
Pink-Beam Serial Synchrotron Crystallography at Pohang Light Source II

Abstract
:1. Introduction
2. Materials and Methods
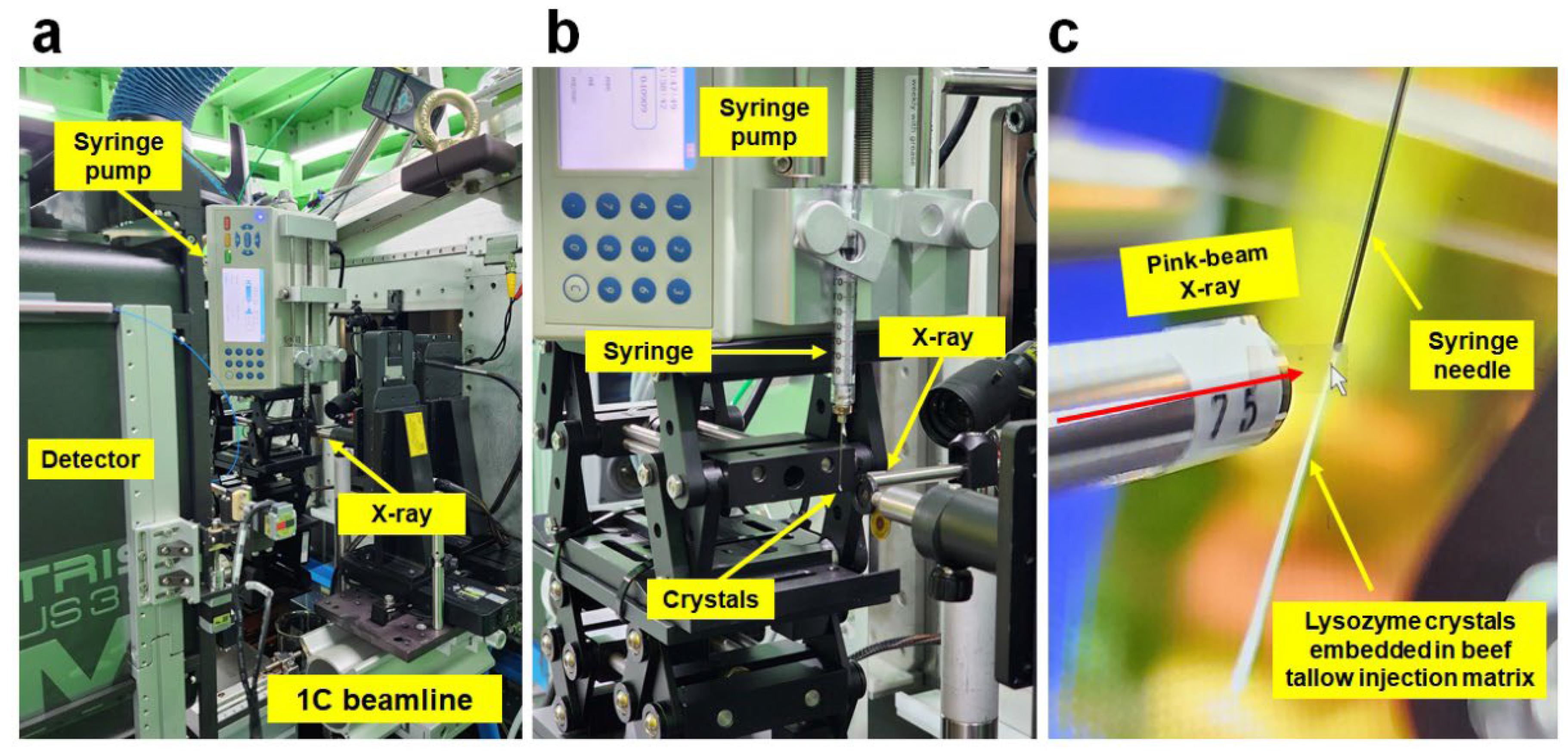
2.1. Sample Preparation
2.2. Data Collection
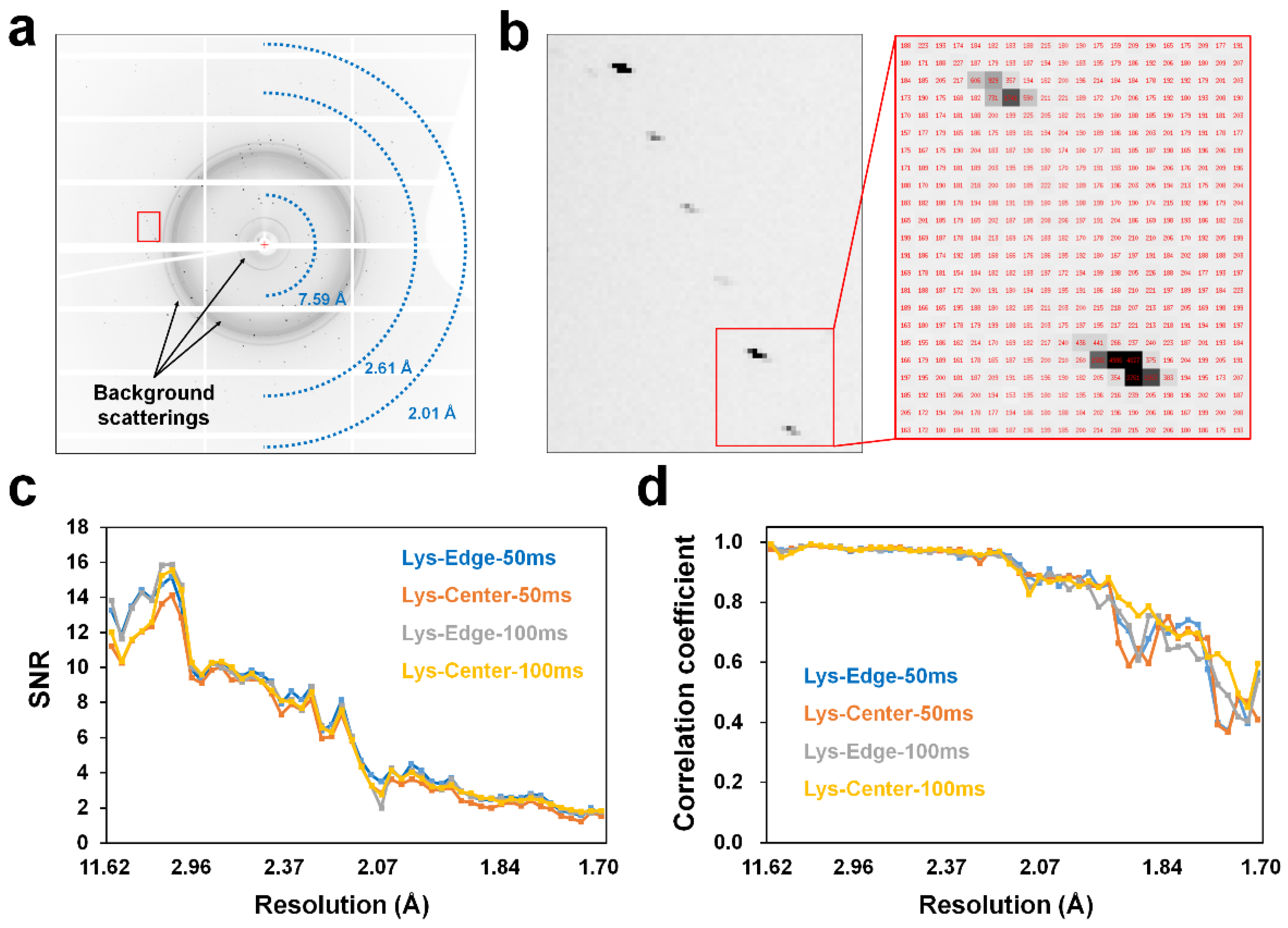
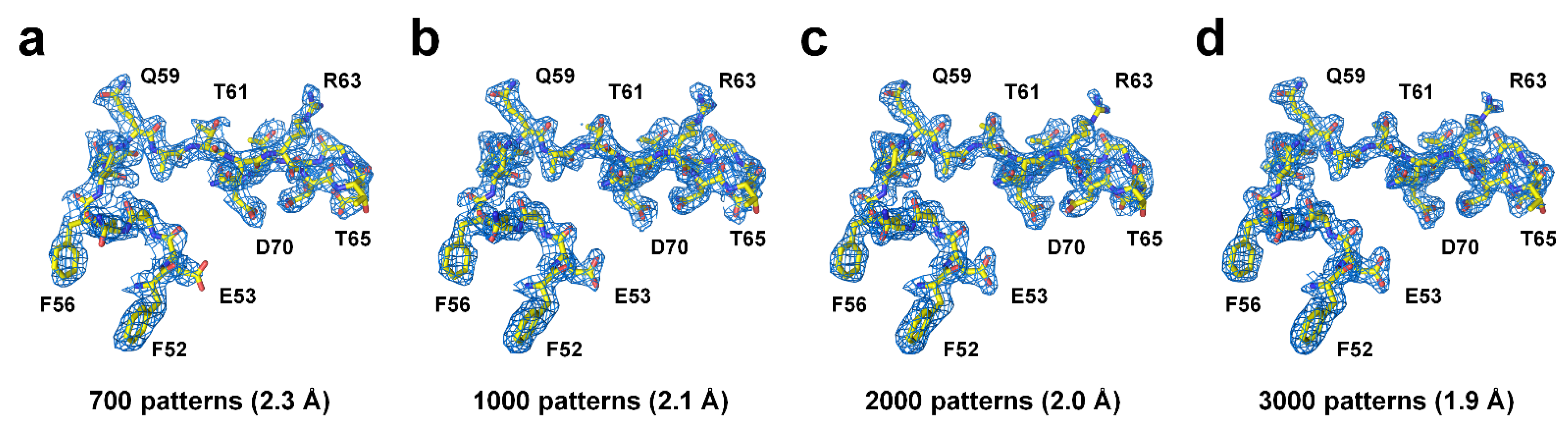
2.3. Data Processing and Analysis
2.4. Structure Determination
3. Results
3.1. Experimental Setup for Pink-Beam SSX
3.2. Pink-Beam SSX Data Collection
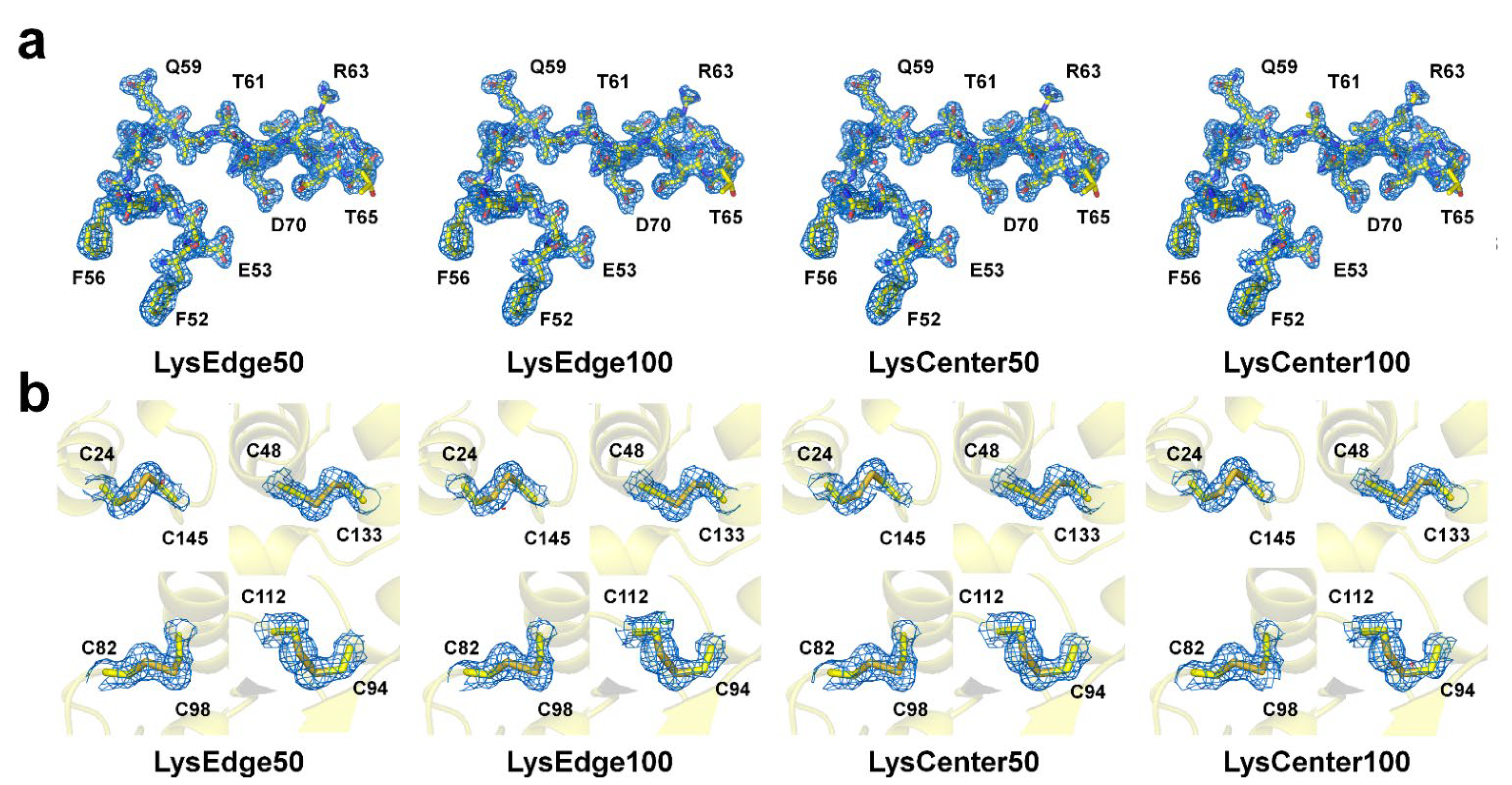
3.3. Structure Determination
3.4. Structure Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapman, H.N.; Caleman, C.; Timneanu, N. Diffraction before destruction. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, H.N.; Fromme, P.; Barty, A.; White, T.A.; Kirian, R.A.; Aquila, A.; Hunter, M.S.; Schulz, J.; DePonte, D.P.; Weierstall, U.; et al. Femtosecond X-ray protein nanocrystallography. Nature 2011, 470, 73–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutet, S.; Lomb, L.; Williams, G.J.; Barends, T.R.; Aquila, A.; Doak, R.B.; Weierstall, U.; DePonte, D.P.; Steinbrener, J.; Shoeman, R.L.; et al. High-resolution protein structure determination by serial femtosecond crystallography. Science 2012, 337, 362–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durdagi, S.; Dag, C.; Dogan, B.; Yigin, M.; Avsar, T.; Buyukdag, C.; Erol, I.; Ertem, F.B.; Calis, S.; Yildirim, G.; et al. Near-physiological-temperature serial crystallography reveals conformations of SARS-CoV-2 main protease active site for improved drug repurposing. Structure 2021, 29, 1382–1396. [Google Scholar] [CrossRef] [PubMed]
