fish. Soc. 2007, 136, 428–444. [Google Scholar] [CrossRef]
- Bovee, K.D. Development and Evaluation of Habitat Suitability Criteria for Use in the Instream Flow Incremental Methodology; National Ecology Center, U.S. Fish and Wildlife Service: Washington, DC, USA, 1986. [Google Scholar]
- Survey Report on the Kizu River; Yodogawa River Bureau, ASIA AIR SURVEY CO., LTD.: Osaka, Japan, 2007. (In Japanese)
- Survey Report on the Kizu River; Yodogawa River Bureau, ASIA AIR SURVEY CO., LTD.: Osaka, Japan, 2009. (In Japanese)
- Survey Report on the Kizu River; Yodogawa River Bureau, ASIA AIR SURVEY CO., LTD.: Osaka, Japan, 2010. (In Japanese)
- Johnson, P.D.; Brown, K.M. The importance of microhabitat factors and habitat stability to the threatened Louisiana pearl shell, Mrgaritifera hembeli (Conrad). Can. J. Zool. 2000, 78, 271–277. [Google Scholar] [CrossRef]
- Yoshihiro, B.A.; Takashi, M. Habitat Characteristics Influencing Distribution of the Freshwater Mussel Pronodularia japanensis and Potential Impact on the Tokyo Bitterling, Tanakia tanago. Zool. Sci. 2010, 27, 912–916. [Google Scholar]
- Terada, M.; Takemon, Y.; Sumi, T. A study on habitat evaluation of tamari for bitterling and mussel. In Proceedings of the JSCE-Kansai, Uji, Japan, 26 August 2011; Volume II-12. (In Japanese). [Google Scholar]
- Global Re-introduction Perspectives: 2011; IUCN/SSC Re-Introduction Specialist Group: Calgary, AB, Canada, 2011.
- Reichard, M.; Liu, H.; Smith, C. The co-evolutionary relationship between bitterling fishes and freshwater mussels: Insights from interspecific comparison. Evolut. Ecol. Res. 2007, 9, 239–259. [Google Scholar]
- Ellis, M.M. Erosion silt as a factor in aquatic environments. Ecology 1936, 17, 29–42. [Google Scholar] [CrossRef]
- Kobayashi, S.; Takemon, Y. Historical Changes of Riffle Morphology for Benthic Invertebrate Habitats in the Kizu River. Annu. Disaster Prev. Res. Inst. Kyoto Univ. 2013, 56B, 681–689. (In Japanese) [Google Scholar]
- Surian, N.; Rinaldi, M. Morphological response to river engineering and management in alluvial channels in Italy. Geomorphology 2003, 50, 307–326. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abundance | Area (cm2) | Water Depth (cm) | Mud Depth (cm) | D50 (mm) | DO (mg/L) | Chlorophyll (μg/L) | Wood Coverage (%) | Best Model | |
|---|---|---|---|---|---|---|---|---|---|
| R | p | ||||||||
| Bitterling | −1.08 * | 2.52 * | 0.238 | <0.05 | |||||
| Mussel | −1.29 | 2.67 ** | 0.272 | <0.05 | |||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.; Takemon, Y.; Ikeda, K.; Jung, K. Relationships Among Animal Communities, Lentic Habitats, and Channel Characteristics for Ecological Sediment Management. Water 2018, 10, 1479. https://doi.org/10.3390/w10101479
Choi M, Takemon Y, Ikeda K, Jung K. Relationships Among Animal Communities, Lentic Habitats, and Channel Characteristics for Ecological Sediment Management. Water. 2018; 10(10):1479. https://doi.org/10.3390/w10101479
Chicago/Turabian StyleChoi, Mikyoung, Yasuhiro Takemon, Kinko Ikeda, and Kwansue Jung. 2018. "Relationships Among Animal Communities, Lentic Habitats, and Channel Characteristics for Ecological Sediment Management" Water 10, no. 10: 1479. https://doi.org/10.3390/w10101479