Crystal Structure of Bovine Alpha-Chymotrypsin in Space Group P65


Abstract
:1. Introduction
2. Materials and Methods
2.1. Macromolecule Production
2.2. Crystallization
2.3. Data Collection and Processing
2.4. Structure Solution and Refinement
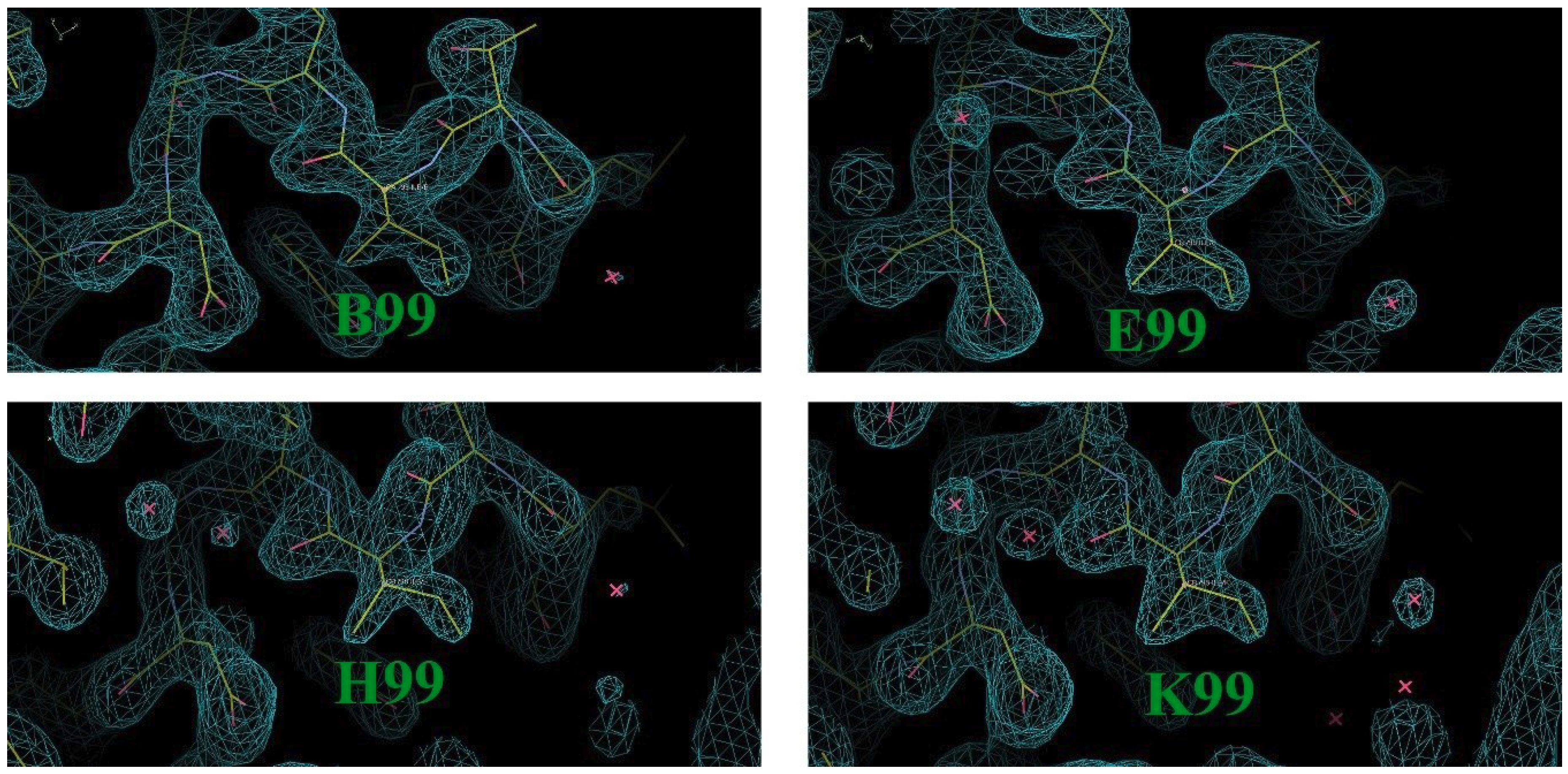
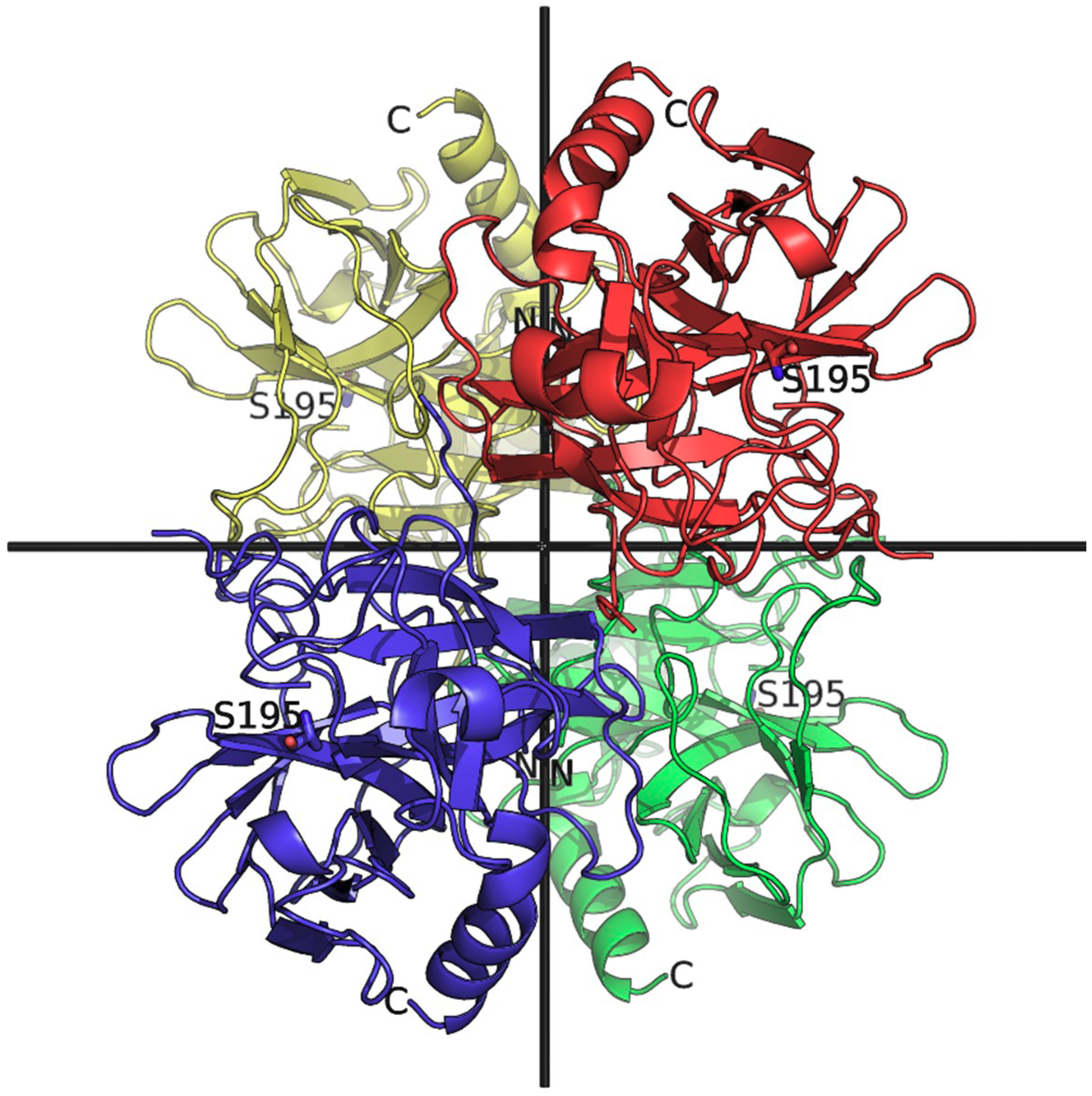
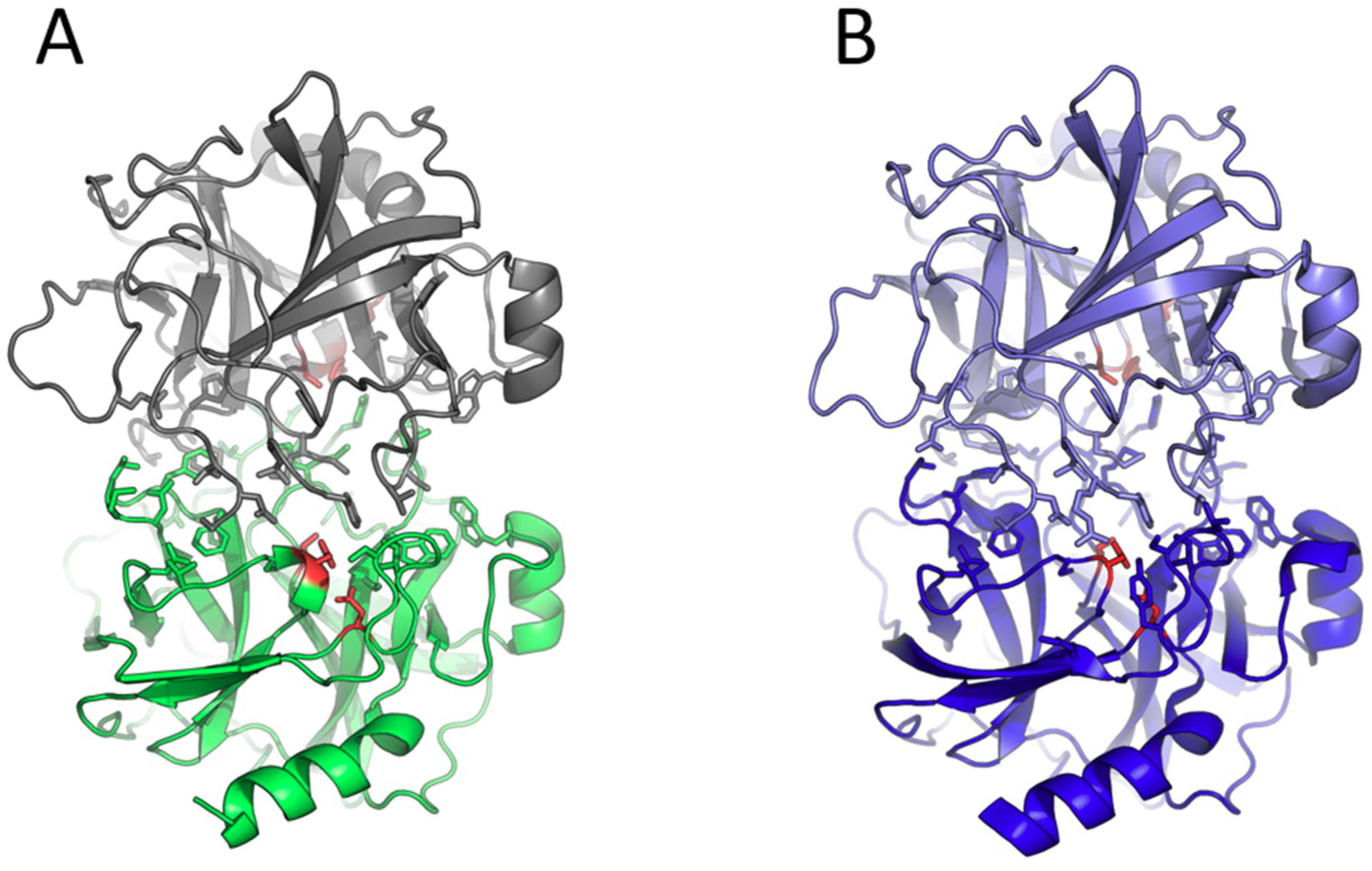
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, S.K.; Hopkins, T.R. Activation of bovine chymotrypsinogen-A - isolation and characterization of mu-chymotrypsin and omega-chymotrypsin. Biochemistry 1979, 18, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.W.; Sigler, P.B.; Henderson, R.; Blow, D.M. 3-dimensional structure of tosyl-alpha-chymotrypsin. Nature 1967, 214, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.C.; Pietsch, M.; Zhang, X.; Hautmann, S.; Chan, H.Y.; Bruning, J.B.; Gutschow, M.; Abell, A.D. Macrocyclic protease inhibitors with reduced peptide character. Angew. Chem. Int. Ed. Engl. 2014, 53, 7828–7831. [Google Scholar] [CrossRef] [PubMed]
- Roussel, A.; Mathieu, M.; Dobbs, A.; Luu, B.; Cambillau, C.; Kellenberger, C. Complexation of two proteic insect inhibitors to the active site of chymotrypsin suggests decoupled roles for binding and selectivity. J. Biol. Chem. 2001, 276, 38893–38898. [Google Scholar] [CrossRef] [PubMed]