- Tenboer, J.; Basu, S.; Zatsepin, N.; Pande, K.; Milathianaki, D.; Frank, M.; Hunter, M.; Boutet, S.; Williams, G.J.; Koglin, J.E.; et al. Time-resolved serial crystallography captures high-resolution intermediates of photoactive yellow protein. Science 2014, 346, 1242–1246. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. Time-resolved macromolecular crystallography at pulsed X-ray sources. Int. J. Mol. Sci. 2019, 20, 1401. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Bean, R.; Sato, T.; Poudyal, I.; Bielecki, J.; Cruz Villarreal, J.; Yefanov, O.; Mariani, V.; White, T.A.; Kupitz, C.; et al. Time-resolved serial femtosecond crystallography at the European XFEL. Nat. Methods 2020, 17, 73–78. [Google Scholar] [CrossRef]
- Schmidt, M. Macromolecular movies, storybooks written by nature. Biophys. Rev. 2021, 13, 1191–1197. [Google Scholar] [CrossRef]
- Stellato, F.; Oberthur, D.; Liang, M.; Bean, R.; Gati, C.; Yefanov, O.; Barty, A.; Burkhardt, A.; Fischer, P.; Galli, L.; et al. Room-temperature macromolecular serial crystallography using synchrotron radiation. IUCrJ 2014, 1, 204–212. [Google Scholar] [CrossRef]
- Ko, I.S.; Kang, H.S.; Heo, H.; Kim, C.; Kim, G.; Min, C.K.; Yang, H.; Baek, S.Y.; Choi, H.J.; Mun, G.; et al. Construction and Commissioning of PAL-XFEL Facility. Appl. Sci. 2017, 7, 479. [Google Scholar] [CrossRef]
- Kang, H.S.; Min, C.K.; Heo, H.; Kim, C.; Yang, H.; Kim, G.; Nam, I.; Baek, S.Y.; Choi, H.J.; Mun, G.; et al. Hard X-ray free-electron laser with femtosecond-scale timing jitter. Nat. Photonics 2017, 11, 708–713. [Google Scholar] [CrossRef]
- Park, S.Y.; Choi, H.; Eo, C.; Cho, Y.; Nam, K.H. Fixed-target serial synchrotron crystallography using nylon mesh and enclosed film-based sample holder. Crystals 2020, 10, 803. [Google Scholar] [CrossRef]
- Nam, K.H.; Kim, J.; Cho, Y. Polyimide mesh-based sample holder with irregular crystal mounting holes for fixed-target serial crystallography. Sci. Rep. 2021, 11, 13115. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.H. Shortening injection matrix for serial crystallography. Sci. Rep. 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.H. Lard injection matrix for serial crystallography. Int. J. Mol. Sci. 2020, 21, 5977. [Google Scholar] [CrossRef]
- Meents, A.; Wiedorn, M.O.; Srajer, V.; Henning, R.; Sarrou, I.; Bergtholdt, J.; Barthelmess, M.; Reinke, P.Y.A.; Dierksmeyer, D.; Tolstikova, A.; et al. Pink-beam serial crystallography. Nat. Commun. 2017, 8, 1281. [Google Scholar] [CrossRef] [Green Version]
- Martin-Garcia, J.M.; Zhu, L.; Mendez, D.; Lee, M.-Y.; Chun, E.; Li, C.; Hu, H.; Subramanian, G.; Kissick, D.; Ogata, C.; et al. High-viscosity injector-based pink-beam serial crystallography of microcrystals at a synchrotron radiation source. IUCrJ 2019, 6, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H. Beef tallow injection matrix for serial crystallography. Sci. Rep. 2022, 12, 694. [Google Scholar] [CrossRef]
- Barty, A.; Kirian, R.A.; Maia, F.R.; Hantke, M.; Yoon, C.H.; White, T.A.; Chapman, H. Cheetah: Software for high-throughput reduction and analysis of serial femtosecond X-ray diffraction data. J. Appl. Crystallogr. 2014, 47, 1118–1131. [Google Scholar] [CrossRef] [Green Version]