- Neidhart, D.; Wei, Y.; Cassidy, C.; Lin, J.; Cleland, W.W.; Frey, P.A. Correlation of low-barrier hydrogen bonding and oxyanion binding in transition state analogue complexes of chymotrypsin. Biochemistry 2001, 40, 2439–2447. [Google Scholar] [CrossRef] [PubMed]
- McPhillips, T.M.; McPhillips, S.E.; Chiu, H.J.; Cohen, A.E.; Deacon, A.M.; Ellis, P.J.; Garman, E.; Gonzalez, A.; Sauter, N.K.; Phizackerley, R.P.; et al. Blu-Ice and the Distributed Control System: software for data acquisition and instrument control at macromolecular crystallography beamlines. J. Synchrotron Radiat. 2002, 9, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Leslie, A.G.W.; Powell, H.R. Processing Diffraction Data with Mosflm. In Evolving Methods for Macromolecular Crystallography; Read, R.J., Sussman, J.L., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 245, pp. 41–51. [Google Scholar]
- Karplus, P.A.; Diederichs, K. Assessing and maximizing data quality in macromolecular crystallography. Curr. Opin. Struct. Biol. 2015, 34, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Winn, M.D.; Isupov, M.N.; Murshudov, G.N. Use of TLS parameters to model anisotropic displacements in macromolecular refinement. Acta Crystallogr. D Biol. Crystallogr. 2001, 57, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrodinger, LLC. The PyMOL Molecular Graphics System, Version 1.8. 2015. [Google Scholar]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, H.; Blow, D.M. Structure of alpha-chymotrypsin refined at 1.68 A resolution. J. Mol. Biol. 1985, 184, 703–711. [Google Scholar] [CrossRef]