- White, T.A.; Mariani, V.; Brehm, W.; Yefanov, O.; Barty, A.; Beyerlein, K.R.; Chervinskii, F.; Galli, L.; Gati, C.; Nakane, T.; et al. Recent developments in CrystFEL. J. Appl. Crystallogr. 2016, 49, 680–689. [Google Scholar] [CrossRef]
- Gevorkov, Y.; Yefanov, O.; Barty, A.; White, T.A.; Mariani, V.; Brehm, W.; Tolstikova, A.; Grigat, R.R.; Chapman, H.N. XGANDALF—Extended gradient descent algorithm for lattice finding. Acta Crystallogr. A Found. Adv. 2019, 75, 694–704. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkoczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, J.; Baek, S.; Park, J.; Park, S.; Lee, J.-L.; Chung, W.K.; Cho, Y.; Nam, K.H. Combination of an inject-and-transfer system for serial femtosecond crystallography. J. Appl. Crystallogr. 2022, 55, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Weik, M.; Ravelli, R.B.G.; Kryger, G.; McSweeney, S.; Raves, M.L.; Harel, M.; Gros, P.; Silman, I.; Kroon, J.; Sussman, J.L. Specific chemical and structural damage to proteins produced by synchrotron radiation. Proc. Natl. Acad. Sci. USA 2000, 97, 623–628. [Google Scholar] [CrossRef] [Green Version]
- De la Mora, E.; Coquelle, N.; Bury, C.S.; Rosenthal, M.; Holton, J.M.; Carmichael, I.; Garman, E.F.; Burghammer, M.; Colletier, J.-P.; Weik, M. Radiation damage and dose limits in serial synchrotron crystallography at cryo- and room temperatures. Proc. Natl. Acad. Sci. USA 2020, 117, 4142–4151. [Google Scholar] [CrossRef] [Green Version]
- Dejoie, C.; McCusker, L.B.; Baerlocher, C.; Abela, R.; Patterson, B.D.; Kunz, M.; Tamura, N. Using a non-monochromatic microbeam for serial snapshot crystallography. J. Appl. Crystallogr. 2013, 46, 791–794. [Google Scholar] [CrossRef]
- Weierstall, U.; James, D.; Wang, C.; White, T.A.; Wang, D.; Liu, W.; Spence, J.C.; Bruce Doak, R.; Nelson, G.; Fromme, P.; et al. Lipidic cubic phase injector facilitates membrane protein serial femtosecond crystallography. Nat. Commun. 2014, 5, 3309. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Park, S.; Kim, J.; Park, G.; Cho, Y.; Nam, K.H. Polyacrylamide injection matrix for serial femtosecond crystallography. Sci. Rep. 2019, 9, 2525. [Google Scholar] [CrossRef]
- Nam, K.H. Polysaccharide-Based Injection Matrix for Serial Crystallography. Int. J. Mol. Sci. 2020, 21, 3332. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.H.; Park, S.; Park, J. Preliminary XFEL data from spontaneously grown endo-1,4-β-xylanase crystals from Hypocrea virens. Acta Crystallogr. F Struct. Biol. Commun. 2022, 78, 226–231. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | LysEdge50 | LysCenter50 | LysEdge100 | LysCenter100 |
|---|---|---|---|---|
| X-ray source | 1C, PLS-II | 1C, PLS-II | 1C, PLS-II | 1C, PLS-II |
| X-ray energy (eV) | 15,860 | 15,860 | 15,860 | 15,860 |
| X-ray exposure (ms) | 50 | 50 | 100 | 100 |
| Hits images | 22,181 | 27,346 | 17,532 | 28,579 |
| Indexed images | 17,987 | 20,064 | 14,410 | 23,708 |
| Indexed patterns | 31,109 | 31,376 | 27,453 | 43,357 |
| Used patterns | 25,000 | 25,000 | 25,000 | 25,000 |