- Ikeda, K.; Kunugi, S.; Ise, N. pH dependence of the formation of dimeric alpha-chymotrypsin and its catalytic activity. J. Biochem. 1982, 92, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Corey, R.B.; Battfay, O.; Brueckner, D.A.; Mar, F.G. Preliminary X-ray diffraction studies of crystal forms of free and inhibited chymotrypsin. Biochim. Biophys. Acta 1965, 94, 535–545. [Google Scholar] [CrossRef]
- Vandlen, R.L.; Tulinsky, A. Changes in the tertiary structure of alpha-chymotrypsin with change in pH: p4 4.2–6.7. Biochemistry 1973, 12, 4193–4200. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Source Organism | Bos taurus |
|---|---|
| Complete amino acid sequence of chymotrypsinogen A (UniProt: P00766) | CGVPAIQPVL SGLSRIVNGE EAVPGSWPWQ VSLQDKTGFH FCGGSLINEN WVVTAAHCGV TTSDVVVAGE FDQGSSSEKI QKLKIAKVFK NSKYNSLTIN NDITLLKLST AASFSQTVSA VCLPSASDDF AAGTTCVTTG WGLTRYTNAN TPDRLQQASL PLLSNTNCKK YWGTKIKDAM ICAGASGVSS CMGDSGGPLV CKKNGAWTLV GIVSWGSSTC STSTPGVYAR VTALVNWVQQ TLAAN |
| Method | Vapour Diffusion, Hanging Drop |
|---|---|
| Plate type | 24-well tissue culture plate with glass cover slips sealed using vacuum grease |
| Temperature (K) | 289 |
| Protein concentration | 20 mg/mL |
| Buffer composition of protein solution | 0.1 M HEPES pH 7.0 |