| Space group | P43212 | P43212 | P43212 | P43212 |
| Cell dimension (Å) a, b, c | 78.88, 78.88, 38.05 | 78.88, 78.88, 38.05 | 78.88, 78.88, 38.05 | 78.88, 78.88, 38.05 |
| Resolution (Å) | 80.6–1.70 (1.76–1.70) | 80.6–1.70 (1.76–1.70) | 80.6–1.70 (1.76–1.70) | 80.6–1.70 (1.76–1.70) |
| Unique reflections | 14,189 (1380) | 14,189 (1380) | 14,189 (1380) | 14,189 (1380) |
| Completeness (%) | 100.0 (100.0) | 100.0 (100.0) | 100.0 (100.0) | 100.0 (100.0) |
| Redundancy | 925.4 (503.9) | 551.7 (300.5) | 1256.7 (687.5) | 830.0 (455.6) |
| SNR | 6.78 (1.76) | 6.13 (1.50) | 6.73 (1.79) | 6.51 (1.88) |
| CC | 0.9880 (0.4404) | 0.9881 (0.4314) | 0.9891 (0.4787) | 0.9876 (0.5542) |
| CC* | 0.9969 (0.7820) | 0.9970 (0.7763) | 0.9972 (0.8046) | 0.9968 (0.8445) |
| Rsplit (%) a | 11.99 (71.60) | 12.43 (82.83) | 12.23 (64.22) | 12.35 (59.72) |
| Refinement | LysEdge50 | LysCenter50 | LysEdge100 | LysCenter100 |
|---|---|---|---|---|
| Resolution (Å) | 55.78–1.70 | 55.78–1.70 | 55.78–1.70 | 55.78–1.70 |
| Rwork a | 21.14 (28.18) | 21.77 (30.01) | 22.55 (27.90) | 22.73 (25.82) |
| Rfree b | 23.15 (33.21) | 23.41 (37.84) | 25.63 (34.56) | 25.64 (29.94) |
| R.m.s. deviations | ||||
| Bonds (Å) | 0.008 | 0.015 | 0.004 | 0.009 |
| Angles (°) | 1.100 | 1.473 | 0.758 | 2.383 |
| B factors (Å2) | ||||
| Protein | 22.17 | 21.02 | 20.40 | 19.45 |
| Ligand | 26.28 | 26.33 | 23.55 | 22.37 |
| Water | 35.43 | 32.06 | 32.27 | 32.23 |
| Ramachandran plot | ||||
| Favoured (%) | 99.21 | 99.43 | 99.21 | 99.21 |
| Allowed (%) | 0.79 | 0.57 | 0.79 | 0.79 |
| PDB | 8H8T | 8H8U | 8H8V | 8H8W |
| Data Processing | LysEdge50 | |||
|---|---|---|---|---|
| Diffraction Patterns | 700 | 1000 | 2000 | 3000 |
| Resolution (Å) | 80.6–2.30 (2.38–2.20) | 80.6–2.10 (2.17–2.10) | 80.6–2.00 (2.07–2.00) | 80.6–1.90 (1.97–1.90) |
| Unique reflections | 5903 (572) | 7674 (742) | 8847 (854) | 10264 (997) |
| Completeness (%) | 99.95 (99.65) | 99.97 (98.87) | 100.0 (100.0) | 100.0 (100.0) |
| Multiplicity | 42.2 (29.8) | 52.8 (37.1) | 88.0 (61.9) | 127.6 (83.4) |
| SNR | 1.92 (1.35) | 2.05 (1.09) | 2.58 (1.21) | 2.95 (1.32) |
| CC | 0.7082 (0.4656) | 0.7628 (0.4447) | 0.8499 (0.4072) | 0.8944 (0.5011) |
| CC* | 0.9106 (0.7971) | 0.9303 (0.7846) | 0.9585 (0.7607) | 0.9717 (0.8171) |
| Rsplit (%) | 53.61 (90.40) | 48.99 (117.90) | 37.08 (102.33) | 31.48 (92.34) |
| MR solution | ||||
| Top LLG | 1313.431 | 1857.711 | 2603.237 | 3253.941 |
| Top TFZ | 34.0 | 39.2 | 43.7 | 47.3 |
| Refinement | ||||
| Resolution (Å) | 55.78–2.30 | 55.78–2.10 | 55.78–2.00 | 55.78–1.90 |
| Rwork | 0.2671 | 0.2691 | 0.2552 | 0.2466 |
| Rfree | 0.3452 | 0.3607 | 0.2890 | 0.2960 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Nam, K.H. Pink-Beam Serial Synchrotron Crystallography at Pohang Light Source II. Crystals 2022, 12, 1637. https://doi.org/10.3390/cryst12111637
Kim Y, Nam KH. Pink-Beam Serial Synchrotron Crystallography at Pohang Light Source II. Crystals. 2022; 12(11):1637. https://doi.org/10.3390/cryst12111637
Chicago/Turabian StyleKim, Yongsam, and Ki Hyun Nam. 2022. "Pink-Beam Serial Synchrotron Crystallography at Pohang Light Source II" Crystals 12, no. 11: 1637. https://doi.org/10.3390/cryst12111637