| Composition of reservoir solution | 2 M (NH4)2SO4 |
| Volume and ratio of drop | 2 µL drop volume with 1:1 ratio of protein solution to reservoir solution |
| Volume of reservoir | 0.5 mL |
| Diffraction Source | Australian Synchrotron Beamline MX1 |
|---|---|
| Wavelength (Å) | 0.9537 |
| Temperature (K) | 100 |
| Detector | ADSC quantum 210r CCD |
| Crystal-detector distance (mm) | 184.95 |
| Rotation range per image (°) | 1 |
| Total rotation range (°) | 179 |
| Exposure time per image (s) | 1 |
| Space group | P65 |
| a, b, c (Å) | 126.8, 126.8, 122.0 |
| α, β, γ (°) | 90, 90, 120 |
| Mosaicity (°) | 0.53 |
| Resolution range (Å) | 29.55–1.86 (1.89–1.86) |
| Total No. of reflections | 868057 (31308) |
| No. of unique reflections | 93385 (4606) |
| Completeness (%) | 100.0 (99.7) |
| Redundancy | 9.3 (6.8) |
| 〈 I/σ(I)〉 | 12.2 (1.0 †) |
| CC1/2 | 0.998 (0.383) |
| Rmerge | 0.119 (1.924) |
| Rp.i.m. | 0.041 (0.779) |
| Overall B factor from Wilson plot (Å2) | 25.83 |
| Resolution range (Å) | 28.132–1.859 (1.925–1.859) |
| Completeness (%) | 99.81 (99.63) |
| σ cutoff | F > 0.0 σ(F) |
| No. of reflections, working set | 93185 (8787) |
| No. of reflections, test set | 4889 (468) |
| Final Rwork | 0.1885 (0.3317) |
| Final Rfree | 0.2252 (0.3416) |
| Cruickshank DPI (Å) | 0.219 |
| No. of non-H atoms | |
| Protein | 7201 |
| Sulphate ion | 65 |
| Solvent | 1100 |
| Total | 8366 |
| R.M.S. deviations | |
| Bonds (Å) | 0.0059 |
| Angles (°) | 0.78 |
| Average B factors (Å2) | |
| Protein | 33.1 |
| Sulphate ion | 80.2 |
| Water | 43.6 |
| Ramachandran plot | |
| Most favoured (%) | 97.45 |
| Allowed (%) | 2.13 |
| Interface # | Symmetry Operation | Interface Area (Å2) | Subunits † |
|---|---|---|---|
| 1 | X-Y, X+1, Z-1/6 | 1049 | B, C |
| X-Y, X, Z-1/6 | 1038 | A, D | |
| 2 | X, Y, Z | 668 | D, B |
| X, Y, Z | 625 | C, A | |
| 3 | X, Y, Z | 562 | D, A |
| X, Y, Z | 539 | C, B | |
| 4 | X, Y, Z | 344 | B, A |
| X, Y, Z | 340 | D, C | |
| 5 | -X-1, -Y, Z-1/2 | 361 | A, C |
| -X-1, -Y, Z-1/2 | 218 | B, D | |
| 6 | -X-1, -Y, Z-1/2 | 145 | A, D |
| -X-1, -Y, Z-1/2 | 136 | B, C |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marshall, A.C.; Keiller, B.G.; Pederick, J.L.; Abell, A.D.; Bruning, J.B. Crystal Structure of Bovine Alpha-Chymotrypsin in Space Group P65. Crystals 2018, 8, 460. https://doi.org/10.3390/cryst8120460
Marshall AC, Keiller BG, Pederick JL, Abell AD, Bruning JB. Crystal Structure of Bovine Alpha-Chymotrypsin in Space Group P65. Crystals. 2018; 8(12):460. https://doi.org/10.3390/cryst8120460
Chicago/Turabian StyleMarshall, Andrew C., Benjamin G. Keiller, Jordan L. Pederick, Andrew D. Abell, and John B. Bruning. 2018. "Crystal Structure of Bovine Alpha-Chymotrypsin in Space Group P65" Crystals 8, no. 12: 460. https://doi.org/10.3390/cryst8120460